
Abstract
The effects of a dry extract of the roots of Angelica sinensis (Oliv.) Diels (ASE) on the growth and differentiation of granulocyte-macrophage progenitor cells (CFU-GM) in normal and Listeria monocytogenes-infected mice were studied. Myelosuppression concomitant with increased numbers of spleen CFU-GM was observed in infected mice. Prophylactic administration of ASE (10, 25, and 50 mg/kg) stimulated marrow myelopoiesis in a dose-dependent manner and reduced spleen colony formation to control values. The dose of 50 mg/kg ASE was the optimal biologically active dose in infected mice, and this dose schedule significantly increased survival of mice infected with a lethal dose of L. monocytogenes, with survival rate up to 30%. Investigation of the production of colony-stimulating factors revealed a dose-dependent increased colony-stimulating activity in the serum of infected mice, with higher response produced by the 50 mg/kg dose. Notably, no effects were observed with the 100 mg/kg dose, compared with infected nontreated controls. Further studies to investigate the production of factors such as inteferon-γ and tumor necrosis factor-α demonstrated increased levels of both cytokines in mice infected with L. monocytogenes and treated with 50 mg/kg ASE. We propose that ASE indirectly modulates immune activity and probably disengages Listeria-induced suppression of these responses by inducing a higher reserve of myeloid progenitors in the bone marrow in consequence of biologically active cytokine release (colony-stimulating factors, interferon-γ, and tumor necrosis factor-α).
Introduction
A
L. monocytogenes has been extensively used as an experimental model to study the mechanisms involved in innate and cell-mediated antimicrobial defense. This Gram-positive, facultative intracellular bacterium replicates primarily within macrophages and triggers a striking increase in innate immune responses that require interactions among neutrophils, macrophages, and natural killer and T cells. As a foodborne human pathogen, it can cause significant disease in newborns, immunocompromised individuals, and pregnant women. 8 –11
During the early stages of infection, the production and migration of granulocytes and macrophages from bone marrow into infected tissues play a major role in limiting bacterial multiplication. 12 –14 These phagocytes are derived from hematopoietic progenitors in the bone marrow, and the production and mobilization of these cells depend on the presence of specific growth factors called colony-stimulating factors (CSFs). 15 –17 Several studies have shown that primary L. monocytogenes infection of mice is associated with a sharp rise in serum CSF levels, an increase in differentiated monocytes and granulocytes at the site of infection, and a decrease in the number of progenitor cells in the bone marrow. 12 –14,18 In addition, there is some evidence that higher numbers of progenitor cells in the bone marrow before infection favors survival of mice after L. monocytogenes infection. 11,19 –23
Using the listeriosis model we determined the effects of oral administration of a dry extract of A. sinensis extract (ASE) on the growth and differentiation of bone marrow and spleen granulocyte-macrophage progenitor cells (CFU-GM), serum CSF levels, and tumor necrosis factor-α (TNF-α) and IFN-γ production in mice. Our results demonstrated that ASE has a stimulating effect on the hematopoietic response and cytokine production of infected mice, which may be responsible, at least in part, for the enhanced protection during a lethal infection. No detectable changes on the parameters studied were observed in the normal mice.
Materials and Methods
Animals
Male BALB/c mice, 6–8 weeks old, were bred at the University Central Animal Facilities (State University of Campinas, Campinas, SP, Brazil), raised under specific pathogen-free conditions, and matched for body weight before use. Animal experiments were done in accordance with institutional protocols and the guidelines of the Institutional Animal Care and Use Committee. 24
L. monocytogenes infection
L. monocytogenes obtained from the Laboratory of Microbiology (Department of Clinical Pathology, Clinical Hospital, State University of Campinas) was used to infect the animals. Bacterial virulence was maintained by serial passages in BALB/c mice. Fresh isolates were obtained from infected spleens, grown in brain-heart infusion medium, and stored at −80°C in 1-mL aliquots. Before use, each sample was thawed and diluted to appropriated concentrations in 0.9% NaCl. Mice were inoculated intraperitoneally with a sublethal dose of 5 × 103 viable L. monocytogenes per mouse for the study of levels of CFU-GM, CSFs, TNF-α, and IFN-γ. To evaluate survival rate, mice were inoculated intraperitoneally with a lethal dose of 6 × 105 viable L. monocytogenes per mouse. In both cases, mice were infected at the end of a 10-day treatment with different concentrations of ASE.
ASE
The ASE was provided by Galena Chemical and Pharmaceuticals Ltd. (Campinas). It is a dry extract marketed by Jiangxi Medicines & Health Products, located in Nanchang, China. The plant and the extract were identified and evaluated according to the description of the characteristics and to its monograph present in the Chinese Pharmacopoeia. The dry extract was obtained using ethanol and water as solvents. The batch number of the raw material used in this study was 288469, and this lot was standardized and subjected to organoleptic, chemical, microbiological, and phytochemical analysis for the determination of its quality. The extract was standardized through its active agent ligustilide, with no fractions showing less than 1% of this active constituent. To quantify this active drug and to confirm the standardization, it was used an Agilent (Palo Alto, CA, USA) high-performance liquid chromatography column (Inertsil ODS-3, 4.6 × 150 mm; particle size, 5 μm) with a flow rate of 1.0 mL/minute; detection at 284 nm, methanol/water (65:35 vol/vol) as the mobile phase, a temperature of 25°C, and an injection volume of 10 μL. Methanol was used as the diluent in preparation of the solutions, and the external standard was prepared at a concentration of 0.04 mg/mL.
Treatment regimen
ASE was dissolved in sterile water and diluted into appropriate concentrations immediately before use. Doses of 10, 25, 50, and 100 mg/kg were administered orally to mice for 10 days prior to infection in a 0.2-mL volume per mouse. Mice were infected with a sublethal dose of bacteria 3 hours after the end of the treatment, and assays were performed 24, 48, and 72 hours after bacterial inoculation. Each experiment included parallel control groups of normal and infected mice treated with an equivalent volume of the vehicle. Selection of these doses was based on previous literature. 25 –28
Progenitor cell assay
Bone marrow cells were prepared aseptically from one complete femur shaft in RPMI 1640 medium (Sigma Chemical Co., St. Louis, MO, USA). Spleens were then aseptically removed and converted to dispersed cell suspensions in RPMI 1640 medium by gently pressing through a stainless steel mesh net. Bone marrow and spleen cells were placed in duplicate 1-mL semisolid agar cultures in 35-mm-diameter Petri dishes using 1 × 105 bone marrow cells or 2.5 × 105 spleen cells per culture. The medium used was Dulbecco's modified Eagle's medium (Sigma) containing 20% fetal calf serum (FCS) and 0.3% agar. Colony formation was stimulated by addition of 0.5 ng/mL recombinant murine granulocyte-macrophage CSF (Sigma). The cultures were incubated for 7 days in a fully humidified atmosphere of 5% CO2 in air, and colony formation (clones >50 cells) was scored at × 35 magnification using a dissection microscope. Morphological analysis was performed in the whole cultures fixed in 2.5% glutaraldehyde and stained with Luxol fast blue/hematoxylin as previously described. 29
Assay for serum colony-stimulating activity
Mice were bled from the heart under halothane anesthesia. Within each experimental group, blood was pooled in periods of 24, 48, and 72 hours after the L. monocytogenes challenge. Pooled blood was left at 37°C for 30 minutes, and the clots were allowed to retract overnight at 4°C. Following centrifugation, the serum was removed and stored at −20°C. Colony-stimulating activity (CSA) was determined by the ability of serum obtained from control and experimental groups to induce the growth and differentiation of bone marrow progenitor cells (1 × 105 cells) from normal mice. Results are expressed as units of CSF/mL, where 1 unit/mL was defined as the lower amount of CSF that induces the formation of colonies. 30
Levels of IFN-γ and TNF-α in spleen mononuclear cell culture
Spleens of mice subjected to the treatment of 50 mg/kg ASE and sacrificed after 48 hours of infection were collected aseptically, macerated, and homogenized in RPMI 1640 medium (Cultilab, Campinas). Mononuclear cells were obtained by adding lysis buffer to the cell button. After being washed, cells were resuspended in 10 mL of RPMI 1640 medium supplemented with 10% FCS and incubated in glass Petri dishes (100 × 20 mm) for 2 hours at 37°C in 5% CO2. After this period, nonadherent cells were discarded, the adherent cells were trypsinized and washed, and the concentration was adjusted to 1 × 105 cells/mL of RPMI 1640 medium enriched with 5% FCS with a hemacytometer. Splenocytes (1 × 105 cells/mL) were suspended in enriched RPMI 1640 culture medium supplemented with 5% FCS and seeded into 24-well culture plates in the presence of 50 μL of lipopolysaccharide solution (5 μg/mL). Simultaneously, each sample of cells was incubated without lipopolysaccharide for control. After 48 hours of incubation at 37°C in 5% CO2, each supernatant was individually collected and frozen at −20°C. IFN-γ and TNF-α were quantified by enzyme-linked immunosorbent assay, using a specific antibody for each substance (DuoSet® enzyme-linked immunosorbent assay development kits, mouse TNF-α/TNFSF1A [catalog number DY410] and mouse IFN-γ [catalog number DY485] [R&D Systems, Minneapolis, MN, USA]). Quantification was performed in microtiter plates (96-well flat-bottom Maxisorp, Nunc, Roskilde, Denmark), following the protocol of use of the manufacturer for each set. The readings were held on the enzyme-linked immunosorbent reader at 450 nm.
Survival analysis
The protective effects of ASE were evaluated by determining the survival time of BALB/c mice treated for 10 consecutive days with oral daily doses of 10, 25, 50, and 100 mg/kg ASE. Animals were infected with a lethal dose of L. monocytogenes 3 hours after the end of treatment and observed for 30 days.
Statistical analysis
For statistical analysis of changes in progenitor cell assays, CSA, and hematological parameters, a parametric method, the one-way analysis of variance followed by Tukey's test, was used to compare data among all groups. Analysis of cytokine levels was done by Kruskal-Wallis nonparametric analysis of variance with Dunn's Multiple Comparisons test to compare single groups. The probability of survival among the experimental groups was calculated by the Kaplan-Meier method, and the logarithm-rank statistic was used to test differences between groups. Statistical significance was considered when P < .05.
Results
Survival analysis
The antibacterial efficacy of the prophylactic administration of ASE was evaluated in mice infected with a lethal dose of L. monocytogenes (Fig. 1). All the untreated infected mice died within 5 days after infection. The prophylatic treatment of these animals with the doses of 10, 25, and 50 mg/kg ASE for 10 days significantly increased survival (P < .001). When the dose of 10 and 25 mg/kg was used, 10% and 20%, respectively, of these animals survived, whereas with the doses of 50 mg/kg, 30% survival was observed. In contrast, the dose of 100 mg/kg did not present any protection against lethality.

Survival of lethally infected mice pretreated with different doses of A. sinensis extract for 10 consecutive days. At the end of A. sinensis extract administration, mice were inoculated intraperitoneally with 6 × 105 Listeria per animal. Groups of 20 mice were checked daily for survival. P < .001 in relation to infected mice.
Medullar and splenic hematopoiesis
The effects of the pretreatment with 10, 25, 50, and 100 mg/kg ASE on bone marrow hematopoietic activity of normal and L. monocytogenes-infected mice are presented in Figure 2. Consistent with our previous results, 20 –23 the number of bone marrow CFU-GM was significantly reduced in infected mice at 48 and 72 hours after bacterial challenge (P < .001). The lower response was observed at 48 hours, when the number of CFU-GM reached less than 50% of control values. On the other hand, at 24 hours after the infection, no significant differences were found between the hematopoietic response of control and infected mice. Significant dose-dependent protection against the CFU-GM suppression induced by the infection was observed in the bone marrow of mice pretreated with ASE. With the 10 and 25 mg/kg doses, CFU-GM numbers reached control levels, whereas stimulation over control values was produced by the 50 mg/kg dose, which was present as soon as 24 hours after bacterial challenge (P < .001) and sustained throughout the experiment. Of note is that no effects were produced by the 100 mg/kg dose on marrow CFU-GM, compared to infected controls.
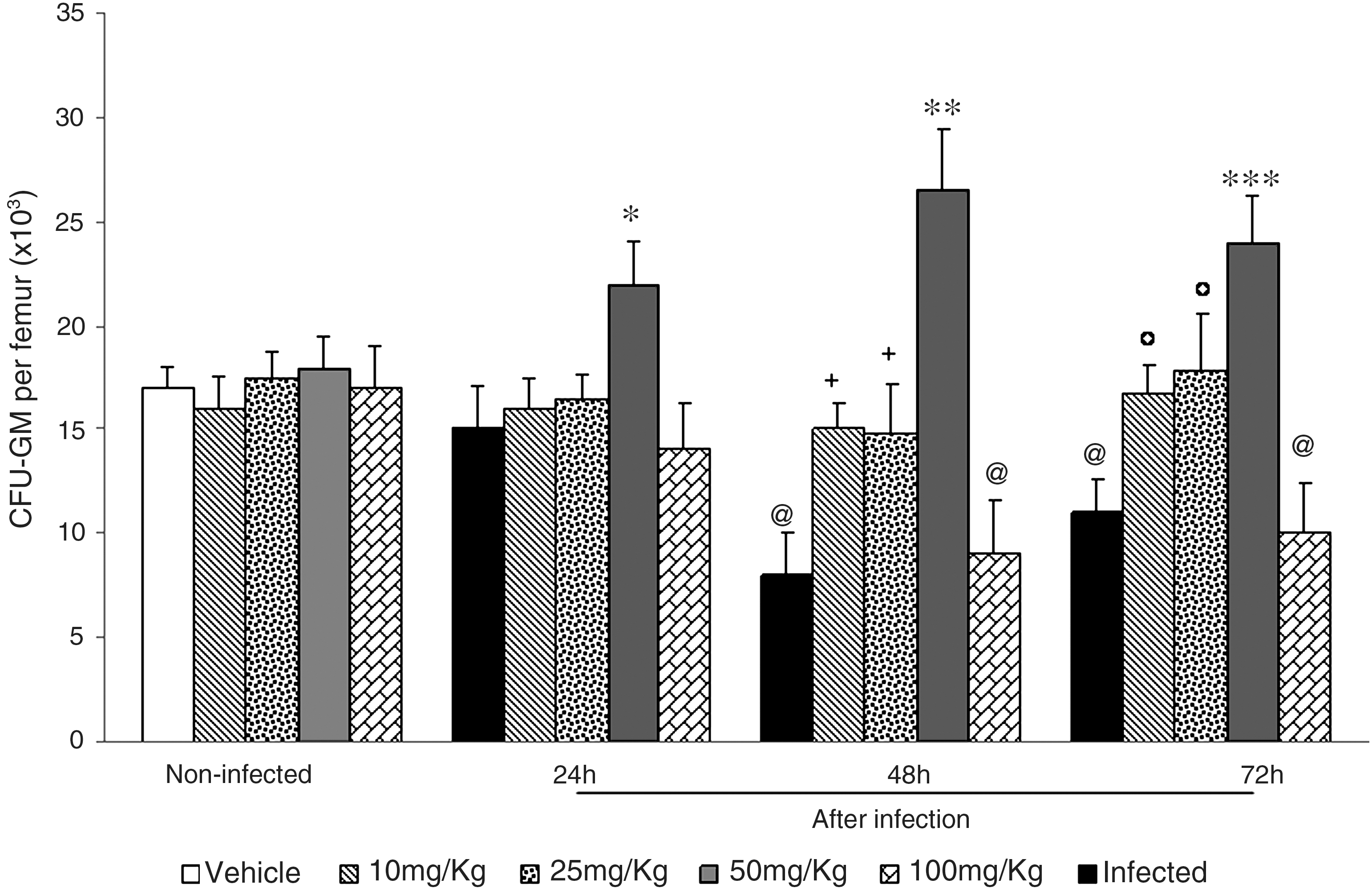
Number of bone marrow granulocyte-macrophage progenitor cells (CFU-GM) in normal and L. monocytogenes-infected mice pretreated with A. sinensis extract. A. sinensis extract at 10, 25, 50, and 100 mg/kg was given by gavage for 10 consecutive days. Three hours after the last dose, mice were infected intraperitoneally with a sublethal dose of 5 × 103 Listeria per mouse. CFU-GM numbers were determined at 24, 48, and 72 hours after infection. Experiments were done twice with six mice per group each time. Control mice received vehicle only. Results are mean ± SD values of 12 mice per group. By analysis of variance and Tukey's test: *P < .001 in relation to noninfected, infected 24 hours, and infected-treated; @ P < .001 in relation to noninfected; + P < .001 in relation to infected 48 hours; **P < .001 in relation to noninfected, infected 48 hours, and infected-treated; °P < .001 in relation to infected 72 hours; ***P < .001 in relation to noninfected, infected 72 hours, and infected-treated.
Because L. monocytogenes infection is generally accompanied by higher numbers of progenitor cells in the spleen, 20 –23 we next evaluated the compartment of committed myeloid progenitors in this organ (Fig. 3). The dramatic increase in spleen CFU-GM numbers observed at 48 and 72 hours after the infection (P < .001) was fully reversed by treatment with 10, 25, and 50 mg/kg ASE for 10 days prior to bacterial inoculation (P < .001). As before, the 100 mg/kg dose was ineffective in reducing CFU-GM numbers in infected mice. In normal, noninfected mice, ASE treatment produced no changes in CFU-GM numbers in bone marrow and spleen.
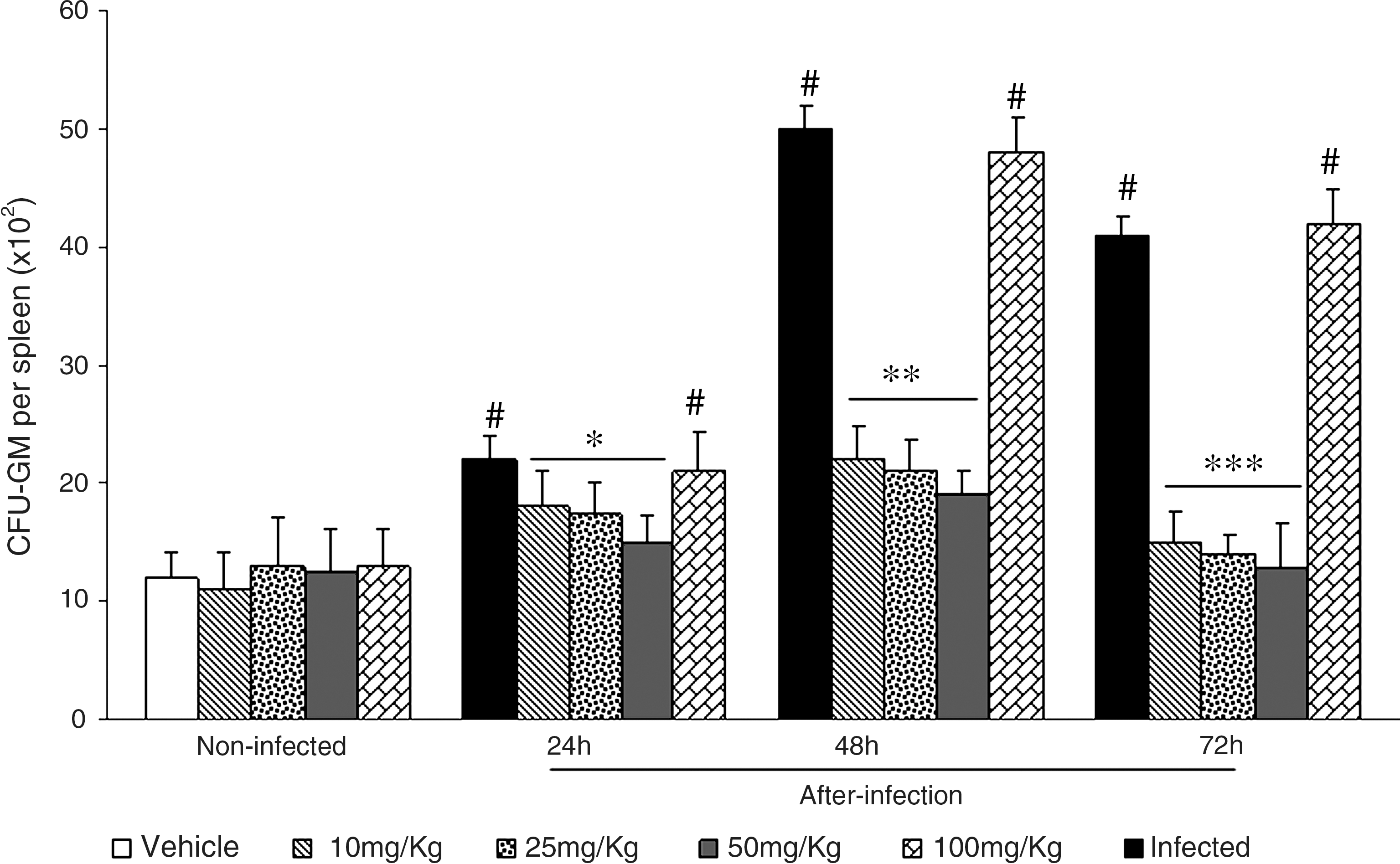
Number of spleen CFU-GM in normal and L. monocytogenes-infected mice pretreated with A. sinensis extract. A. sinensis extract at 10, 25, 50, and 100 mg/kg was given by gavage for 10 consecutive days. Three hours after the last dose, mice were infected intraperitoneally with a sublethal dose of 5 × 103 Listeria per mouse. CFU-GM numbers were determined at 24, 48, and 72 hours after infection. Experiments were done twice with six mice per group each time. Control mice received vehicle only. Data are mean ± SD values of 12 mice per group. By analysis of variance and Tukey's test: # P < .001 in relation to noninfected; *P < .001 in relation to infected 24 hours; **P < .001 in relation to infected 48 hours; ***P < .001 in relation to infected 72 hours.
Serum CSA
The effects of ASE in serum CSA are presented in Figure 4. The treatment with 10, 25, 50, and 100 mg/kg ASE produced no effects in CSA levels of normal, noninfected mice. The presence of the infection significantly increased CSA titers, with a peak at the interval of 48 hours after bacteria inoculation, reaching levels fivefold higher than noninfected controls. Interestingly, at 24 hours of infection, the 50 mg/kg dose was the only one to produce a significant increase in CSA titers, reaching titers 1.5-fold higher than in infected controls. At 48 and 72 hours after infection, however, in addition to the 50 mg/kg, the doses of 10 and 25 mg/kg also increased CSA titers, in a dose-dependent manner. Of importance is that the CSA response produced by the 50 mg/kg dose was, in all instances, significantly higher than the one produced by the two lower doses. It is interesting to mention that no further increase in CSA titers was produced by the higher dose of 100 mg/kg in relation to the infected group.

Colony-stimulating activity (CSA) in serum samples of normal and L. monocytogenes-infected mice pretreated with A. sinensis extract. A. sinensis extract at 10, 25, 50, and 100 mg/kg was given by gavage for 10 consecutive days. Three hours after the last dose, mice were infected intraperiteonally with a sublethal dose of 5 × 103 Listeria per mouse. CSA was determined at 24, 48, and 72 hours after infection. Experiments were done twice with six mice per group each time. Control mice received vehicle only. Data are mean ± SD values of 12 mice per group. By analysis of variance and Tukey's test: °P < .001 in relation to control noninfected; # P < .001 in relation to noninfected and infected 24 hours; + P < .001 in relation to noninfected and infected 48 hours; @ P < .001 in relation to noninfected, infected 48 hours, and infected-treated; *P < .001 in relation to noninfected and infected 72 hours; **P < .001 in relation to noninfected, infected 72 hours, and infected-treated.
Levels of IFN-γ and TNF-α in spleen mononuclear cell culture
The effects of the pretreatment with 50 mg/kg ASE on IFN-γ and TNF-α levels of normal and L. monocytogenes-infected mice are presented in Figures 5 and 6. Both cytokines were measured 48 hours after L. monocytogenes infection. The treatment of noninfected mice with ASE produced an increase in IFN-γ (P < .001) and TNF-α (P < .001) production. Similarly, the infection alone produced significant rises in the levels of both cytokines (P < .001), compared to noninfected controls. Treatment of infected mice with ASE further increased the level of both cytokines, reaching levels 1.2-fold higher for IFN-γ and 1.9-fold higher for TNF-α, compared to those of controls that were only infected. The rise in level of both cytokines was consistent with higher numbers of CFU-GM in the bone marrow (Fig. 2).
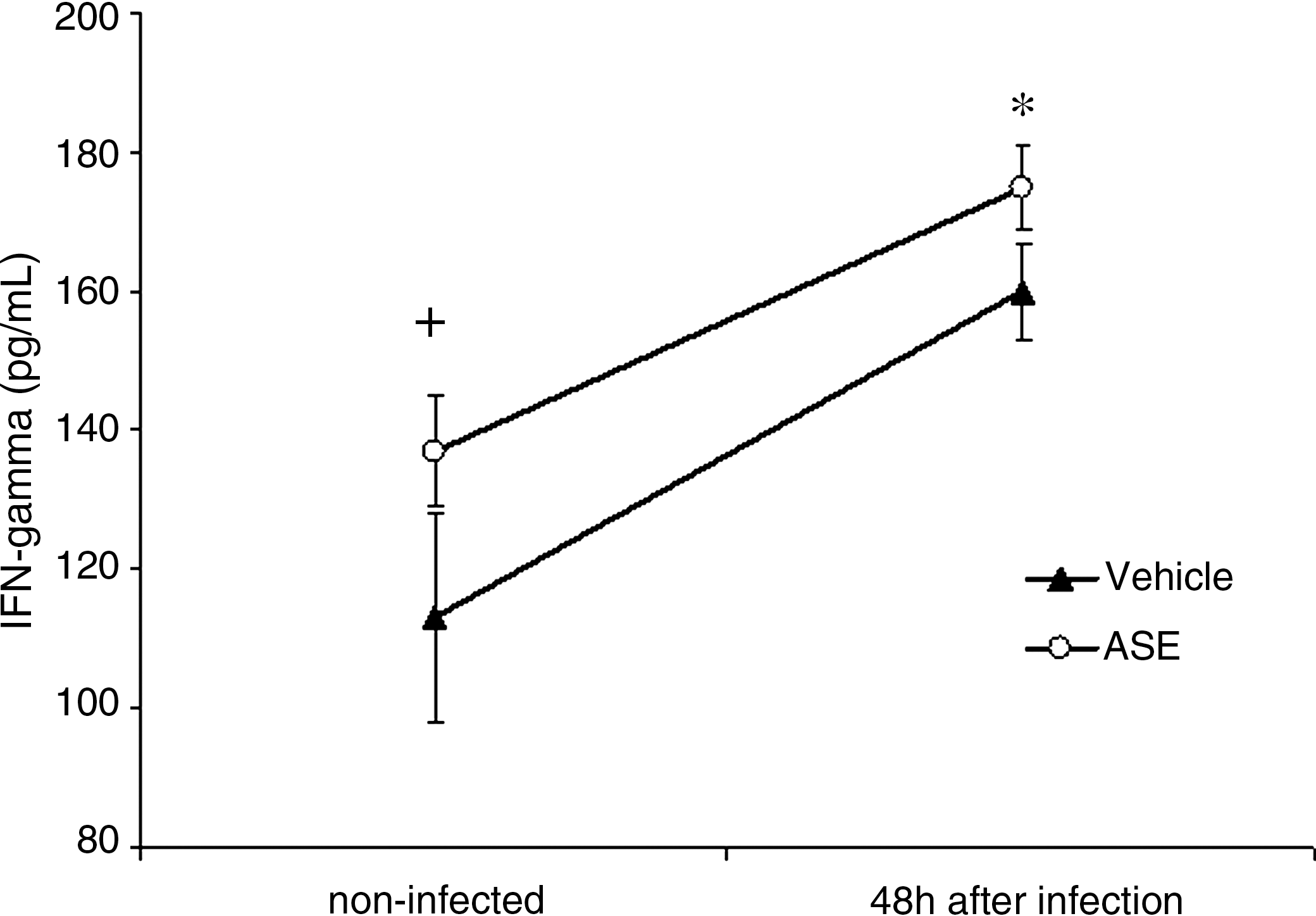
Interferon-γ (IFN-gamma) levels of normal and L. monocytogenes-infected mice pretreated with A. sinensis extract (ASE). The dose of 50 mg/kg was given by gavage for 10 consecutive days. Three hours after the last dose, mice were infected intraperitoneally with a sublethal dose of 5 × 103 Listeria per mouse. IFN-γ was determined at 48 hours after infection. Experiments were done twice with six mice per group each time. Control mice received vehicle only. Data are mean ± SD values of 12 mice per group. + P < .001 in relation to noninfected; *P < .001 in relation to noninfected and infected 48 hours.

Tumor necrosis factor-α (TNF-alpha) levels of normal and L. monocytogenes-infected mice pretreated with ASE. The dose of 50 mg/kg was given by gavage for 10 consecutive days. Three hours after the last dose, mice were infected intraperiteonally with a sublethal dose of 5 × 103 Listeria per mouse. TNF-α was determined at 48 hours after infection. Experiments were done twice with six mice per group each time. Control mice received vehicle only. Data are mean ± SD values of 12 mice per group. + P < .001 in relation to noninfected; *P < .001 in relation to noninfected and infected 48 hours.
Discussion
Studies from our laboratory have defined listeriosis as a useful experimental model to study the influence of various natural compounds and toxicants in the many components of the host immune responses involved in the defense against L. monocytogenes, with special emphasis on the ability of hematopoietic tissues to produce and mobilize phagocytes. 20 –23 It has been well demonstrated in the literature that during the early phase of Listeria infection there is a decrease in the number of CFU-GM in bone marrow related with a massive migration to the spleen, showing an extramedullar hematopoiesis with consequently splenomegaly and an increase in concentrations of CSFs. 11,19,20,23,24,29 Granulocytes and macrophages are vital to the defense against bacterial infection. 14,17 Several works have demonstrated that in listeriosis the ability of bone marrow to produce and mobilize granulocytes and macrophages to the site of infection is an essential component of the host defense during the early inflammatory response and consequently survival. 12 –23
Our results corroborate previous data showing that the reduced frequency of progenitor cells in the bone marrow of BALB/c mice infected with Listeria determines a deficient inflammatory response and an impaired natural resistance. 20 –23 The oral administration of ASE to infected mice at doses varying from 10 to 50 mg/kg, for 10 days prior to a sublethal dose of bacteria, prevented myelosuppression and concomitantly restored spleen CFU-GM numbers. Interestingly, the optimal biologically active dose of ASE (50 mg/kg) further increased CFU-GM production in the bone marrow of infected mice to levels above the control, as soon as 24 hours after bacterial inoculation, and this effect was sustained at least up to 3 days. Similarly, this dose schedule also increased CSA titers already at the stage of 24 hours of infection, in the same manner as for CFU-GM. The persistent elevation of CSF levels in infected mice serves as a continuing stimulus that support the survival, proliferation, differentiation, and end cell function of granulocytes and monocytes, once these levels return to normal when bacterial counts in the liver and spleen approach zero. 13 –15,31 –37 This is in keeping with the observations that ASE stimulated CSA activity and marrow myelopoiesis were observed only in mice that were subsequently infected by Listeria and not in noninfected mice.
Although the precise mechanism whereby ASE improved the survival rate of mice infected with Listeria is not completely elucidated, our results clearly demonstrated that the increased reserve of bone marrow CFU-GM induced by this plant extract plays an important role in the resistance to infection. This is in agreement with literature data showing that higher numbers of myeloid progenitors in the bone marrow of genetically resistant mice are associated with earlier influx of phagocytes to the site of infection and increased resistance. C57BL/10 mice, which are genetically resistant to L. monocytogenes, present higher numbers of CFU-GM in the bone marrow both before and after the infection, compared to BALB/C mice. In addition, an earlier influx of phagocytes to the site of infection is observed in the resistant C57BL/10 strain, probably as a consequence of the higher numbers of precursor cells in the bone marrow observed in these animals. 14,18,38 –40 It seems, therefore, that the optimal response produced by the 50 mg/kg dose schedule might be related to a rapid mobilization of phagocytes to infected organs, thus accounting for a greater ability to recover from bacterial challenge. The significant increase (30%) of survivors detected in mice challenged with a lethal dose of Listeria after the treatment with 50 mg/kg ASE corroborates this assumption. By the same token, the lack of protection observed with the treatment with 100 mg/kg ASE might be related to the fact that with this dose schedule there was no increase in serum CSA and CFU-GM numbers throughout the study.
Another important observation was that the increase in myeloid progenitors in the bone marrow of L. monocytogenes-infected mice mediated by the pretreatment with the extract occurs concomitantly with a reduction in the splenic CFU-GM. It is known that alterations in the distribution of hematopoietic activity in the tissues may occur as a result of increased hematopoietic stress, like those provoked by experimental murine infections. 19,41,42 In addition, hematopoietic stem cells and progenitor cells can migrate from the bone marrow in blood to other tissues like the spleen as part of the host defenses against intracellular bacteria, sustaining a second level of compensatory hematopoietic response. 12,43,44 Therefore, the reduced hematopoietic activity found in the spleen of mice treated with the 10, 25, and 50 mg/kg doses, which was not observed with the 100 mg/kg treatment, presumably reflected a lower number of bacteria as a result of the improved protective mechanisms induced by the three lower dose schedules.
The lack of beneficial effects by ASE at 100 mg/kg might also be associated with the fact that substances that activate the mitogen-activated protein kinase pathways, as is the case of ASE, might elicit diverse pharmacological effects depending on the dose used. At low doses, they may lead to the expression of survival (c-Fos, c-Jun) and defensive genes (Phase II detoxifying enzymes), resulting in survival and protective mechanisms. With increased doses, these substances will additionally activate the caspase pathway, leading to apoptosis (potential cytotoxicity). Further increment to suprapharmacological concentrations will lead to nonspecific necrotic cell death, 45 thus offsetting the beneficial effects of the active ingredients. Similar findings were obtained by other studies investigating naturally occurring lactones, such as coumarin, a component present in ASE, which, depending on its concentration, can be converted to a toxic substance, 3,4-hydroxyphenylacetaldehyde. 46,47
Macrophages are able to interact with various cellular products of the innate and acquired immune response. Of these, the synergistic action of TNF-α and IFN-γ has a fundamental role in the control of infection. 48 –52 TNF-α is an important cytokine mainly produced by activated macrophages, which is able to increase the oxidative burst after phagocytosis. Macrophage stimulation with IFN-γ induces direct antimicrobial and antitumor mechanisms and up-regulates antigen processing and presentation pathways. IFN-γ orchestrates leukocyte attraction and directs growth, maturation, and differentiation of many cell types, in addition to enhancing natural killer cell activity and regulating B cell functions. The infection alone produced a rise in the levels of both cytokines. The treatment of mice with 50 mg/kg ASE prior to bacterial inoculation further increased the levels of both cytokines. These findings show that a possible interaction of TNF-α, IFN-γ, and CSFs in mice treated with ASE contributes as an additional stimulus to the production, migration, and functional activity of neutrophils and macrophages during infection because higher numbers of CFU-GM were concomitantly found in the bone marrow. Of interest is the observation that ASE up-regulated TNF-α and IFN-γ production in noninfected mice treated with 50 mg/kg ASE, which seems to corroborate previous findings in the literature showing a direct activation of the T helper 1 population of lymphocytes by this plant extract. 7
All together, the results presented herein suggest that ASE indirectly modulates immune activity and probably disengages Listeria-induced suppression of these responses by up-regulating the production of CSFs and inducing a higher reserve of myeloid progenitors in the bone marrow, favoring an efficient and sustained response of phagocytes in the control of infection, as a consequence of release of biologically active cytokine (CSFs, TNF-α, and IFN-γ). Taking into account the limitations of dose schedule, it is encouraging in this context to consider ASE for combination therapy to protect the host from hematotoxicity as well as to supplement bactericidal efficacy.
Footnotes
Acknowledgment
Financial support was provided by the Conselho Nacional de Desenvolvimento e Tecnológico.
Author Disclosure Statement
No competing financial interests exist.