
Abstract
Feijoa sellowiana Berg. fruit, widely used for human consumption, is well appreciated for its good nutritional characteristics and for its pleasant flavor and aroma. In a prior study we showed that the acetonic extract of F. sellowiana fruit exerts a potent antibacterial activity against some Gram-positive and Gram-negative bacterial strains. To confirm and deepen the study on antimicrobial activities of the fruit, in this article we analyze the same extract for its effectiveness against different kind of bacteria both as American Type Culture Collection standard strains and clinically isolates, the Gram-negative Helicobacter pylori, and three fungal strains (one human pathogen and two phytopathogens). Our aim was also to isolate and define the active component(s) involved in the antibacterial, antifungal, and anti-H. pylori activity of the extract through activity-guided fractionation procedures. This allowed us to identify flavone as the active compound of F. sellowiana fruit. Flavone showed a high antibacterial activity against the nine standard bacterial strains tested and the matched clinically isolated bacterial strains and was significantly more active against H. pylori than metronidazole. Pseudomonas aeruginosa, Proteus mirabilis, and Proteus vulgaris from both standard and clinical sources showed a very high sensitivity to flavone (minimal inhibitory concentration [MIC] = 1.95 μg/mL for standard strains and 3.9 μg/mL for clinical isolates of the three bacteria), showing also good minimal bactericidal concentration values. Among fungi tested, Rhyzoctonia solani was the most sensitive strain to the action of flavone (MIC = 62.5 μg/mL).
Introduction
F
Different biological activities of F. sellowiana have been reported. In previous studies we showed the antioxidant activity of the aqueous extract of the fruit 11,12 and strong activity against different bacteria of the acetonic and aqueous extracts of leaf, stem, and different parts of fruit. 11,13 In addition, F. sellowiana peel contains promising levels of multidrug resistance-modulating and cytotoxic activities. 14 We have also evaluated the anti-inflammatory activity of an acetonic extract from F. sellowiana fruits on the nitric oxide (NO) pathway, demonstrating that the extract is able to inhibit NO production because of the presence of flavone and stearic acid. 15 Finally, we showed that the same acetonic extract exerts anticancer activities on solid and hematological cancer cells, displaying a tumor-selective activity. Fractionation and subsequent purification identified flavone as the active component of the extract. 16
To deepen our understanding of the various biological activities shown by the acetonic extract of F. sellowiana fruit and in an attempt to contribute to its use as alternative products for microbial control and food preservation, in this study we analyze the inhibitory effect of the extract against two Gram-positive and seven Gram-negative bacterial strains, both American Type Culture Collection (ATCC) standard strains and clinical isolates (CIs). A large number of Gram-negative bacteria have been chosen for our study because resistance of these bacteria against the more used antibiotics is common. We have also evaluated the effectiveness of the extract against three fungal strains and Helicobacter pylori. Because current medical treatments of H. pylori have had undesirable effects on nontarget organisms such as intestinal microorganisms and have sometimes caused serious side effects, 17 –20 actually there is a strong demand for compositions having all of the beneficial properties of current therapy agents but with fewer side effects. A number of natural substances used for food such as garlic extracts, peas, oregano, cranberry, cumin, propolis, and olive oil have been shown to have antibacterial effects against H. pylori in vitro. 21 Given the potent antimicrobial activity of F. sellowiana acetonic extract against various Gram-positive and Gram-negative bacterial strains, 13 it was suspected that this activity could also be exerted also against H. pylori and fungi. Our aim was also the identification of the substance(s) responsible for the antibacterial, antifungal, and anti-H. pylori activities of F. sellowiana fruit, and to this purpose we have defined a fractionation procedure.
Materials and Methods
Plant material
F. sellowiana fruits were collected in the Botanical Garden of Naples, Naples, Italy. The identification was done by one of us (A.B., Section of Plant Biology, Department of Biological Sciences, University “Federico II,” Naples). A voucher specimen (NAP number 96-125) is deposited at the Herbarium Neapolitanum, Department of Biological Sciences, University “Federico II.”
Extraction and isolation of active compounds
Six hundred ninety grams of whole F. sellowiana fruits were blended, freeze-dried, and then extracted for 3 days with acetone at room temperature (3 × 5 L). After filtration, the solvent was evaporated under reduced pressure and moderate temperature (35°C) to give a gum (46.45 g). An amount of the extract (5.8 g) was dissolved in methanol and then subjected to column chromatography on silica gel 60 (Merck, Darmstadt, Germany) (4 × 100 cm, 300 g, 70–230 mesh). Elution with n-hexane/ethyl acetate (from 100:0 to 0:100 stepwise gradient) and successively methanol afforded 11 fractions (A–M) of 100 mL each, gathered according to thin-layer chromatography analysis [eluent system n-hexane/ethyl acetate (1:1, vol/vol), spray reagent Ce(SO4)2 in H2SO4], that were successively subjected to biological testing: fraction A (57 mg, eluted with 80:20 n-hexane/ethyl acetate), B (35.6 mg, eluted with 60:40 n-hexane/ethyl acetate), C (30.1 mg, eluted with 50:50 n-hexane/ethyl acetate), D (28.4 mg, eluted with 40:60 n-hexane/ethyl acetate), E (18.2 mg, eluted with 30:70 n-hexane/ethyl acetate), F (22 mg, eluted with 100% ethyl acetate), G (31.4 mg, eluted with 90:10 ethyl acetate/methanol), H (457 mg, eluted with 70:30 ethyl acetate/methanol), I (1.4 g, eluted with 60:40 ethyl acetate/methanol), L (410 mg, eluted with 10:90 ethyl acetate/methanol), and M (58.2 mg, eluted with 100% methanol). Fraction B, which was shown to be the most active (data not shown), was further purified by direct-phase high-performance liquid chromatography on a silica column, performed with a TSP (ThermoQuest, San Jose, CA, USA) Spectra Series P 100 pump equipped with a Rheodyne (Cotati, CA) injector, a refractive index detector, and a Hypersil silica column (250 × 4.6 mm; flow rate, 1.5 mL/minute) (Thermo, Waltham, MA, USA). The purification of fraction B (n-hexane/ethyl acetate 45:55 as eluent) yielded pure flavone (30 mg). The structure of this compound was determined by comparison of its spectroscopic data (ultraviolet, nuclear magnetic resonance, and mass spectrometry) with literature values 22 : mass spectrometry, [M–H]− = 221; ultraviolet, λ max (methanol) (log ɛ) 250 (4.05), 296 (4.22); nuclear magnetic resonance, see Blasko. 22
Antimicrobial activity assays
Microorganisms
Nine bacterial strains from the ATCC (Rockville, MD, USA) were used. They included the Gram-positive bacteria Staphylococcus aureus (ATCC 13709) and Enterococcus faecalis (ATCC 14428) and the Gram-negative bacteria Proteus mirabilis (ATCC 7002), Proteus vulgaris (ATCC 12454), Pseudomonas aeruginosa (ATCC 27853), Salmonella typhi (ATCC 19430), Enterobacter aerogenes (ATCC 13048), Enterobacter cloacae (ATCC 10699), and Klebsiella pneumoniae (ATCC 27736). The same bacterial strains as CIs were used to compare the sensitivity to fractions and flavone.
H. pylori (CCUG strain) was kindly provided by the Cellular and Molecular Biology and Pathology Department, University Federico II.
Fungi
Antifungal tests were done on three strains of fungi, including one potentially pathogenic yeast (Candida albicans CI) and two filamentous phytopathogenic fungi, Botrytis cinerea and Rhyzoctonia solani, kindly provided by the Plant Pathology Department of the University Federico II.
Preparation of the fractions and pure substance for antibacterial assay
The fractions of the acetonic extract (A–M) were centrifuged at 2,800 g, and the supernatants were oven-dried at 45°C. The samples were prepared according to the method used by Ieven et al. 23 in which 100 mg of the dry residues were dissolved in 10 mL of sterile physiological Tris buffer (pH 7.4) and tested for antimicrobial activities from 1 mg/mL to 10−4 mg/mL, at 10-fold dilution. The fractions and pure substance were added with 5 × 10−2 M stock solution in dimethyl sulfoxide. They were diluted from 10−2 to 1,000 μg/mL concentrations in sterile physiological Tris buffer (pH 7.4, 0.05 M) immediately before being used. 23 The dose-dependent inhibition found (up to 10−3 mg/mL) led us to isolate and test the active fraction and the active substance.
Minimal inhibitory concentration (MIC) and minimal bactericidal concentration (MBC) determination
Bacterial strains were grown on Mueller-Hinton agar plates (Difco, Detroit, MI, USA) and suspended in Mueller-Hinton broth (Difco). The MIC values against bacterial strains were performed using the broth-dilution method (Mueller-Hinton broth) of Ericcson and Sherris. 24 The inoculum suspensions were prepared from 6-hour broth cultures and adjusted to obtain a 0.5 McFarland standard turbidity. The F. sellowiana acetonic extract and the fractions were sterilized by filtration through Millipore (Bedford, MA, USA) filters (pore size, 0.45 μm) and added to Mueller-Hinton broth medium. Serial 10-fold dilutions were made for a concentration range between 0.01 and 1,000 μg/mL for the samples. In the range between the minimum active and the maximum inactive concentrations were tested twofold dilutions of the samples to obtain a more precise measure of the MIC. The bacterial suspensions were aerobically incubated for 24 hours at 37°C. The MIC was defined as the lowest concentration able to inhibit any visible bacterial growth. Cultures containing only sterile physiological Tris buffer (pH 7.4, 0.05 M), which did not influence bacterial growth, were used as controls. The MIC values were also determined for tetracycline hydrochloride (Pharmacia, Milan, Italy), benzyl penicillin sodium (Cynamid, Catania, Italy), and cefotaxime sodium (Roussel Pharma, Milan) in Mueller-Hinton broth using a standard method.
The MBC determination was carried out by transferring to the fresh Mueller-Hinton broth aliquots of bacterial suspensions from the test tubes containing samples concentrations equal or higher (up to 1,000 μg/mL) than the MIC. Each sample was tested in triplicate, and the experiment was performed four times.
Anti-H. pylori activity
H. pylori was inoculated on a Brucella agar plate containing 10% sterile defibrinated sheep blood, 5 mL of Vitox (OXOID, Cambridge, UK), and 2 mL of Skirrow (BD Diagnostic Systems, Heidelberg, Germany) and cultured at 37°C for 72 hours. The bacterial colonies were collected and diluted to 107 colony-forming units/mL with 0.9% NaCl solution. The fractions and pure substance were diluted with dimethyl sulfoxide and then added to the liquid culture medium. The culturing medium was made of 14 g of Brucella broth, 1.25 mg of FeCl3 · 6H2O, 500 mL of double-distilled H2O, 25 mL of fetal calf serum, 5 mL of Vitox, and 2 mL of Skirrow. Each bacterial suspension was added to the culturing medium obtaining a well density of 106 colony-forming units/100 mL. The mixture was incubated at 37°C for 72 hours. 25,26 Control cultures were made by culturing the bacterium without and with metronidazole at 0.5 μg/mL.
Antifungal activity
The fungi were maintained on Sabouraud glucose agar medium (Diagnostics Pasteur, Paris, France).
Two methods were used for the in vitro assays: (1) The preliminary screening for antifungal activity was done by the method of agar incorporation (dilution on a solid medium) described by Taudou
27
and Dwivedy and Dubey.
28
The strains were cultivated on Sabouraud glucose agar medium in 3-cm-diameter Petri dishes. One hundred microliters of each extract solution was aseptically mixed with 1.9 mL of Sabouraud glucose agar (liquified and maintained at the melting point in a water bath at 45°C) to give final concentrations of 1, 2, and 4 mg/mL. The solvent of each extract was used as a negative control. After cooling and solidification, the seeding was carried out by a double inoculation in the middle of the Petri dish with a small amount of a 7-day-old mycelium culture of fungi to be tested. The Petri dishes were then incubated at 25°C for 7 days. Three replicates for each concentration and microorganism were carried out. The antifungal activity of the all fractions and flavone was evaluated according to the method of Singh et al.
29
by calculating the percentage inhibition (I%) from the diameter values of the colony in the control plate (dC) and the colonies in the plates added with the assayed extracts (dE): I% = (dC − dE)/dC. (2) The MIC that inhibits the visible growth of fungi was determined by the standardized broth microdilution method of Torres-Rodriguez et al.
30
using Sabouraud glucose broth (Diagnostics Pasteur). It was determined for fractions and flavone selected by the incorporation method. A volume of 850 mL of Sabouraud glucose broth, 100 mL of fractions and flavone previously diluted in decreasing concentrations (a serial twofold dilution ranging in concentration from 1 to 0.125 mg/mL), and 50 mL of a suspension of spores (C. albicans and B. cinerea) or mycelium (R. solani) were introduced into each well of a sterile 24-well flat-bottom microplate with a lid, except for the negative control wells, which contained solvent or distilled and sterilized water. Inocula strains of fungi were prepared by suspending spores in sterilized distilled water from 72-hour- and 7-day-old cultures, respectively, on Sabouraud glucose agar to a final concentration of 4 × 103 spores/mL. The microplates were incubated for 7 days at 25°C with slow shaking (100–200 rpm). Growth was observed each day from day 2 to day 7. The MIC was thus determined every day. Each test was repeated at least three times. The minimal fungicidal concentration (MFC) was measured according to the method of Rotimi et al.
31
For confirmation of the fungistatic or fungicidal activity, all wells showing no visible growth after 7 days were subcultured onto Sabouraud agar or broth using an inoculum of 50 mL and incubated at 25°C for 10 days. The MFC was recorded as the lowest concentration that did not yield growth. Positive control cultures were made culturing the fungi without and with ketoconazole at 0.2 μg/mL.
Results
Antibacterial and anti-H. pylori activity
Acetonic extract of F. sellowiana fruit was analyzed for its activity against the bacterial strains listed in Table 1 and against the same bacteria clinically isolated, all pathogens for humans and known to cause respiratory, gastrointestinal, skin, and urinary disorders. Results are shown in Table 1. Antibacterial activity with standard antibiotics showed combined resistance to cefotaxime and benzyl penicillin sodium in four CIs, combined resistance to cefotaxime and tetracycline in two standard ATCC strains and four CIs, combined resistance to benzyl penicillin sodium and tetracycline in eight CIs, and combined resistance to all three antibiotics in four CIs (E. aerogenes, E. cloacae, E. faecalis, and S. aureus) (Table 1).
CTAX, cefotaxime; PENG, benzyl penicillin sodium; R, resistant; TET, tetracycline.
The activity of the extract against the tested bacteria was high, with MICs between 3.9 and 31.3 μg/mL for ATCC standards and 7.8 and 62.5 μg/mL for CIs. Chromatographic procedures on whole acetonic extract of F. sellowiana afforded 11 fractions (A–M, see Materials and Methods) that were all tested for their antibacterial activity against the same bacterial strains. Antibacterial activity was remarkable in only one of the fractions tested (fraction B), whereas it was negligible in the other fractions studied (data not shown). Purification of the active fraction by direct high-performance liquid chromatography yielded pure flavone as the main component. Pure flavone isolated from the fraction was therefore tested for its action against the bacterial strains listed in Table 1: the activity of flavone against the tested bacteria was generally high (MICs between 1.95 and 15.6 μg/mL for ATCC standards and 3.9 and 31.3 μg/mL for CIs). E. aerogenes and E. cloacae, both ATTC standards and CIs, showed the lowest sensitivity to the F. sellowiana extract and flavone, which were active towards S. aureus, E. faecalis, S. typhi, and K. pneumoniae. The other Gram-negative strains were instead strongly inhibited by extract and flavone: in particular, P. aeruginosa, P. mirabilis, and P. vulgaris from both standard and clinical sources showed a very high sensitivity to extract and flavone, showing also good MBC values. P. aeruginosa showed a MIC even lower than standard antibiotics.
Antibacterial activity of F. sellowiana acetonic extract against H. pylori is shown in Figure 1. We found a significant in vitro effect of plant extract against this bacterium. Among all the fractions tested, fraction B showed the highest activity (data not shown). Subsequently an aliquot of flavone isolated from the active fraction was assayed for its activity against H. pylori; as we can see in Figure 2, flavone showed high activity and was significantly more active than metronidazole.
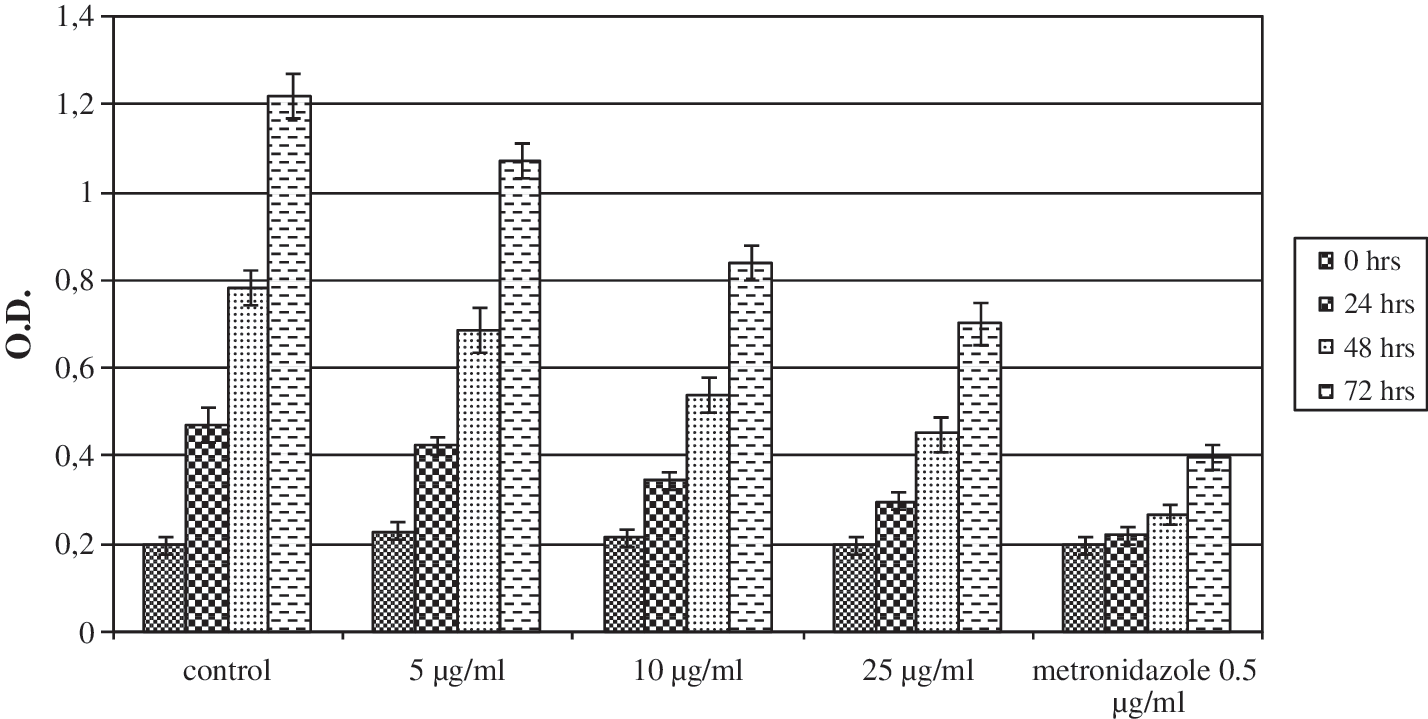
Antibacterial activity of F. sellowiana acetonic extract against H. pylori growth expressed as optical density (O.D.) at 450 nm. Control cultures were made by culturing the bacterium without and with metronidazole.

Antibacterial activity of flavone from F. sellowiana against H. pylori growth expressed as optical density (O.D.) at 450 nm. Control cultures were made by culturing the bacterium without and with metronidazole.
Antifungal activity in solid medium
We have studied the antifungal activity of the acetonic extract of F. sellowiana fruit and of fractions A–M against three strains of fungi, including one potentially pathogenic yeast (C. albicans CI) and two filamentous phytopathogenic fungi (B. cinerea and R. solani). The results of the assays with dilution on solid medium are not shown. Only whole acetonic extract, fraction B, and pure flavone isolated from it, at the three concentrations tested, inhibited the three fungi: in particular, R. solani from 0% to 50% and C. albicans and B. cinerea from 0% and 25%.
Antifungal activity in liquid medium
Because only the samples with a fungal growth inhibition superior or equal to 50% (high) were the object of MIC determination, only whole acetonic extract, fraction B, and flavone were successively tested. Results for acetonic extract and flavone are shown in Table 2 and indicate that the samples had a fungistatic effect on all the fungi tested, with MICs ranging from 63 and 250 μg/mL. R. solani was the strain most sensitive to the extract (MIC = 123 μg/mL) and above all to flavone (MIC = 62.5 μg/mL). C. albicans and B. cinerea were less sensitive to the action of both extract (MIC = 250 μg/mL) and flavone (MIC = 123 μg/mL).
KCON, ketoconazole. A dash indicates not active.
Discussion
It has been well recognized that plant extracts and phytochemicals could be developed into products suitable for bacteria, fungi, and H. pylori eradication because many of them are selective, often biodegrade to nontoxic products, and may be applied to humans in the same way as other conventional chemical drugs. 32 In a previous study we have discovered a very high antimicrobial activity of F. sellowiana acetonic extract. 13 This finding led us to examine its activity against a broad spectrum of bacterial pathogens for humans and known to cause respiratory, gastrointestinal, skin, and urinary disorders, against the same bacteria clinically isolated, some of which showed resistance to the standard antibiotics, H. pylori, and three fungal strains, one potentially pathogenic yeast (C. albicans CI), and two filamentous phytopathogenic fungi (B. cinerea and R. solani).
All the bacteria tested showed sensitivity to the extract (Table 1), and the compound responsible for this activity seems to be flavone. P. aeruginosa, P. mirabilis, and P. vulgaris from both standard and clinical sources were the most sensitive to the action of the extract and flavone. The activity of F. sellowiana acetonic extract and flavone mainly against Gram-negative bacteria has remarkable importance; in particular, the most significant datum is the effectiveness of the extract and flavone against P. aeruginosa and S. aureus, insofar as infections due to these bacteria are particularly problematic because of their intrinsic resistance to multiple classes of antibiotics and their ability to acquire adaptive resistance during a therapeutic course. 33
The in vitro effect of plant extract against H. pylori (Fig. 1) and its antifungal activity (Table 2) were also significant, and we showed that these actions seem to be due to the presence of flavone too (Table 2 and Fig. 2). Interestingly, previously we demonstrated that flavone seems to be the compound responsible for the anti-inflammatory and antitumor activities shown by F. sellowiana fruit. 15,16
Flavonoids and flavonoid glycosides have always been a component of human diet, with no harmful side effects even at high dosages, and they are thought to be chemically stable under the conditions of the human stomach. 34 Flavonoids are well known for their antibacterial activity against different Gram-positive and Gram-negative bacterial strains; interestingly, most of the active flavonoids were found in flavones and flavonols, whose A/C rings take planar structures, such as flavone. 35 Literature shows also that flavonoids could have a therapeutic potential for treatment of gastrointestinal diseases associated with H. pylori infection. In fact, anti-Helicobacter activity of different flavonoid derivatives and their metabolites, such as genistein, hesperitin, ponciretin, irisolidone, and cabreuvin, was reported in several studies 34,36,37 ; for these reasons it was expected that the antimicrobial activity of F. sellowiana would be related to this class of compounds. Our finding that flavone is the compound responsible for the anti-Helicobacter action of F. sellowiana is in addition confirmed by a previous study, 38 in which an inhibitory action of flavone on H+/K+-ATPase activity was shown, along with its capacity to stimulate prostaglandin E2 production in isolated gastric mucosal cells.
The antifungal activity showed by F. sellowiana fruit and flavone is an important finding too, as C. albicans is a causal agent of opportunistic oral and genital infections in humans, responsible for most cases of yeast infection. Our study shows also that F. sellowiana and its main compound flavone could potentially be used against fungal diseases of plants, as they show a good activity against R. solani and B. cinerea, two filamentous phytopathogenic fungi that infect a great variety of plants of economic importance. The effectiveness of flavone against different fungal strains was demonstrated before, 39 and our results support these findings.
We have to remark, however, that most likely the biological activities of F. sellowiana acetonic extract could be due to a possible synergic effect of the different compounds contained in the F. sellowiana fruit. In fact, F. sellowiana is known to contain high amounts of compounds that are presumed to contribute to its antimicrobial activity such as polyphenols, flavonoids, and tannins 6 –8 whose antibacterial and antifungal activities have been shown before. 34 –39 Furthermore, the vigorous and effective activity against the microorganisms tested may be also sustained by the high iodine content in the plant, as compounds containing iodine are known to be active against bacteria in vegetative form, yeast, and viruses. 13
In conclusion, this article confirms that flavonoids from natural sources, and particularly flavone, could be considered a valuable support in the treatment of infections and may contribute to the development of new and safe agents for inclusion in antimicrobial, antifungal, and anti-H. pylori regimens. Plant extracts as antimicrobial agents present two main positive features: the first is their natural origin, which means more safety for consumers, and the second is that they are considered to be low risk for resistance development by pathogenic microorganisms.
Footnotes
Author Disclosure Statement
The authors declare that no competing financial interests exist for all the authors of the manuscript.