
Abstract
Bilberry (European blueberry) has been reported to have many biological effects, including anticancer activity. In this study, we investigated the antiproliferative effects of bilberry extract in relation to its ability to induce apoptosis and affect microtubule assembly and organization in MCF7 human breast cancer cells. We observed that bilberry extract inhibited cell proliferation in a concentration-dependent fashion with a 50% inhibitory concentration of 0.3–0.4 mg/mL, in concert with induction of apoptotic cell death. At these concentrations there was no selective inhibition of mitosis or any other cell cycle stage, nor was there any apparent effect on the microtubule or actin cytoskeletons. However, somewhat higher extract concentrations (0.5–0.9 mg/mL) did cause an increase in the fraction of cells at the G2/M phase of the cell cycle, together with destruction of microtubules and formation of punctate tubulin aggregates in the cells. Bilberry extract at 0.3–0.4 mg/mL did not appreciably inhibit microtubule polymerization in vitro, but significant inhibition of polymerization (∼30%) did occur at higher extract concentrations (0.5–1 mg/mL). We conclude that bilberry extract as ingested by humans, not just the purified anthocyanins it contains, inhibits proliferation of and induces apoptosis in breast cancer cells at its lowest effective concentrations via a mechanism that does not involve action on microtubules or on mitosis. We further conclude that at somewhat higher concentrations the extract modifies microtubule organization in cells and causes accumulation of cells at mitosis by a direct action on microtubules.
Introduction
A
Because consumption of berries is the predominant means of anthocyanin ingestion, this means human exposure to these putative anticancer agents is via a mixture of several anthocyanins as well as other berry molecular components. Given the potential for synergistic action of mixtures of anthocyanins 17 and that these anthocyanins are typically consumed through diet in the form of berries, we wanted to analyze the cytotoxic effects of a full extract from bilberries.
One of the major mechanisms of cancer suppression involves induction of apoptosis. 18,19 Bilberry extract has been shown to induce apoptotic cell bodies and nucleosomal DNA fragmentation in human leukemia HL60 cells 8 and in human colon cancer HT-29 cells. 20 To study bilberry extract's ability to inhibit proliferation in association with induction of apoptosis, we used a sulforhodamine B cell proliferation assay together with terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) and single-stranded DNA (ssDNA) apoptosis assays. In addition, to examine the cytotoxic effects of bilberry extract, we reasoned that evaluation of cellular microtubules would also be an important tool. Microtubules are highly dynamic cytoskeletal fibers composed of tubulin subunits that are important in the process of mitosis, during which the duplicated chromosomes of a cell are separated into two identical sets before cleavage of the cell into two daughter cells. Their involvement in mitosis and cell division makes microtubules an important target for anticancer drugs. 21 They are also important in organization of the cytoplasm. Thus in this study, we used MCF7 human breast cancer cells stably transfected with green fluorescent protein (GFP)/α-tubulin (MCF7-GFP-tubulin cells), which have been used as an experimental model to evaluate effects of antimitotic agents due to their sensitivity to microtubule-targeted antimitotic drugs. 22,23
Materials and Methods
Materials
All chemicals and reagents used in this work were purchased from Sigma (St. Louis, MO, USA) unless otherwise specified. The capsulated form of a standardized preparation of bilberry extract (Nature's Way, Springville, UT, USA) was used as the source of bilberry. In a typical extraction procedure, the content of three capsules was sequentially extracted 10 times with 70% ethanol per extraction step followed by a single extraction step with 100% ethanol as described below. A total of 50 mL of 70% ethanol was used to extract the contents of bilberry extract capsules. For each extraction step, 5 mL of 70% ethanol was added to the capsule content, the extraction mixture was mixed on a vortex-mixer for 1 minute, and the undissolved matter was collected as a pellet by centrifugation for 15 minutes at 12,000 g. For the last extraction cycle, 5 mL of 100% ethanol was added to the pellet, vortex-mixed for 1 minute, and incubated overnight (at −5°C), and the undissolved matter was pelleted by centrifugation for 15 minutes at 12,000 g. Solvent was evacuated from combined supernatants from all extraction cycles under vacuum on a Rotovac® rotary evaporation system (model R-124, Büchi Labortechnik AG, Flawil, Switzerland), and the dry substance was dissolved in 70% ethanol at a concentration of 50 mg/mL. This preparation of bilberry extract was used as a stock source for all experiments and stored at −20°C. Although the stability was not examined systematically, there was no apparent decrease in biological activity over 18 months.
Cell culture
MCF7 cells (ATCC number HTB-22, American Type Culture Collection, Rockville, MD, USA) stably transfected with GFP and α-tubulin (MCF7-GFP-tubulin cells), which exhibit high sensitivity to microtubule-targeted antimitotic drugs, 22,23 were grown in HEPES-fortified high-glucose Dulbecco's Modified Eagle's medium (DMEM) (catalog number D1152) supplemented with 10% fetal bovine serum (Atlanta Biological, Atlanta, GA, USA), 1× Minimum Essential Medium nonessential amino acid solution, and sodium bicarbonate at 37°C in a humidified atmosphere containing 95% air and 5% CO2. 24,25 Culture media were changed every 3–4 days, and doubling time was 36 hours.
Cell proliferation assay
MCF7-GFP-tubulin cells were seeded at a density of 2 × 105/mL (0.1 mL per well) in 96-well microtiter plates and allowed to attach for 24 hours to keep the cells in log-phase growth at the time of drug treatment. Bilberry extract was dissolved in 70% ethanol and administered in DMEM supplemented with 10% fetal bovine serum (final concentrations of ethanol were at or below 1.4%). When 1.4% of 70% ethanol alone was added to the vehicle control wells, it did not affect proliferation of the cells. After incubation with the indicated drug concentrations for 72 hours, cell proliferation was determined using a sulforhodamine B assay. 26 In the cell proliferation and cell cycle experiments, the ethanol concentration in all samples, including controls, was 1.4%. All other experiments reported here used an ethanol concentration of 0.7%.
Cell cycle analysis
To investigate cell cycle distribution, 5 × 104 cells were plated in 24-well plates, and, after a 24-hour recovery period, they were treated with vehicle alone or bilberry extract for 24 hours. After treatment, cells were collected by trypsinization, fixed with 70% ice-cold ethanol, and washed in phosphate-buffered saline. Fixed cells were stained with Guava Cell Cycle Reagent containing propidium iodide (Guava Technologies, Hayward, CA, USA) in the dark, at room temperature for 30 minutes, according to the manufacturer's directions. The propidium iodide-stained DNA fluorescence was measured using a Guava EasyCyte™ flow cytometer (Guava Technologies). The populations of G0/G1, S, and G2/M cells were calculated using ModFit LT cell cycle analysis software (Verity Software House, Topsham, ME, USA).
Immunofluorescence microscopy
MCF7-GFP-tubulin cells were prepared for immunofluorescence microscopy by formalin/methanol fixation as previously described. 22,23 In brief, MCF7-GFP-tubulin cells were incubated with bilberry extract at the indicated concentrations for 24 hours. Immunofluorescence microscopy of microtubules and DNA staining were performed as follows: fixed cells were first stained with a mouse monoclonal anti-α-tubulin antibody (DM1A; 1:1,000 dilution), followed by staining with fluorescein isothiocyanate-conjugated anti-mouse secondary antibody to stain the microtubules and by rhodamine-conjugated phalloidin (Invitrogen, Carlsbad, CA, USA) to stain actin filaments. Cells were mounted to the microscope slides with 4′,6-diamidino-2-phenylindole-containing ProLong® Gold (Invitrogen) medium. Microscopy was performed on a spinning disk confocal microscope (model IX81 DSU, Olympus, Tokyo, Japan). Images were taken with an ImagEM camera (Hamamatsu, Shizuoka, Japan) under the control of SlideBook software (Olympus).
Microtubule polymerization
Microtubule polymerization was carried out as described previously 27 beginning with a solution of tubulin and microtubule-associated proteins purified from bovine brain. Microtubules (17 μM tubulin) and various concentrations of bilberry extract were polymerized at 30°C for 75 minutes in assembly buffer containing 2.0 mM GTP. The amount of protein in microtubule pellets was determined after sedimentation and resuspending the microtubules in ice-cold buffer.
Apoptosis measurements
The percentage of apoptotic cells was measured in two ways: by using a commercial Guava TUNEL kit (Millipore, Bedford, MA, USA) and by using an Apo ssDNA kit (Cell Technology Inc., Mountain View, CA, USA).
For the Apo ssDNA Assay, MCF7 cells were seeded at 1 × 106 cells/mL in T25 flasks with 5 mL of DMEM supplemented with 10% fetal bovine serum and allowed to recover for 24 hours at 37°C. Apoptosis was induced by bilberry extract in concentrations of 0.125, 0.25, and 0.5 mg/mL for 24 hours and with 10 μM camptothecin (positive control) for 6 hours. Cells were collected and fixed with methanol, and DNA was denatured in a water bath at 80°C for 10 minutes. Cells were blocked for 30 minutes in 1% bovine serum albumin in phosphate-buffered saline. Cells were further stained with mouse anti-ssDNA (primary antibody) and goat anti-mouse immunoglobulin M fluorescein isothiocyanate-labeled antibody (secondary antibody) in accordance with the manufacturer's protocol. The samples were analyzed using a Guava EasyCyte flow cytometer, and the results analyzed with FCS version 3.0 Express (De Novo Software, Los Angeles, CA, USA).
For the TUNEL assay, cells were seeded at 20 × 104 cells per well in 2 mL (10 × 104 cells/mL) in a six-well plate and given 24 hours of recovery time before adding extract. Cells incubated with extract for the desired time span were collected, fixed in 3.7% formaldehyde solution, and stored in 70% ethanol for > 12 hours at −20°C. Just before analysis, cells were rinsed and treated with terminal deoxynucleotidyl transferase in the presence of bromo-deoxyUTP to introduce bromodeoxyuridine tails at exposed 3′-hydroxyl ends of DNA. The relative number of 3′-hydroxyl ends of DNA was assessed by painting cells with tetramethyl rhodamine isothiocyanate-conjugated anti-bromodeoxyuridine monoclonal antibody over a Guava EasyCyte flow cytometer.
Statistics
Results are reported as average values of at least three independent experiments. Means of individual groups were converted to a percentage of the control value and expressed as the mean ± SD. The difference between groups was assessed using Student's t test for unpaired samples. A value of P < .05 was considered as significantly different.
Results and Discussion
Bilberry extract inhibited proliferation of MCF7-GFP-tubulin cells in a concentration-dependent fashion (Fig. 1). There was a steep concentration dependence for this antiproliferative effect, with a large increase between 0.25 and 0.5 mg/mL extract. As compared with the control cells (no extract), inhibition of proliferation of extract-treated cultures was 7 ± 10% with 0.125 mg/mL bilberry extract, 11 ± 9% with 0.25 mg/mL, 66 ± 10% with 0.5 mg/mL, and 75 ± 10% with 1.0 mg/mL, with an IC50 of approximately 0.3–0.4 mg/mL (Fig. 1). This concentration dependence of the extract is consistent with reports on the antiproliferative effects of purified anthocyanins on several other cancer cell lines. 1,17,28 –30 Additionally, the sub-mg/mL levels of bilberry needed to elicit this effect is lower than that reported on another cancer cell line (HT-29 colon cancer cells), where 10 mg/mL was needed to produce a 30% reduction in cell proliferation. 20
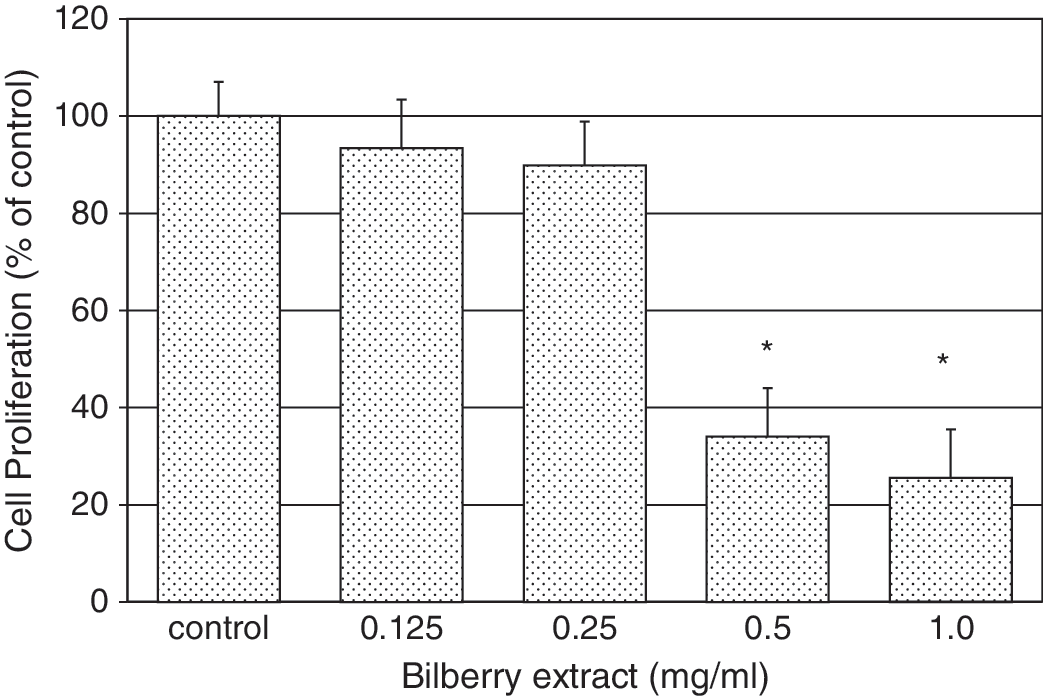
Effect of bilberry extract on proliferation of MCF7-GFP-tubulin cells. Cells were incubated for 72 hours in the presence of varying concentrations of bilberry extract. Cell viability was determined using the cell proliferation assay described in Materials and Methods. Each bar represents the mean ± SD of three independent experiments in comparison with that of the control set to 100%. Significant inhibition of proliferation (*P < .05) occurred with bilberry extract concentrations in excess of 0.25 mg/mL, with an IC50 of 0.43 mg/mL.
We examined whether bilberry extract affects the cell cycle of MCF7-GFP-tubulin cells (Fig. 2). The populations of cells in G0/G1, G2/M, and S phase after treatment with low concentrations of bilberry extract (0.3 mg/mL) were the same as controls, but at higher concentrations of extract, the G2/M population increased, and the G0/G1 cell population decreased. This was reflected in the values observed at 0.6 mg/mL compared to 0.9 mg/mL: the fraction of cells in G0/G1 was reduced from 48% to 35%, those in G2/M increased somewhat from 24% to 32%, and those in S increased from 28% to 33%. Thus we observed a decrease of cells in the G0/G1 phase and a small increase in the G2/M phase at extract concentrations greater than the IC50. This indicated that bilberry extract at least partially arrested the cell cycle at the G2/M phase.
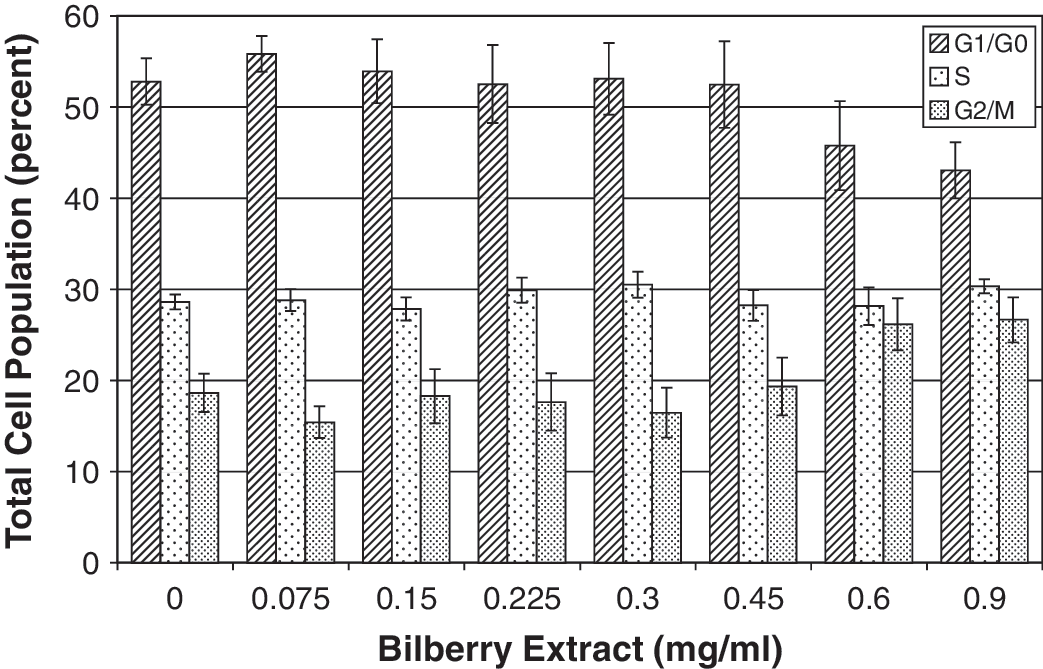
Effect of bilberry extract on cell cycle of MCF7-GFP-tubulin cells. Cell cycle distribution after a 24-hour incubation with bilberry extract was determined in samples stained with propidium iodide and measured by flow cytometry. Relative to the control, more cells were seen in G2/M phase as the concentration of the bilberry extract was increased, indicating a partial block in this phase of the cell cycle, with fewer cells progressing to G0/G1 at higher bilberry concentrations. Data shown represent the average and SE of three cell cycle experiments.
We next wanted to determine whether inhibiting proliferation was associated with apoptosis. We observed a substantial induction of maximal apoptosis at extract con-centrations of 0.25 mg/mL, near the IC50 for inhibition of proliferation.
We used an Apo ssDNA assay, observing a bilberry extract induction of apoptosis in the MCF7 cells within 24 hours (Fig. 3). The Apo ssDNA assay takes advantage of the fact that chromatin of apoptotic cells is more susceptible to denaturation at lower temperatures than healthy cells. We observed an increase in the percentage of apoptotic cells at a lower IC50 inhibition of proliferation concentration of approximately 0.20 mg/mL for bilberry extract, indicating that the extract's mechanism for reducing cell proliferation may be by inducing early apoptosis. The Apo ssDNA assay (Fig. 3) revealed an apoptotic cell population of 18% for control (untreated) cells. For camptothecin (a positive control), the apoptotic cell percentage was 55%. Bilberry extract proved to be a potent inducer of apoptosis with 49% induction of apoptosis at 0.125 mg/mL (Fig. 3). We also used a TUNEL assay and obtained results similar to those with the Apo ssDNA assay (data not shown). Our TUNEL and Apo ssDNA data agree with the results of Katsube et al. 8 and Wu et al. 20 using a different assay, that bilberry extract induces apoptosis in MCF7 cells.
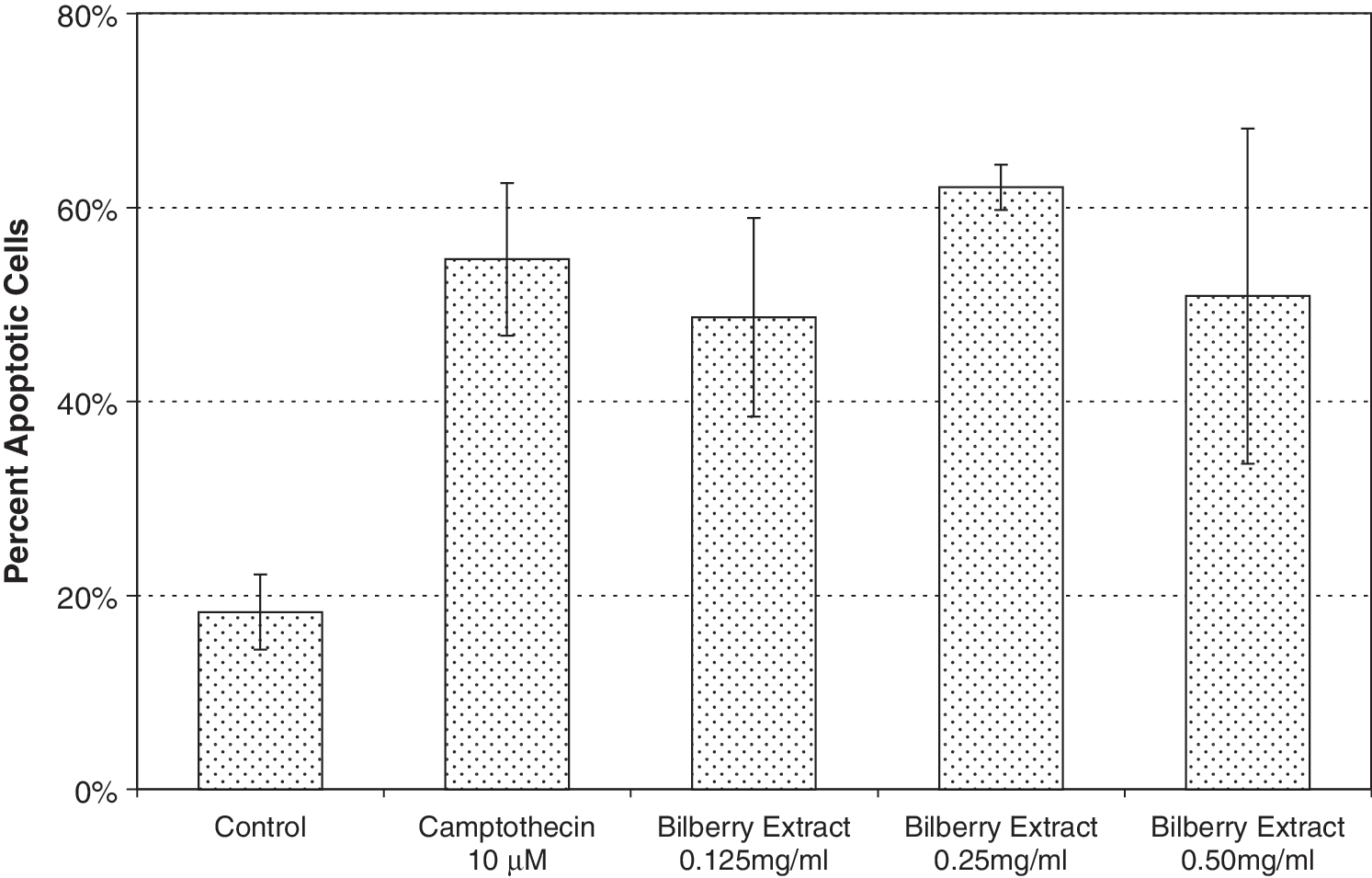
Apo ssDNA Assay. MCF7 cells were incubated for 6 hours with camptothecin and for 24 hours with various concentrations of bilberry extract. The amount of apoptotic cells was determined by flow cytometry after fixation and monoclonal antibody staining. A similar increase in apoptotic cells occurred with 0.25 and 0.5 mg/mL bilberry extract. Camptothecin served as a positive control.
Our data clearly show that bilberry extract inhibits cell proliferation in association with induction of apoptosis. However, cell cycle analysis indicates that at the IC50 the extract does not block at a specific cell cycle stage. However, higher concentrations of extract did increase the fraction of cells at G2/M. Such an action could involve modulating microtubule organization or polymerization. To explore this possibility further, we used immunofluorescence microscopy to examine the cellular effects of bilberry extract on microtubules, actin filaments, and DNA. We obtained immunofluorescence images of microtubules (green), actin (red), and DNA (blue) in fixed MCF7-GFP-tubulin mitotic or interphase cells after incubation in the absence or presence of bilberry extract for 24 hours.
Shown in Figure 4 are cell images taken at a number of extract concentrations. At increasing concentrations, larger numbers of cells were visibly rounded along their perimeters, and the cytoskeletal elements were depolymerized. At 0.125 mg/mL extract (Fig. 4D–F), there was little cell damage, with a few cells appearing rounder than normal, and most looking like control cells (Fig. 4A–C) with microtubules through the cell and distinct actin filaments along the cell periphery. With 0.25 mg/mL extract (Fig. 4G–I), substantial numbers of cells appeared to be dead and dying, with a breakdown of microtubules and actin filaments. The tubulin appeared to be bundled into punctate spots at this concentration and higher. Only a small subset of cells retained their normal morphology at 0.25 mg/mL extract. With 0.5 mg/mL extract (Fig. 4J–L), all cells appeared to round up, with no microtubules or actin filaments present, and most of the tubulin bundled into punctate spots. The control was normal, with cell proliferation widely evident, but there were no cells in mitosis in 0.25–0.5 mg/mL extract-treated preparations that remained adhered to the coverslip. At each concentration, the images are typical of the majority of cells that remained on the coverslip.
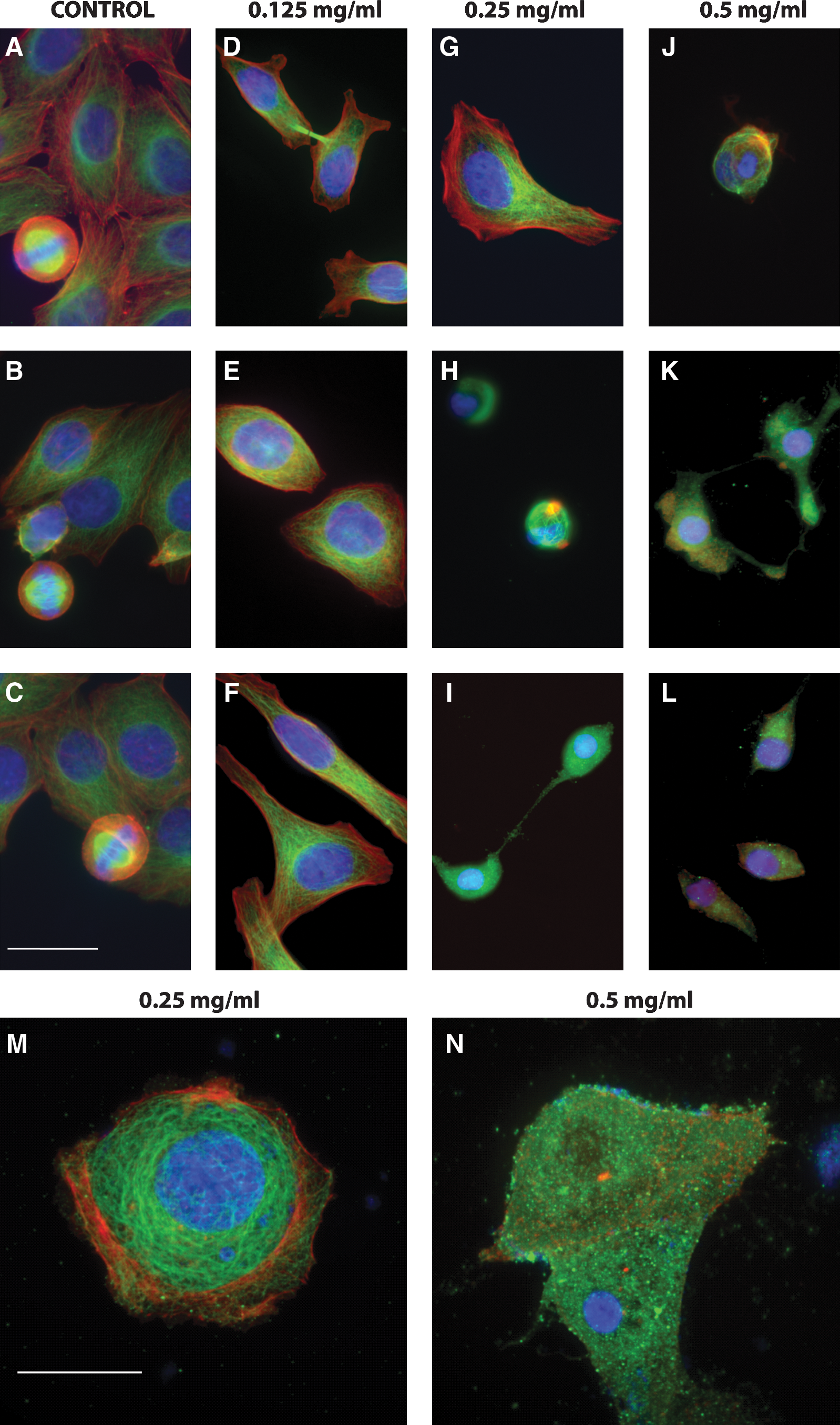
Effect of bilberry extract on microtubule organization in MCF7-GFP-tubulin cells shown in immunofluorescence images of microtubules (green), actin (red), and DNA (blue) in fixed MCF7-GFP-tubulin mitotic or interphase cells after incubation in the absence or presence of bilberry extract for 24 hours. Dividing cells were commonly seen (
Given that bilberry extract has a clear effect on the cytoskeleton and induces cell rounding, it is likely that the lack of mitotic cells on the coverslips is due to detachment of the rounded mitotic cells during processing.
Figure 4 contrasts our immunocytochemical findings at two key concentrations of extract. At a concentration of 0.25 mg/mL extract (Fig. 4G–I, enlarged in Fig. 4M), the microtubules and actin filaments were depolymerized in over half the cells. Tubulin was present in punctate spots peppered throughout the cytoplasm. The tubulin appeared to be aggregating in these spots. There was no evidence of cytokinesis. Actin filaments were also depolymerized in 0.25 mg/mL extract, with no filaments apparent and any visible actin aggregated throughout the cytoplasm. In contrast, at 0.125 mg/mL extract (Fig. 4D–F), microtubules were typically evident, and actin was primarily in a filamentous form. At this lower concentration, a minority of cells appeared more like those seen at 0.25 mg/mL extract, indicating that the extract affected a smaller fraction of the cells at this concentration.
To evaluate whether bilberry extract directly damages microtubules, we measured the effect of bilberry extract on tubulin polymerization in vitro. 27 Bilberry extract had only a modest effect on microtubule polymerization at low concentrations (Fig. 5). Only 5–10% inhibition of polymerization occurred with extract concentrations of 0.2 mg/mL or lower. However, inhibition was more pronounced at 0.5 mg/mL extract, increasing to approximately 30%. These data are consistent with the immunofluorescence microscopy data shown in Figure 4, where significant effects on the microtubules in cells occurred at an extract concentration of 0.5 mg/mL. Consistent with the effects of bilberry extract on microtubule polymerization in vitro, accumulation of cells at G2/M occurred only at a concentration of 0.6 mg/mL and above.
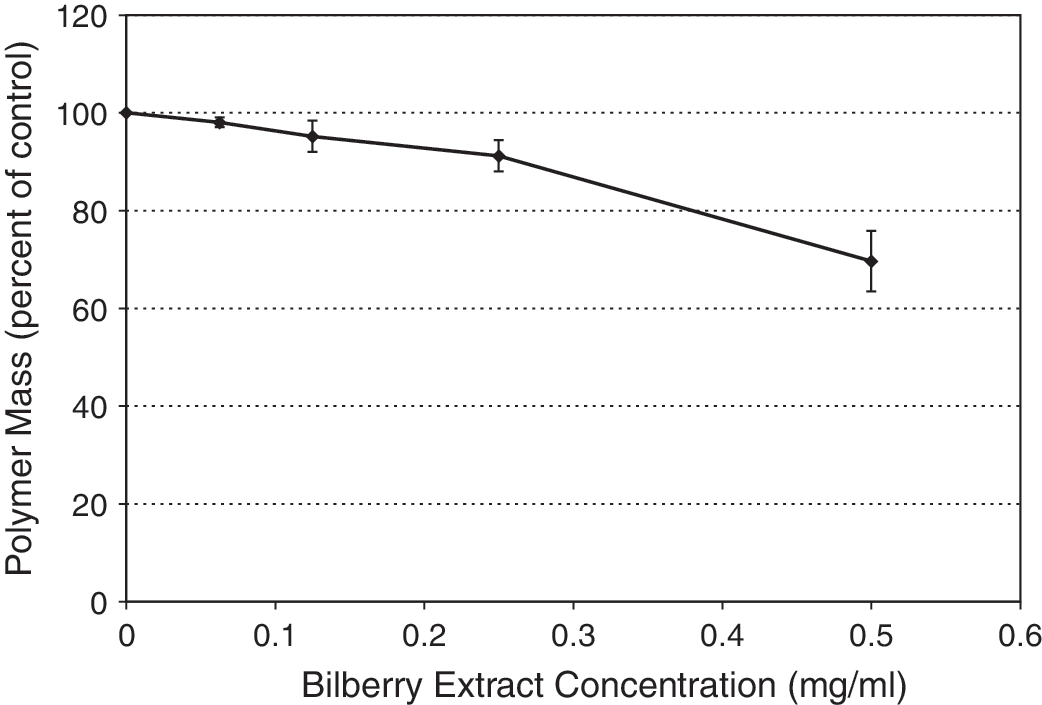
Polymerization of microtubules in the presence of bilberry extract. Microtubules were polymerized from a mixture of about 70% tubulin and 30% microtubule-associated proteins obtained from bovine brain at 30°C for 75 minutes in the presence of 2 mM GTP. 27 The concentration of tubulin was 17 μM. The amount of protein in the microtubule pellets was determined after sedimenting and resuspending the microtubules in ice-cold buffer. Results represent averages and SE of the data from three separate experiments.
Thus, we observed two distinct effects of bilberry extract in MCF7 cells: one that is likely due to microtubule damage, and the other that is not. At extract concentrations of 0.3–0.4 mg/mL, we observed induction of apoptosis and inhibition of proliferation. This effect occurred with extract concentrations well below that needed to affect tubulin polymerization and thus does not likely involve microtubule polymerization, but must occur through another mechanism. This mechanism remains to be determined. There was no accumulation of cells at G2/M at these concentrations, indicating that the bilberry extract was not inhibiting proliferation or inducing apoptosis by modulating the dynamics or organization of microtubules. 21
At higher extract concentrations (0.5 mg/mL and greater), the accumulation of cells at G2/M stage, the cell rounding, and the accumulation of tubulin in punctate spots correlate with a separate direct action of bilberry extract that only occurs at higher concentrations of the extract.
Footnotes
Acknowledgments
We thank the Santa Barbara Cottage Hospital Research Grant Program and Mr. Roy Mankovitz for providing the funding for this study. The technical assistance of Herb Miller, Sarah Giguere, and Katie Shanley is gratefully acknowledged. The use of cyanins in sunscreen applications is covered by U.S. Patent 6,783,754. The contact for licensing opportunities is: Roy Mankovitz, Director, Montecito Wellness LLC, 1482 East Valley Rd, Suite 808, Santa Barbara, CA 93108, USA, 1-805-969-4604,
Author Disclosure Statement
No competing financial interests exist.