
Abstract
Interleukin-2 (IL-2) is a cytokine that acts in dual and paradoxical ways in the immunotherapy of cancers and autoimmune diseases. Numerous clinical trial studies have shown that the use of different doses of this cytokine in various autoimmune diseases, transplantations, and cancers has resulted in therapeutic success. However, side effects of varying severity have been observed in patients. In recent years, to prevent these side effects, IL-2 has been engineered to bind more specifically to its receptors on the cell surface, decreasing IL-2 toxicities in patients. In this review article, we focus on some recent clinical trial studies and analyze them to determine the appropriate dose of IL-2 drug with the least toxicities. In addition, we discuss the engineering performed on IL-2, which shows that engineered IL-2 increases the specificity function of IL-2 and decreases its adverse effects.
Introduction
Cytokines are immune-modulating protein molecules that are secretory or membrane-bound, enabling immune cells to communicate and produce modified, strong, but self-limiting responses to antigens (Lee and Margolin, 2011). Researchers have investigated cytokines for their clinical activities in immunotherapy (Sim and Radvanyi, 2014). Interleukin-2 (IL-2) is a cytokine that plays an essential role in intercellular signaling, boosting the proliferation of T cells, and also maintaining immune homeostasis by initiating both immune-suppressing and activating responses (Kim et al., 2021; Papillion and Ballesteros-Tato, 2021).
IL-2 plays dual functions in immune responses. IL-2 is a powerful tool for immunologists to spread activated T cells and natural killer (NK) cells for stimulating immunity when present in purified and recombinant forms (Sim et al., 2014). However, by proliferating Treg cells, IL-2 can maintain homeostasis of innate and adaptive immune responses (Mitra and Leonard, 2018).
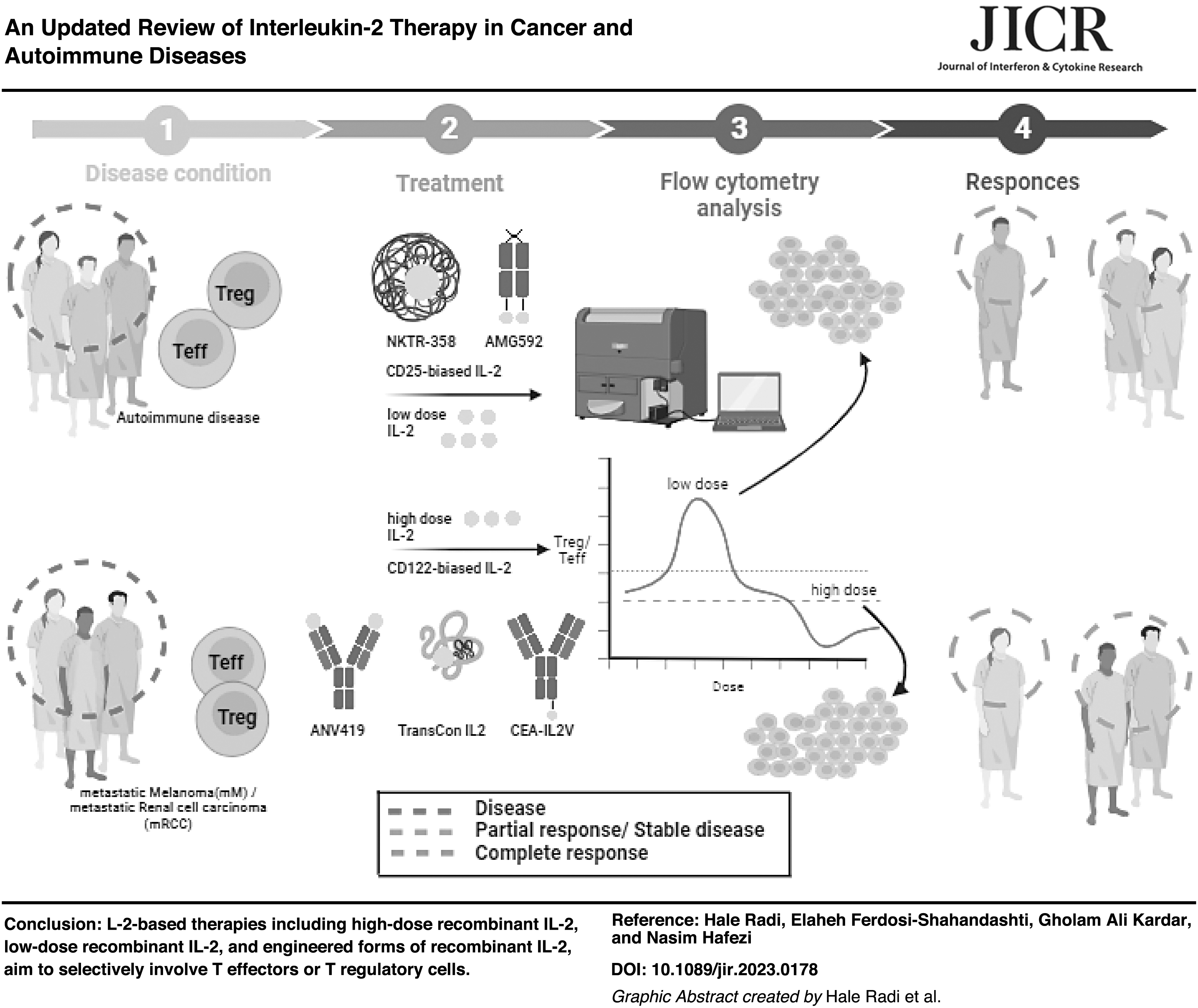
Given the significant therapeutic potential of IL-2 in the treatment of various diseases, this review article focuses on the molecular pathway of IL-2 and its effects on disease. We then examine recent studies on the optimal dose of IL-2 in autoimmune diseases and combined therapy in renal cell carcinoma (RCC) and metastatic melanoma (Mm). In addition, we review the latest IL-2 engineering developments.
Structure of IL-2, IL-2 Receptors, and Its Molecular Pathway
The accuracy of IL-2 receptor-binding sites and the structure of IL-2 is necessary for evaluating the foundation for the systematic design of their structure (Glassman et al., 2021). IL-2 is a member of a type I cytokines family that includes IL-4, IL-7, IL-9, IL-15, and IL-21, all of which consist of α-helical structures (De Paula et al., 2020; Hosseinian et al., 2018). IL-2 is a 15.5 kDa glycoprotein composed of 4 alpha helices (A-D) with a typical up-up-down-down 4-helix topology, which is produced by activated CD4+ and CD8+ T cells (Shao et al., 2019; Wang et al., 2021).
In addition, IL-2 made of 153 amino acids and post-translational modifications of O-linked glycosylation of threonine in position 3 and the intramolecular disulfide bridge between cysteines in positions 58 and 105 to link them together (Mitra et al., 2015).
The structure–activity relationship of IL-2 with IL-2 receptor (IL-2R) interaction may provide a clue to understanding how the cell senses the threshold of triggered IL-2s (Stauber et al., 2006). IL-2 has a multimeric receptor that is expressed on the cell surface. IL-2 signals preferably through a trimeric receptor complex consisting of the IL-2Rα chain (CD25), the IL-2Rβ chain (CD122), and the common gamma chain, IL-2Rγ (γc, CD132) (Zamecnik et al., 2020).
Variability in the expression of the IL-2Rα subunit (CD25) affects cell affinity for IL-2 (Beig Parikhani et al., 2022; Mitra et al., 2015). Respectively, the intermediate-affinity IL-2R heterodimer (β + γ) (KD ≈1 nM) and high-affinity IL-2R heterotrimer (β + α + γc) (KD ≈10 pM) comprise 3 separate noncovalent linked chains are expressed in CD4+ or CD8+ effector T (Teff) and CD4+ FoxP3+ regulatory T cells (Treg). Both dimeric and trimeric receptors are able to activate the cytoplasmic signaling cascade alongside IL-2 binding (Kim et al., 2021; Nirschl et al., 2022; Stauber et al., 2006; Zamecnik et al., 2020).
IL-2Rα is a nonsignaling chain that causes the rapid association rate of IL-2 binding and is expressed on the Treg cells' surface, whereas IL-2Rβ is responsible for its slow dissociation rate (Nirschl et al., 2022; Spangler et al., 2018; Stauber et al., 2006; Zamecnik et al., 2020), Therefore, Treg cells are more prone to IL-2 effects than T effector cells (Marcovecchio et al., 2020).
Binding IL-2 to IL-2Rβ and γc activates intracellular Janus kinase (JAK) 1/3. JAK proteins phosphorylate key tyrosine residues in the receptor intracellular domains, leading to the recruitment and activation of phosphoinositide-3-kinases (PI3Ks)/phosphatidylinositol 3,4,5-trisphosphate (PIP3)/AKT/mechanistic target of rapamycin/p70S6K, and Ras/Raf/mitogen-activated protein kinase kinases 1 and 2 (MAP2K1/2, also known as MEK1/2)/extracellular signal-regulated kinases 1 and 2 (ERK1/2) signaling cascades and the phosphorylation of STAT5 to trace gene expression of immune-related and regulated functional results (Pol et al., 2020; Spangler et al., 2018) (Fig. 1).
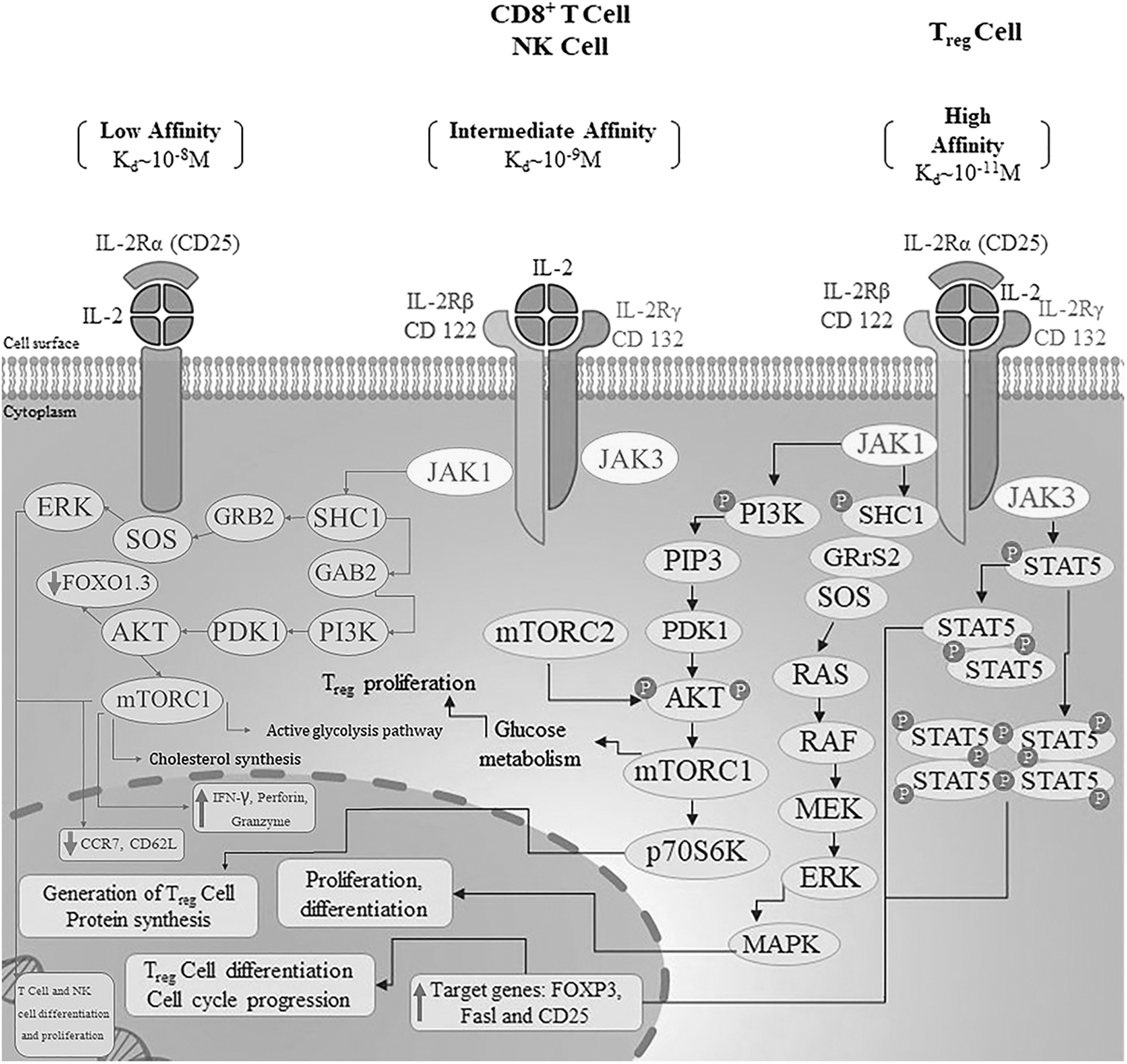
Schematic model illustrating types of IL-2Rs on CD8+ T/NK or Treg cells and the IL-2R molecular pathway in the cytoplasm and nuclear of T/NK cells. NK, natural killer.
Owing to the key role of STAT5 in the biogenesis and the maintenance of peripheral CD4+CD25+ Tregs, IL-2 cytokine has been characterized in autoimmune diseases (Spangler et al., 2018; Zorn et al., 2006). This introduces a new therapeutic approach for immunotherapy in patients with autoimmune and inflammatory diseases where using ultralow doses of IL-2 can selectively promote Treg cells in vivo, whereas Teff cells show favorable response to high concentrations of IL-2 (Marcovecchio et al., 2020; Zhang and Huang, 2021).
It should be mentioned that IL-2 synergistically boosts t cell receptor (TCR) signals (Ma et al., 2023). According to a recent study, the importance of IL-2 signaling depends on TCR signaling intensity. In the absence of efficient TCR signals, potent IL-2 signaling plays an inevitable role to determine CD4+ and CD8+ T cells' fate toward effector cells. Besides, it helps in generating durable functional memory T cells that contain valuable effects during recall responses (Chin et al., 2022).
Achievements About the IL-2-IL-2 Receptor Interaction, a Glance in the Past
Research on receptor-binding site of IL-2 to scrutinize the interaction of IL-2R α and β chains with IL-2 has been conducted during the 1980s–1990s. In the study by Arima et al. (1991) the D20K (replacement of aspartic acid with lysine) IL-2 variant, a β- subunit mutation, could attach to cells expressing IL-2Rα with low affinity. But, in the presence of α chain blocking antibody, the formation of the IL-2-IL-2R complex was completely inhibited. It could be inferred that α chain enhances the IL-2-IL-2R interaction even in the presence of mutants disrupting β chain binding.
This finding was confirmed in a subsequent research, which showed that IL-2 D20K caused the requirement of 10 times more IL-2 concentration to stimulate the IL-2R on T cells (Flemming et al., 1993). Based on the epitope mapping of neutralizing antibodies against α chain, it was found that mutations at positions 30–60, including R38A, F42A composed new IL-2 versions that were incapable of α chain binding, but not β (Sauve et al., 1991).
In addition, researchers tended to generate IL-2 mutants with increased binding affinity to α chain as high as IL-15-IL-15Rα to improve the antitumor or antiviral activity of T cells (Rao et al., 2005). The management of inflammatory and autoimmune diseases with the IL-2-IL-2R targeting approaches has garnered significant interest. As the Zheng et al. (1999) study reported, fusing IL-2 with fragment crystallization (FC) fragment of immunoglobulin G (IgG) was one of the early strategies to get a long-acting IL-2 response. This was accomplished through site-directed mutagenesis that blocked the C1q and FCγR binding sites (Zheng et al., 1999).
IL-2 as Immunostimulation Drug in Cancer Therapy
Since the 1970s, when the cytokine IL-2 was first discovered and cloned, it has been known for its potency in stimulating proliferation and activation of T effector and NK cells. Recombinant IL-2 (aldesleukin) has been approved by the US Food and Drug Administration (FDA) as the first cancer immunotherapeutic drug for the treatment of Mm and RCC (Graßhoff et al., 2021; Matsuoka, 2018; Shi et al., 2020).
The clinical dose for cancer immunotherapy is often as high as 60,000–720,000 IU/kg administrated once every 8 h, causing many side effects, including fever, joint pain, vascular leakage syndrome, pulmonary edema, hypotension, prerenal azotemia, oliguria, liver, and kidney function damage (Shah et al., 2021; Zhang and Huang, 2021). Because of these symptoms, standard high-dose (HD) IL-2 administration usually requires hospitalization and a dedicated medical team to monitor for capillary leak syndrome and other treatment-related toxicities (Bulgarelli et al., 2021).
In addition, only ∼5%–10% of patients get a result from high-dose IL-2 treatment (Zhang and Huang, 2021). Several trials are undergoing to determine the IL-2 optimal dose to obtain clinical efficacy without serious adverse effects in Mm and RCC (Table 1). A new point of view for getting maximum antitumor response to IL-2-based therapies is targeting tumor-specific T cells with high IL-2R in tumor microenvironment (TME).
Selected Clinical Studies with High-Dose Interleukin-2 Therapy at Different Dose Levels and Combination Treatment for Reducing Side Effects
MM, metastatic melanoma; mRCC, metastatic renal cell carcinoma; ccRCC, clear cell renal cell carcinoma; HD, high dose; LD, low dose; ED, escalating dose; IV, intravenous; PEM, pembrolizumab; IPI, ipilimumab; PR, partial response; SD, stable disease; CR, complete response; PD, progressive disease; SR, survival rate; AE, adverse event; AKI, acute kidney injury; LCD, lymphocyte count decreased; IRAEs, immune response adverse events; MTD, maximum tolerated dose; TIL, tumor infiltrating lymphocytes; RR, response rate; DCR, disease control rate; IMDC, international metastatic RCC database consortium.
According to a recent study, a mouse IL-2-CD25 fusion protein engineered to target high-affinity receptors, suppressed tumor growth by increased number and activation of tumor-specific CD8+ T cells without changing the frequency of Tregs in the TME, but not peripheral. The positive response to treatment occurs when the effector T cells are dominant over Tregs within the TME (LaPorte et al., 2023). Moreover, nano-based drug delivery systems to specifically release IL-2 at the tumor sites could be efficient to reduce stimulation of peripheral Tregs and risk of autoimmune presentations.
Furthermore, IL-2 therapy faces other challenges in stimulation of antitumor immunity. Tumor acid environment imposes an attenuated and unstable binding in the IL-2-IL-2R interaction. There have been efforts to design new versions of IL-2, such as Swich-2 and pH low insertion peptides (pHLIPs) with high performance and durability at acidic PH. Swich-2 showed strong binding affinity to IL-2Rα in an acidic environment as compared with neutral pH. By contrast, wild-type IL-2 showed an opposing function in the similar situation (Gaggero et al., 2022).
pHLIPs also opened new promising way to efficient function of antitumor agents. The pHLIPs are highly efficient in delivering antitumor agents into the cancer cell's membrane. This is due to their potential resistance at low pH environments, which undergoes a coli-α-helix transformation (Chu et al., 2023). Besides, to selective targeting of Teff cells and limiting therapy-related toxicities, other IL-2 variants with binding capability to intermediate-affinity have been developed and are undergoing clinical evaluation. A more detailed description of this part was mentioned in the “engineered IL-2-IL-2R” section.
IL-2 as Immunosuppression Drug
Today, IL-2 should rather be called an “immune regulatory” cytokine (Graßhoff et al., 2021). Thus, currently, establishing the doses of IL-2 and careful dose titration of it in autoimmune patients and transplant recipients is necessary to trigger Tregs but not Teffs (Marcovecchio et al., 2020; Spangler et al., 2018). Increasing the number of Treg cells to balance the immune system is essential to prevent the rejection of transplantation.
Calcineurin inhibitors (CNIs), including cyclosporine and tacrolimus, are immunosuppression drugs that are used in organ transplantation and suppressing activation and function of both effector T cells (Teffs) and Tregs. Previous reports showed that CNI therapy in both liver and kidney transplant recipients causes a low frequency of Tregs (van Kooten, 2017). In adults and children with autoimmune diseases, IL-2 at low doses, in the order of 1.0 × 106 IU per day, or ultra-low-dose (ULD), less than 1.0 × 106 IU per week, induces Tregs in a dose-related manner with no drug-dependent side effects but a small temporary non-itchy rash at the injection site (Marcovecchio et al., 2020).
In several recent clinical trials, IL-2 with different doses have been performed on autoimmune diseases such as systemic lupus erythematosus (SLE), type 1 diabetes (T1D), and rheumatoid arthritis (RA) to prove the appropriate therapeutic dose without any unusual safety signal. Several ongoing investigators-initiated trials are trying to find an efficient and safety dose for IL-2 (0.5–3 × 106 IU/m2/day) for graft-versus-host disease (GVHD) management and prophylaxis (Table 2).
Examples of Innovative Immunosuppressive Interleukin-2 Therapies Undergoing Clinical Evaluation
ALS, amyotrophic lateral sclerosis; SLE, systemic lupus erythematous; RA, rheumatoid arthritis; T1D, type 1 diabetes; GVHD, graft-versus-host-disease; RC GVHD, refractory chronic GVHD; ILI, influenza-like illness; MTX, methotrexate; SAE, severe adverse event; LCMV, lymphocytic choriomeningitis virus; Tfr, T follicular regulatory; HTreg, Treg high responder; DLT, dose-limiting toxicity; PPs, pediatric patients; DE, dose escalation; alloSCT, allogeneic stem cell transplantation; ULD IL-2, ultralow-dose IL-2; MRD, match related donor; ISI, injection-site induration; OR; overall response; IHIF, intrahepatic inflammatory response.
The investigators also observed a possible link between low-dose (LD) IL-2 augmented Tregs and inhibition of GVHD (Kennedy-Nasser et al., 2014). ULD IL-2 (1 × 105 U/m2) injection in pediatric patients with stem cell transplantation, especially from match-related donors, showed a significant elevation in the number of Tregs, along with reduced rate of GVHD and viral infection (Kennedy-Nasser et al., 2014).
According to a recent report, in addition to CD4+ CD25+ Tregs, LD IL-2 can enrich the IL-10 producing CD25+ regulatory B cells in both peripheral and splenic B cells (Inaba et al., 2023). However, the process behind this expansion remains unclear, given that B cells are generally not known for expressing functional IL-2 receptors due to the lack of CD122. The main challenges regarding the efficient and fast LD IL-2 response in controlled randomized trials, are optimizing the dosage schedule, limiting IL-2 off-target effects, and extending its serum half-life.
For example, administration of LD IL-2 in 4 treatment cycles with a dose of 1 million IU every other day for 2 weeks and 2 weeks interval did not show a notable difference in the improvement rate after 12 weeks of treatment. Whereas at week 24 an acceptable response rate was observed. In liver transplant recipients under treatment with tacrolimus, LD IL-2 could revive the number of Tregs but did not result in allogeneic tolerance.
Besides, in GVHD cases, it has been detected that both IL-2 and Tregs decreased over time after peaking, despite the continuation of IL-2 infusion. A dose-escalating trial on children and adult cGVHD established that for children LD IL-2 at the maximum tolerated dose (MTD) of 1 × 106 IU/m2/day reached to the highest level with a 7.5-fold increase at week 5, followed by 5-fold higher than baseline at week 8. In adults the maximum IL-2 injected was 2 × 106 IU/m2/day, lead to a rapid 10-fold increase from the baseline at week 1. Thereafter, a rapid decrease was observed as it was detected just a 4-fold increase than baseline at week 8 (Whangbo et al., 2019).
Current trials illustrated that IL-2 activity can be adjusted with engineering methods to engage effectively with high-affinity IL-2R that leads to amplify immunoregulation in autoimmune conditions and GVHD. We presented them in “engineered IL-2-IL-2R” section.
Engineered IL-2 Recombinant Proteins for Therapeutic Development
Nowadays, several approaches have been pursued to develop next-generation engineered IL-2 to increase half-life and decrease toxicity of it while retaining beneficial effects in antitumor immunity, or switching to immunomodulatory actions in autoimmune conditions. The modifications are designed to guide its binding to low-affinity dimeric IL-2 receptors on CD8+ T and NK cells or high-affinity trimeric receptors on Treg cells.
The 4 main strategies being used for IL-2 engineering are IL-2-anti IL-2 immune complexes, IL-2 muteins with affinity to IL-2Rα or IL-2Rβγ, site-specific pegylation, and IL-2 fusion proteins (Fig. 2). Developing new variants of IL-2 has opened up a new way to selectively involve immune responses and minimize the activation of unwanted cells. Herein, new achievements in the field have been mentioned.
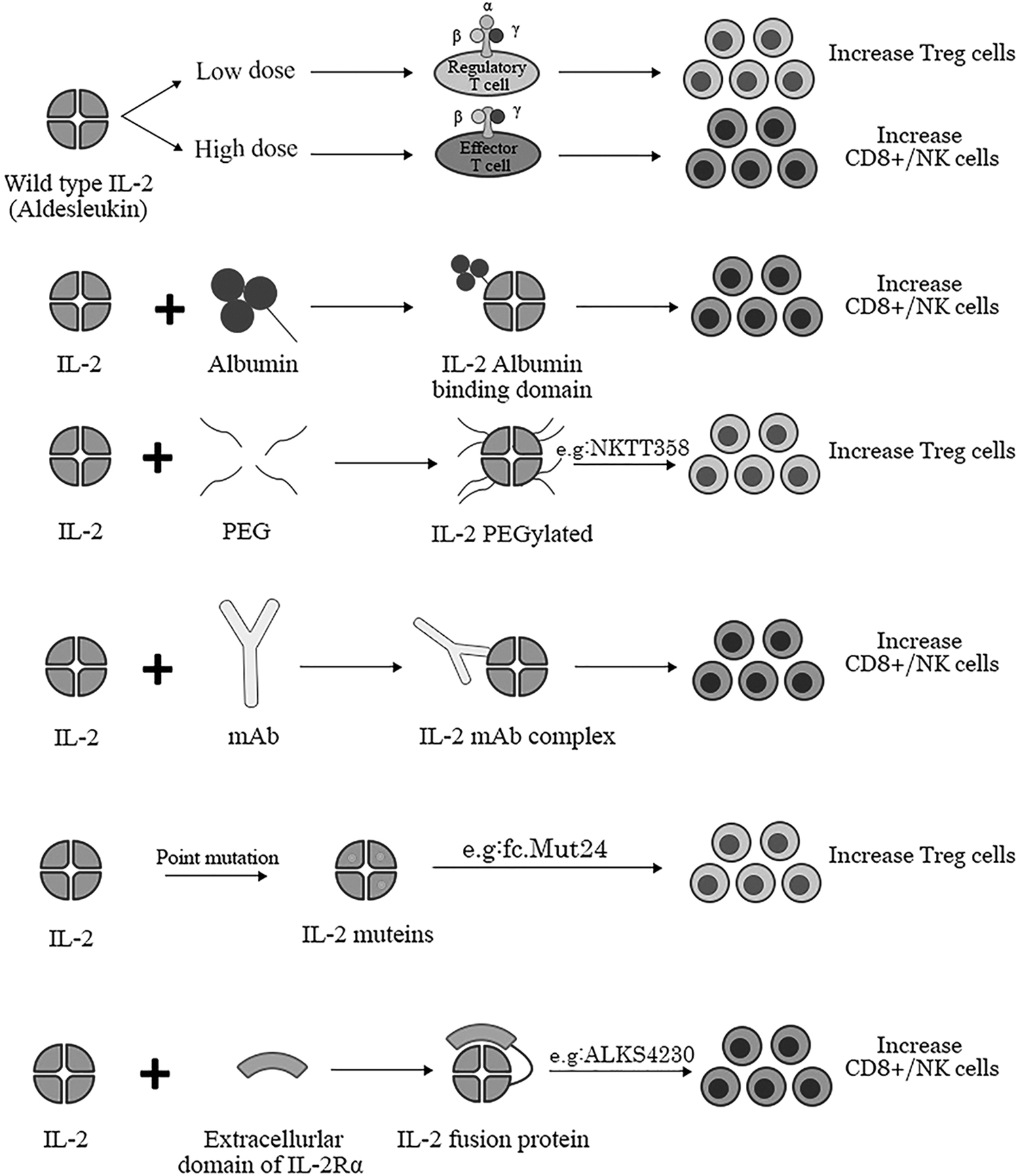
Schematic model of different types of engineering IL-2 and their effects on CD8+ T cells, Tregs, and NK cells proliferation.
Anti-neutralizing IL-2 antibody bound IL-2 elevated the half-life of IL-2 and targeting the high-affinity or intermediate-affinity receptors depending on the antibody structure. In this category, NARA1leukin (ANV419) is the first one in which Aldesleukin is between the K67 and N77 positions, incorporated into the complementarity-determining region 1 (CDR1) of NARA1 anti-IL-2 monoclonal antibody.
In melanoma and breast cancer models, ANV419 demonstrated extended half-life and the ability to abolish CD25 binding, increased the ratio of effector CD8+ T cells to CD4+ Tregs than IL-2/NARA1 complexes. A phase 1 trial in advanced solid tumor patients, the high proliferation rate of CD8+ T cells and minimal increase of Tregs was reported at the MTD of 243 μg/kg with half-life of 12 h (Sahin et al., 2020).
IL-2 muteins composed of 2 classes from the aspect of selectivity for the dimeric (intermediate-affinity IL-2R) or trimeric (high-affinity IL-2R) IL-2 receptors. The first one conferred conformational changes, which avoids CD25 recognition and has greater capacity for activation of CD8+ T cells and NK cells. Conversely the second type by an efficient binding to CD25 preferentially stimulated Treg cells.
In the study by Carmenate et al. (2013), a new IL-2 mutant (no-α-mutein) has been produced with amino acid substitution at positions of R38, F28, Y45, and E62 by alanine. This mutant showed no affinity to IL-2Rα chain and triggered effector function on activated T cell, but not on Tregs. Moreover, in vivo melanoma models showed, the mutein could dampen tumor cells growth and metastasis along with less toxicity as was evaluated by edema and organ lymphocyte infiltration, as compared with wild-type IL-2 (Carmenate et al., 2013).
The antitumor functional of this IL-2 mutant was further confirmed with the ability to increment the rate of memory CD8+ T cells and CD8/Treg ratio in the tumor environment (Carmenate et al., 2022). Swich-2 is an identified IL-2 mutein composed of 5 mutation with the capability to bind α receptor at low PH. This is a PH-sensitive IL-2 variant as it showed high efficacy to bind CD25+ CD8+ active T cells in an acidic PH environment, but not in a neutral one (Gaggero et al., 2022).
This is a novel therapeutic approach based on IL-2 and just being assessed in the preclinical phase. Moreover, the concomitant administration of the engineered IL-2 with other immunotherapy options has been shown to induce more effective antitumor responses. For example, the combination of anti–PD-L1 and Switch-2 treatment controlled established B16.SIY tumors more effective than anti–PD-L1 and IL-2 treatment alone (Gaggero et al., 2022).
Fc.Mut24 is another IL-2-FC mutant containing N103R and V106D point mutations at the contact residues with CD122. The IL-2 mutant showed notable potency to active Tregs but not effector T cells, even at the highest dose. Moreover, in nonobese diabetic mice Fc.Mut24-treated mice exhibited the most sustainable recovery, in contrast to the wild-type IL-2-FC. A number of Fc.Mut24 clinical trials are now underway in SLE, RA, and GVHD (Khoryati et al., 2020).
IL-2-fusion proteins with the extracellular domain of IL-2Rα, FC fragment of Abs or albumin designed to elevate IL-2 half-life and selective interaction with the appropriate cells target. The fusion proteins composed of IL-2 and tumor-specific antibody has been indicated long-term efficacy and safety due to the preferential interplay of IL-2-IL-2R at the tumor site.
CEA-IL-2v (Cergutuzumab amunaleukin) is an IL-2 variant lacking CD25-binding site fused to the anti-carcinoembryonic (CEA) mAb, CEA-IL-2v enhanced CD8+ T and NK cells antitumor responses with only slightly increased number of Tregs in both the peripheral and tumor environment compared with the CEA-IL-2wt (Klein et al., 2017). The outcome in phase I clinical trials of CEA-IL-2v in advanced solid tumors showed the increased frequency of CD8 and NK cells in both tumor environment and periphery (Schellens et al., 2015).
RG-7835 (RO7049665) was designed by fusion of an effector-silenced FC IgG1 to 2 human IL-2 muteins with N88D mutation, IgG-(IL-2N88D)2. The N88D mutation preserved high-affinity binding of IL-2 to the IL-2Rα chain, whereas impeding its binding to the IL-2Rβγ chain. Preclinical evidence has confirmed that RG-7835 selectively activates Tregs, in contrast to Aldesleukin and IgG-(IL-2)2. which also stimulate conventional T cells. According to the pSTAT5 assay results, the RG-7835 treatment on Tregs, CD4 effector T cells, and CD56 bright NK cells indicated, respectively, 6-fold, >300-fold, and >100-fold increase of EC50 compared with the Aldesleukin (Peterson et al., 2018).
Efavaleukin (AMG 592), another IL-2 mutein-FC fusion protein with high-affinity binding to the IL-2Rα has been shown to selectively increase the number of Tregs, slightly elevated Teff but does not affect NK cells. Efavaleukin has been suggested as a better therapeutic option compared with Aldesleukin and is currently under phase I/II clinical trial for SLE, RA, and GVHD patients (Tchao et al., 2017).
Nemvaleukin alpha (ALKS 4230) is composed of circularly permuted IL-2, which is fused to the extracellular domain of IL-2Rα and interacts with the intermediate-affinity IL-2R. Thus, the region bound to the soluble CD25 blocked the IL-2 binding to the IL-2R on the cell surface. This modification could potentially activate NK cells and CD8+ T cells with slight expansion of Tregs. Currently, phase I–III clinical trials are in progress to evaluate the effects of nemvaleukin alpha on solid tumors. According to the available results of a phase I/II trial in patients with melanoma, partial response (PR) was observed in 4 out of 30 patients. Moreover, in 14 patients who were administered both nemvaleukin and pembrolizumab, 2 complete responses and 2 PRs were recorded (Lopes et al., 2020).
In contrast, Ward et al. (2018) designed a mouse IL-2/CD25 fusion protein with the aim of increasing the long-term activity of IL-2 and boosting the response of cytotoxic T cells. Surprisingly, the results showed that the conjugation of IL-2 with CD25 is more effective in expanding Tregs and managing autoimmunity compared with rIL-2 in NOD mice. According to the molecular analysis, the IL-2-CD25 fusion protein formed inactive dimers in trans. In this manner, IL-2 may temporarily dissociate from CD25, making IL-2 accessible to cells expressing IL-2Rα (Ward et al., 2018).
Utilizing pHLIP conjugated to target agents is now considered for the efficient delivery of therapeutic substances across the cell membrane in the acidic environment of tumors. Under acidic conditions, the nature of these peptides changes from being water-soluble to hydrophobic, subsequently integrated into the membrane as an alpha helix (Andreev et al., 2009). Currently, a study has shown that the pHLIP-IL-2 fusion protein enhances IL-2 insertion in tumors acidic environment. As a result, IL-2 could promote the potency of tumor killer cells and effectively reduced tumor size in melanoma and breast tumor models (Chu et al., 2023).
PEGylated IL-2 is composed of 6 polyethylene glycol (PEG) chains that extend the half-life of IL-2 and also change its conformation for preferential binding to the dimeric or trimeric IL-2 receptor. Among the engineered forms of IL-2, bempegaldesleukin (BEMPEG) has been subjected to the highest number of IL-2-based clinical trials in cancer patients. BEMPEG is a PEGylated version of Aldesleukin in which PEG insertion at the interface region of IL-2-IL-2Rα helps in βγ receptor specificity. The latest finding from phase III trials shows that the combination of BEMPEG with nivolumab not only lacks superiority in ORR, PFS, and OR outcome but also presents greater treatment-related toxicity compared with nivolumab alone (Diab et al., 2023).
TransCon IL-2 βγ is a novel IL-2 Rβ/γ selective IL-2 made by constant attachment a small segment of methoxy polyethylene glycol (mPEG) within the IL-2Rα chain binding site. Moreover, to improve the variant half-life and its pharmacokinetic, the TransCon IL-2 βγ was produced as a prodrug using TransCon linker technology (Rosen et al., 2022). In preclinical evaluations, its active biologic form had >30 h half-life, along with robust expansion of CD8+ T cells and NK cells, with just a slight increase in Tregs. In patients with advanced solid tumors, TransCon IL-2 βγ at the recommended dose of 120 mcg/kg alone or in combination with pembrolizumab was well-tolerable and achieved sustained anticytotoxic response with insignificant Tregs expansion (Starodub et al., 2023).
An alternative method to improve IL-2- based therapy is liposome-encapsulated IL-2 mRNA. Delivery of nucleic acid has been optimized by encapsulated lipid nanoparticles for efficient cellular uptake and entrance into the cytosol. In the study by de Picciott et al. (2022) a variant of IL-2 with enhancing IL-2Rα binding-affinity mutants (N88D plus V69A and Q74P) was produced as mRNA encapsulated in lipid nanoparticles. The modified IL-2 showed not only selective stimulation of Tregs, but also increased half-life of the expanded Tregs (de Picciotto et al., 2022).
All of these approaches are designed to increase effector or regulatory T cells based on targeting the dimeric or trimeric IL-2 receptors (Shah et al., 2021). In Table 3, we examine some examples of engineered changes that have been made in recent years on the recombinant IL-2 protein to improve the patient's immune response.
Examples of Engineered Interleukin-2 Recombinant Proteins
BEMPEG, Bempegaldesleukin; UAM, untreated advanced melanoma; CT, clinical trial; UC, ulcerative colitis; HNSCC, head and neck squamous cell carcinoma; CEA, carcinoembryonic antigen; PEG, polyethylene glycol; TransCon, transient conjugate; laIL-2, low-affinity IL-2; PK, pharmacokinetics; PD, pharmacodynamics; ABD, albumin-binding domain; Fc.WT, Fc.Wild type; MP, memory phenotype; LCDR1, light chain complementary determining region1.
Conclusion
IL-2 therapy in patients with cancer and autoimmunity presents a challenge in getting the balance between activating Teffs and Tregs. Therefore, finding the optimal therapeutic dose for each disease has considerable impact to improve the immune response. Besides, hereinafter the immunobiology implications of IL-2 therapy should be clarified in Bregs in preclinical and trial studies.
It is of interest that LD IL-2 treatment in autoimmune patients with infection showed modulation of autoimmune response simultaneously with increased number of antiviral effector T cells (Liang et al., 2021; Zhang et al., 2021). It can be assumed that microbial-specific effector T cells express the high-affinity IL-2R and capable of responding to LD IL-2. LD IL-2 therapy in autoimmune disease is undergoing early phase clinical trials, so it is not possible to draw a decisive conclusion.
HD IL-2 has been suggested for adjuvant treatment of melanoma and RCC patients who are refractory to the immune checkpoint blockers. The reports from multi-IL-2 treatment centers showed good affectivity of HD IL-2 to achieve progression-free survival for >5 years without further systematic therapy in half of the treated patients (Clark et al., 2021). Accordingly, >50% of HD IL-2 recipients failed to get an optimal response.
In addition, HD IL-2 is limited and not offer as a first choice of treatment due to the severe high-grade toxicity in some patients. Engineered IL-2 proteins are considered an attractive option for treatment of cancers or autoimmunity due to the preferential selectivity against target cells. These emerging approaches are still facing obstacles for enhancing therapeutic success rate because of the wide expression of the IL-2 α chain in various stimulatory and regulatory immune cells, for example, Tregs, effector T cells, NK cells, subsets of DC, and endothelial cells (Pol et al., 2020).
Moreover, the CD25 expression doses not necessarily imply the expression of functional IL-2R. For example, conventional DC may present CD25 upon stimulation, but lacks CD122 (Boyman and Sprent, 2012). The off-target effects could be diminished by finding the most potent target-selective IL-2 variant with an effective and safe dose. It should be noted in small amounts of IL-2, Tregs could restrict nearby T cells, but this did not occur in the continuous and strong IL-2 stimulation (Busse et al., 2010). In general, based on the future published results, it will be determined whether the IL-2-based therapeutic options will lead to novel treatments for patients with autoimmunity and cancer.
Footnotes
Acknowledgment
The authors are primarily involved in education and research and did not receive the direct government support.
Authors' Contribution
All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by H.R., G.K., and E.F.-S. The first draft of the article was written by H.R. and N.H. revised it. All authors commented on previous versions of the article. All authors read and approved the final article.
Data Availability Statement
Because of type of this study, data sets have just collected from various recent studies and no data sets were generated.
Author Disclosure Statement
No interests to disclose.
Funding Information
No funds were allocated for the study.