
Abstract
In this study, the protective role of grape seed extract (GSE) against doxorubicin (DOX)-induced cardiotoxicity and genotoxicity has been evaluated in male Mus musculus var. albino mice. The micronucleus (MN) test in erythrocytes and the chromosome aberration (CA) test in bone marrow cells were used. Also, levels of reduced glutathione (GSH) and lipid peroxidation as malondialdehyde (MDA) in heart homogenates were measured, and in addition the changes in heart histology were investigated. The mice were randomly divided into six groups. Group I (negative control) received intraperitoneal injections of isotonic saline (0.02 mL/g) for 6 consecutive days, Group II received intraperitoneal injections of DOX (2.5 mg/kg of body weight, six doses every other day; cumulative dosage, 15 mg/kg of body weight) for 6 consecutive days, Group III received GSE (50 mg/kg of body weight, 21 doses every other day; cumulative dosage, 1,050 mg/kg of body weight) for 21 consecutive days, Group IV received GSE (150 mg/kg of body weight, 21 doses every other day; cumulative dosage, 3,150 mg/kg of body weight) for 21 consecutive days, Group V received GSE (50 mg/kg of body weight, 28 doses every other day; cumulative dosage, 1,400 mg/kg of body weight) for 28 consecutive days plus DOX (2.5 mg/kg of body weight, six doses every other day; cumulative dosage, 15 mg/kg of body weight) for 6 consecutive days, and Group VI received GSE (150 mg/kg of body weight, 28 doses every other day; cumulative dosage, 4,200 mg/kg of body weight) for 28 consecutive days plus DOX (2.5 mg/kg of body weight, six doses every other day; cumulative dosage, 15 mg/kg of body weight) for 6 consecutive days. DOX induced heart damage as indicated from a pronounced change in heart histology. In the DOX-treated group, there was a significant increase in MDA content in the heart homogenate, and the level of GSH was significantly decreased. DOX induced genotoxicity by increasing the number of aberrant metaphases (AMNs), MNs, and structural chromosomal aberrations (CAs) such as chromatid breaks, dicentrics, acentric fragments, and gaps and showed a detractive effect on the mitotic index (MI) of cells. Pretreatment with GSE before treatment with DOX significantly protected the heart tissue by ameliorating its antioxidant activity. In Groups V and VI, the MDA level of heart tissue was significantly decreased, and the GSH level was increased compared to the DOX-treated group. Moreover, GSE significantly protected bone marrow chromosomes from DOX-induced genotoxicity by reducing the total AMNs and the frequency of structural CAs. GSE treatment also decreased the frequency of MNs and increased the MI values. It could be concluded that GSE acts as a potent antioxidant to prevent heart damage and genotoxicity of bone marrow cells.
Introduction
D
The use of certain antioxidants such as selenium, N-acetylcysteine, and vitamin C may help to decrease of the cardiotoxicity and genotoxicity created by DOX and may inhibit mutagenesis and carcinogenesis. 8,9 Recently, biopolymer materials such as green tea, Ginkgo biloba, lycopene, and grape seed have been used for specially decreasing the toxic effects of DOX. 10 –13
Grapes and grape products are good sources of dietary flavonoids, which are powerful antioxidant compounds. 14 Grape seed's antioxidative activities are much stronger than those of vitamin C and vitamin E and may involve radical scavenging, quenching, and enzyme-inhibiting actions. Grape seed extract (GSE) is a complex mixture of polyphenols containing dimers, trimers, and other oligomers (procyanidins) of catechin and epicatechin and their gallate derivatives together known as the proanthocyanidins. 15 Beyond their antioxidant powers, proanthocyanidins may protect against atherosclerosis, gastric ulcer, large bowel cancer, cataracts, and diabetes. Thus, grape seed and GSE may benefit people with heart disease and cancer. 16 GSE also is marketed as a dietary supplement in countries such as the United States and Turkey, owing to several health benefits mainly attributed to its antioxidant property. Several experimental studies have demonstrated that GSE is highly bioavailable and provides significantly greater protection against free radicals and free radical-induced lipid peroxidation and DNA damage than vitamins C and E and β-carotene. GSE was also shown to demonstrate cytotoxicity towards human breast, lung, and gastric adenocarcinoma cells, while enhancing the growth and viability of normal human gastric mucosal cells. 17
The aim of the present study was to evaluate the protective role of GSE in DOX-induced genotoxicity and cardiotoxicity in Swiss albino mice.
Materials and Methods
Animals
The experiments were carried out on 36 male Mus musculus var. albino mice (12–14 weeks old, 25–30 g in body weight). Healthy mice were obtained from the Animal Research Center of the Refik Saydam Hifzissiha Institute (Ankara, Turkey). The mice were housed in stainless steel cages (26 × 15 × 50 cm in size) and kept under controlled conditions at 22 ± 3°C, 55 ± 5% relative humidity, and a 12-hour light/dark cycle throughout the experiment. The mice were acclimatized 1 week prior to the planned experiments and fed with standard pellet diet (Samsun Food Industry, Samsun, Turkey) and water ad libitum. In this study, the methods and techniques applied to mice were carried out according to the guidelines set by the World Health Organization (Geneva, Switzerland) and the ethical standards of the local ethical committee for animal experiments at Giresun University.
Products and chemicals
Grape seed (50 mg) (formerly Grape Seed PCO Phytosome 50–120 tabs) was purchased from Health Genesis Corp. (Bay Harbor Island, FL, USA). DOX powder for injection (Doxo-Teva, 10 mg per ampule) was purchased from Med-Ilaç San. Tic. A.S. (Istanbul, Turkey). Colcemid solution (10 μg/mL, catalog number 12-004-1) was purchased from Biological Industries Ltd. (Kibbutz Beit Haemek, Israel). Fast Green FCF (Sigma) and Grünwald Giemsa (Sigma) stains were purchased from Interlab A.S. (Istanbul). Saline (0.9% NaCl) (Baxter, Mediflex, Eczacibasi, Istanbul) was purchased from local pharmacies.
Experimental protocol
Albino mice were randomly divided into six groups each, consisting of six animals. Group I (negative control) received intraperitoneal injections of 0.9% NaCl (0.02 mL/g) for 6 consecutive days. Group II received intraperitoneal injections of DOX (2.5 mg/kg of body weight, six doses every other day; cumulative dosage, 15 mg/kg of body weight) for 6 consecutive days. Group III received orally GSE (50 mg/kg of body weight, 21 doses every other day; cumulative dosage, 1,050 mg/kg of body weight) for 21 consecutive days. Group IV received orally GSE (150 mg/kg of body weight, 21 doses every other day; cumulative dosage, 3,150 mg/kg body weight) for 21 consecutive days. Group V received orally GSE (50 mg/kg of body weight, 21 doses every other day; cumulative dosage, 1,400 mg/kg of body weight) for 28 consecutive days + DOX (2.5 mg/kg of body weight, six doses every other day; cumulative dosage, 15 mg/kg of body weight) for 6 consecutive days. Group VI received orally GSE (150 mg/kg of body weight, 21 doses every other day; cumulative dosage, 4,200 mg/kg of body weight) for 28 consecutive days + DOX (2.5 mg/kg of body weight, six doses every other day; cumulative dosage, 15 mg/kg of body weight) for 6 consecutive days.
For Groups V and VI, GSE application was started 7 days before exposure to DOX, continued together with DOX during 6 consecutive days, and was continued alone for 15 days after the last drug administration. Peripheral blood samples were obtained once 24 hours after the final treatment for all groups.
The dose of DOX in this study has been determined as 2.5 mg/kg of body weight. This dose was chosen because it induced an increase in the frequency of toxicity that was essential to determine the protective role of GSE. 18,19 Also, GSE doses were used that were comparable to those daily consumption amounts recommended by practitioners of nutritional medicine to support optimal health, and 50 and 150 mg/kg of body weight were effective doses for protection by GSE. 20
Mouse erythrocyte micronucleus assay for erythrocytes
The mouse erythrocyte micronucleus (MN) assay, a modified mouse MN test that conventionally scores the MN frequencies in bone marrow polychromatic erythrocytes, was used throughout this study. In this assay, the MN frequency is scored in mature normachromatic erythrocytes in the circulating blood obtained from the tail of the mouse. The mouse erythrocyte MN assay was performed according to the protocol of Te-Hsiu et al. 21 In brief, the mice were anesthetized with ether, and the blood samples were collected from a small puncture on the tail of the mouse. About 5 μL of peripheral blood collected from each mouse was immediately mixed with 3% EDTA solution in a cell well and smeared onto a clean slide. The erythrocytes were fixed in 70% methanol for 2 minutes and allowed to dry in a high temperature (40°C) incubator overnight. Then, slides were stained with 5% May-Grünwald Giemsa stain (15 minutes). Usually three or four slides were made and repeatedly double scored by two different observers. From prepared slides, a total of 1,000 normachromatic erythrocytes were scored for the presence of MNs under oil immersion at ×100 magnification using a binocular light microscope (model BX51, Olympus, Tokyo, Japan), and MN cells were photographed 22 at a magnification of × 500.
Preparation of bone marrow cells for chromosome analysis
Mice were injected intraperitoneally with 0.025% colcemid before killing and sacrificed 2 hours later under ether anesthesia. Femurs were dissected out, and bone marrow cells were aspirated, washed in physiological saline, treated hypotonically (0.075 M KCl), fixed in Carnoy's fixative, and stained with 5% Grünwald-Giemsa stain. 23 CAs were scored using oil immersion (with a ×100 object lens) under a light microscope (model BX51, Olympus). CAs were classified according to the classification of Savage. 24
Analysis of cells
Mitotic index
The mitotic index (MI) was determined as the percentage of dividing cells among 1,000 nucleated cells in slides prepared for each group.
Abnormal metaphase number
The number of aberrant metaphases (AMNs) was counted as the number of damaged metaphases among 100 metaphases in slides prepared for each group.
CA analysis
For all treatment groups 100 metaphases were counted for CAs such as chromatid breaks, chromatid gaps, fragments, and dicentrics, and they were considered to be equal.
Lipid peroxidation and glutathione
At the end of the treatment period, animals were sacrificed after an overnight fasting by exsanguination under ether anesthesia. The heart tissue of each animal was removed, cleaned, dried, and processed for biochemical measurements. The tissues were then homogenized in ice-cold 0.15 M KCl by a homogenizer (Ultraturrax type T25-B, IKA Labortechnik, Staufen, Germany) at 16,000 rpm for 3 minutes. The homogenates were centrifuged at 5,000 g at 4°C for 1 hour. The supernatants were stored at −40°C until they were analyzed. Tissue reduced glutathione (GSH) and malondialdehyde (MDA) contents were colorimetrically measured by the methods of Beutler et al. 25 and Yoshoiko et al., 26 respectively, using an ultraviolet-visible spectrophotometer (UVmini-1240, Shimadzu, Kyoto, Japan).
Histopathological examinations
For light microscopic examination, fresh tissue samples including heart were fixed in 10% neutral buffered formalin solution for routine processing, embedded in paraffin wax, sectioned at 5 μm, and stained with hematoxylin and eosin. Histopathological changes were semiquantitatively assessed under a light microscope with an ocular with grids and 4 ×, 10 ×, and 40 × objectives. A total of 10 high-power fields were randomly chosen. Changes in the experimental histopathological parameters were graded as follows: (–) indicated no changes, and (+), (++), and (+++) indicated mild, moderate, and severe changes, respectively.
Statistical analysis
The statistical analysis was carried out using SPSS for Windows version 10.0 statistical software (SPSS, Inc., Chicago, IL, USA). Statistically significant differences between the groups were compared using one-way analysis of variance and Duncan's test. The data are given as mean ± SE values, and values of P < .05 are considered statistically significant.
Results
CAs
Table 1 presents the results of CAs analysis in mitotic chromosomes of bone marrow cells of albino male mice treated with DOX alone, GSE alone, and GSE + DOX. The mice treated with 50 and 150 mg/kg of body weight doses of GSE did not show any significant difference in the total number of CAs, AMNs, and MI compared to the control group (P > .05). According to the classification of Savage, 24 four structural CAs were determined in the treatment groups. We found a high frequency of CAs such as chromatid breaks, dicentrics, acentric fragments, and chromatid gaps. Chromatid breaks occurred more frequently than the other CAs. Treatment of animals with DOX resulted in the induction of CAs and AMNs. As expected, mice treated with DOX at a dose of 2.5 mg/kg of body weight presented a high frequency in both the total number of CAs and AMNs compared with the negative control (P < .05). There was also a considerable decrease in MI (Table 1), and Duncan's test showed that this decrease was statistically significant (P < .05).
Data are mean ± SE values (n = 6). One hundred cells were analyzed per animal (six animals per group, for a total of 600 cells per treatment) for CAs and AMNs. The MI has been calculated by analyzing 1,000 cells per animal (for a total of 6,000 cells per treatment), and percentage of the MI was calculated for each treatment group. Statistical significance between means was assessed using one-way analysis of variance followed by Duncan's test as a post-analysis of variance test.
Within a column, means not sharing the same letter are significantly different (P < .05).
AMNs, abnormal metaphases; CAs, chromosome aberrations; DOX, doxorubicin; GSE, grape seed extract; MI, mitotic index.
However, treatment of mice exposed to DOX with GSE resulted in a significant reduction in the frequency of CAs and AMNs. The number of CAs and AMNs showed a tendency to decrease in mice treated with 50 and 150 mg/kg of body weight doses of GSE compared with the DOX only-treated group. With the 150 mg/kg of body weight dose of GSE, this reduction for CAs such as chromatid breaks and dicentrics was by at least twofold and for AMNs was by at least threefold compared with the DOX group. In addition, the MI again presented a rising trend.
MN assay
The frequency of MNs is presented in Table 2. There was no statistically significant difference in the MN numbers of erythrocyte cells in the control and GSE-treated groups (P > .05). The mean number of MNs in erythrocytes was for the control group 3.80 ± 0.80, for the 50 mg/kg of body weight GSE group 4.00 ± 0.92, and for the 150 mg/kg of body weight GSE group 3.70 ± 0.68.
Data are mean ± SE values (n = 6). Statistical significance between means was performed using one-way analysis of variance followed by Duncan's test as a post-analysis of variance test.
Within a column, means not sharing the same letter are significantly different (P < .05).
The frequency of MNs clearly increased in the DOX-treated mice, and the mean number of micronucleated cells was significantly higher in the DOX only-treated group than in the negative control (P < .05). The treatment of animals with DOX associated with GSE (50 and 150 mg/kg of body weight) showed a significant reduction (P < .05) in the frequency of MNs compared with the group treated with DOX alone. The MN frequency decreased with the increase in the GSE doses. The MN frequency in the mice treated with the 150 mg/kg of body weight dose of GSE was lower than in mice treated with the 50 mg/kg of body weight dose of GSE. In brief, there was a significant dose–effect relationship between the MN frequency and GSE dose.
Lipid peroxidation and glutathione level
Table 3 shows the changes in GSH and MDA activities in the heart tissues of all groups after treatment with GSE (50 and 150 mg/kg of body weight) and DOX (2.5 mg/kg of body weight). Statistically significant differences between the control group and the group treated with the 50 mg/kg of body weight dose of GSE were not observed in GSH activity of the heart tissue (P > .05). But, this difference in GSH activity was statistically significant for the 150 mg/kg dose (P < .05). Moreover, statistically significantly differences between the control and both doses (50 and 150 mg/kg of body weight) of GSE were observed in MDA activity of heart tissue (P < .05). DOX application for 6 days altered the levels of GSH and MDA (P < .05). GSH levels significantly decreased, whereas MDA levels in the heart tissue of the DOX-treated group significantly increased compared with the control group (P < .05). However, oral administration of the two different doses of GSE (50 and 150 mg/kg of body weight) for 28 days reversed the GSH and MDA levels back to the control levels in the tissues, so that these differences were not statistically significant relative to the control group for the 150 mg/kg of body weight dose of GSE (P > .05).
Data are mean ± SE values (n = 6). Statistical significance between means was performed using one-way analysis of variance followed by Duncan's test as a post-analysis of variance test.
Within a column, means not sharing the same letter are significantly different (P < .05).
GSH, reduced glutathione; MDA, malondialdehyde.
Pathological findings
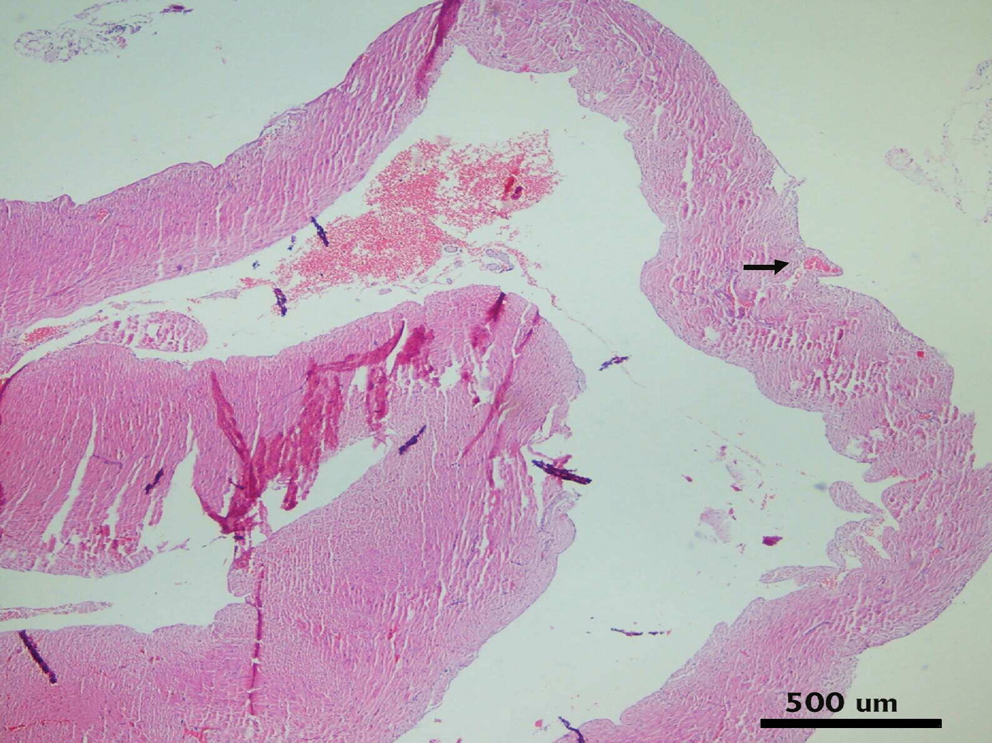
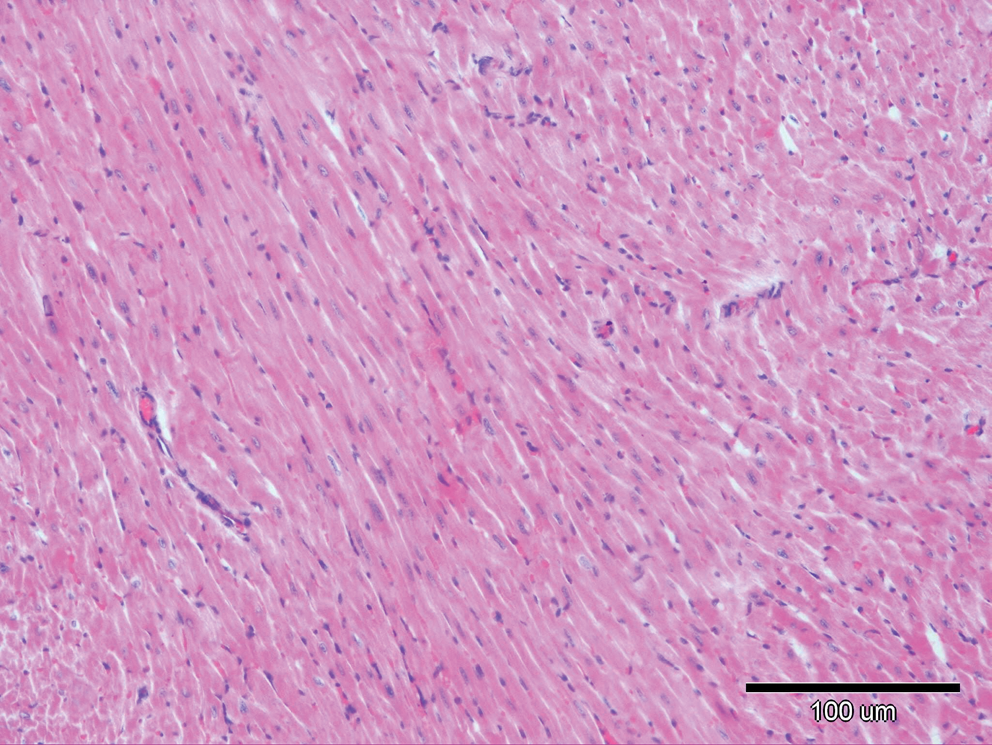
There were no histopathological lesions except limited hyperemic vessels in some myocardial tissue of Groups I, III, and IV (Figs. 1 –3). Severe histopathological changes were observed in group II. Many vessels were expanded and hyperemic (Fig. 4). Severe hemorrhagic foci (Fig. 4, inset) and degenerated myofibrils (Fig. 5) were seen. Similar lesions were detected in Groups V and VI. Hyperemic vessels were detected in many sections. However, a marked decrease in the severity of hemorrhage was observed in these groups (Figs. 6 and 7). Besides, degenerative myofibrils were noticed also in these groups (Figs. 6 and 7). Severities of histopathological lesions in the heart tissues were semiquantitatively displayed in Table 4.

Photomicrograph of Day 6 heart. In Group I (negative control), limited hyperemic vessels (arrows) are seen. Hematoxylin and eosin stain. Color images available online at
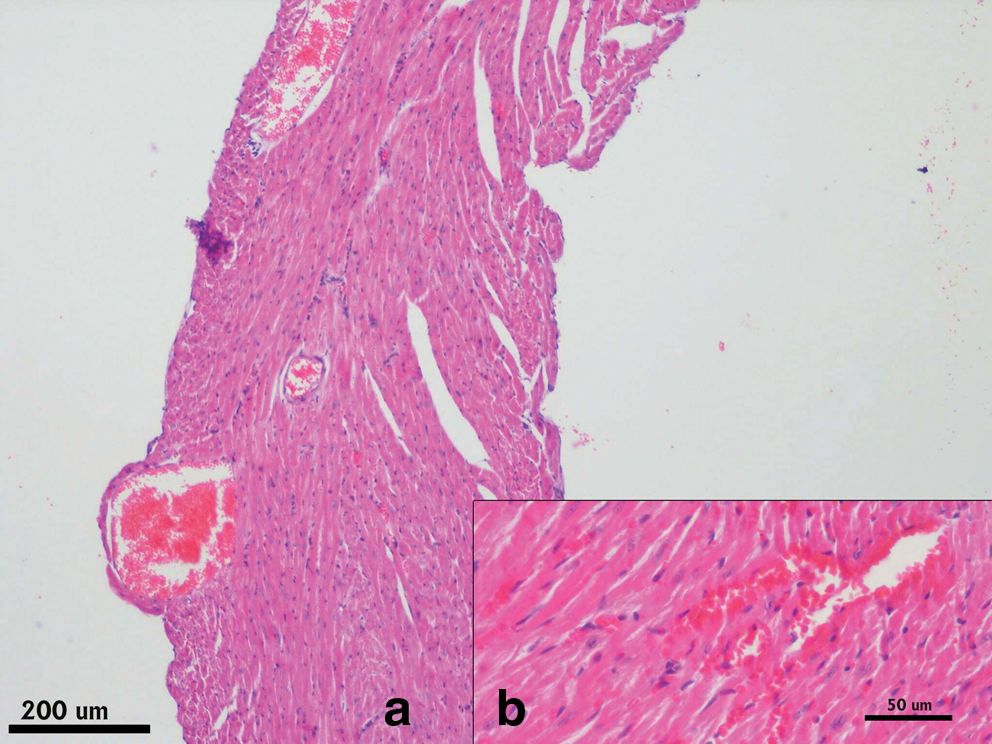
Photomicrograph of Day 21 heart. In Group III (GSE 50), there are no hemorrhagic foci, but limited hyperemia (arrow) is seen. Hematoxylin and eosin stain. Color images available online at
In Group IV (GSE 150), hemorrhagic foci are not seen. Hematoxylin and eosin stain. Color images available online at
(
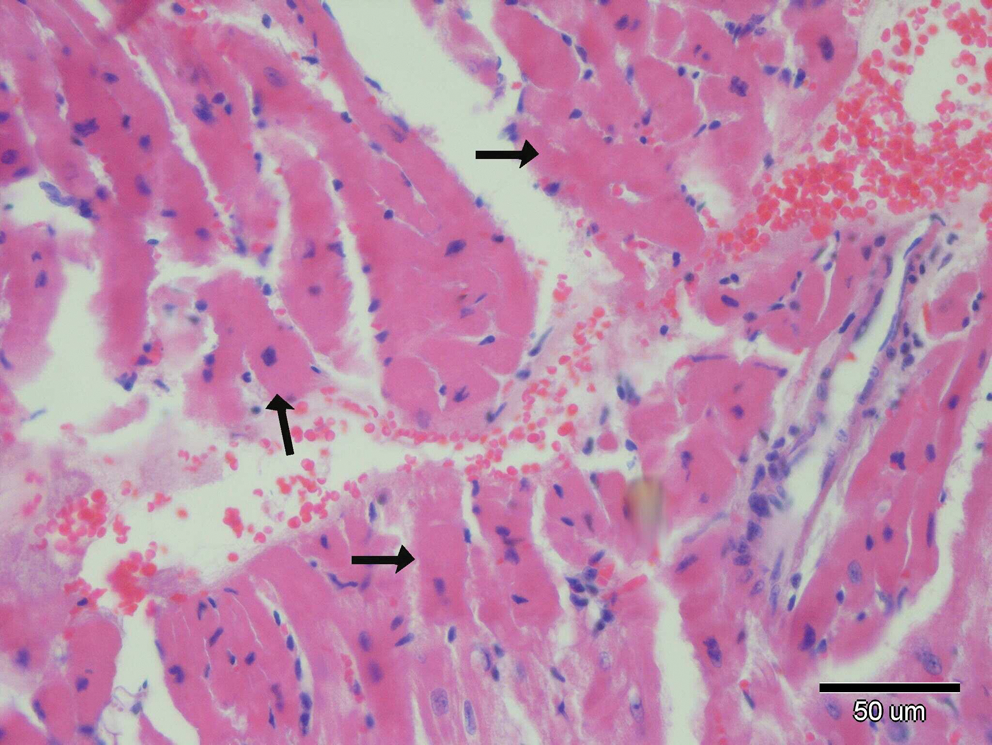
Microscopic changes in heart tissue of mice treated with DOX. In the DOX alone-treated group, interstitial hemorrhages and degenerative myofibrils (arrows) are seen. Hematoxylin and eosin stain. Color images available online at
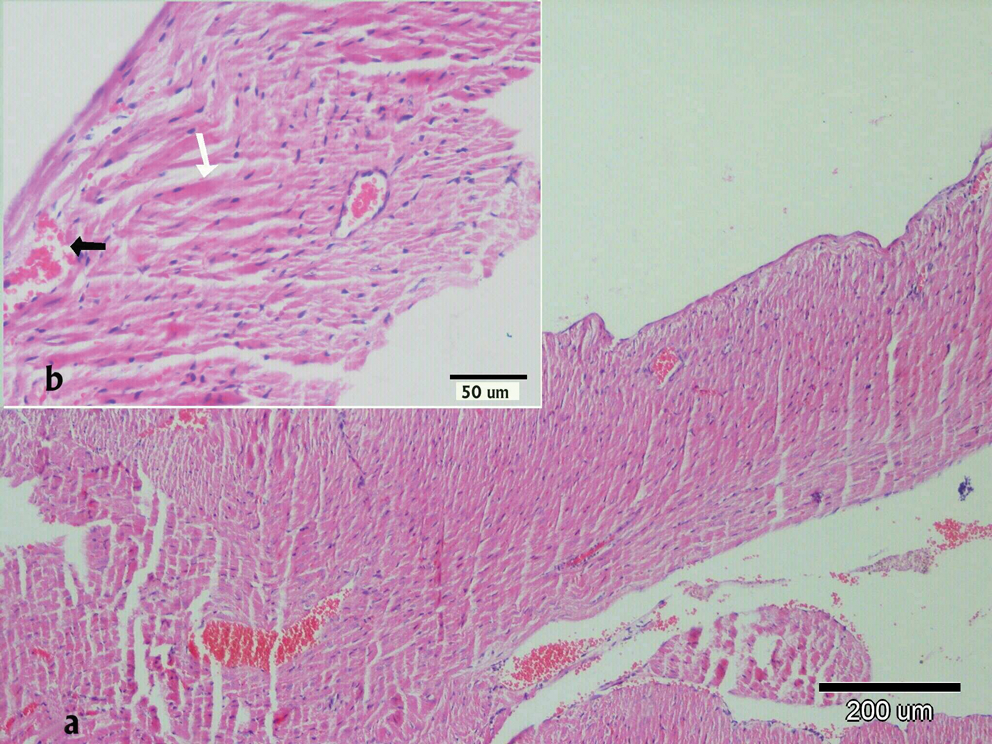
(
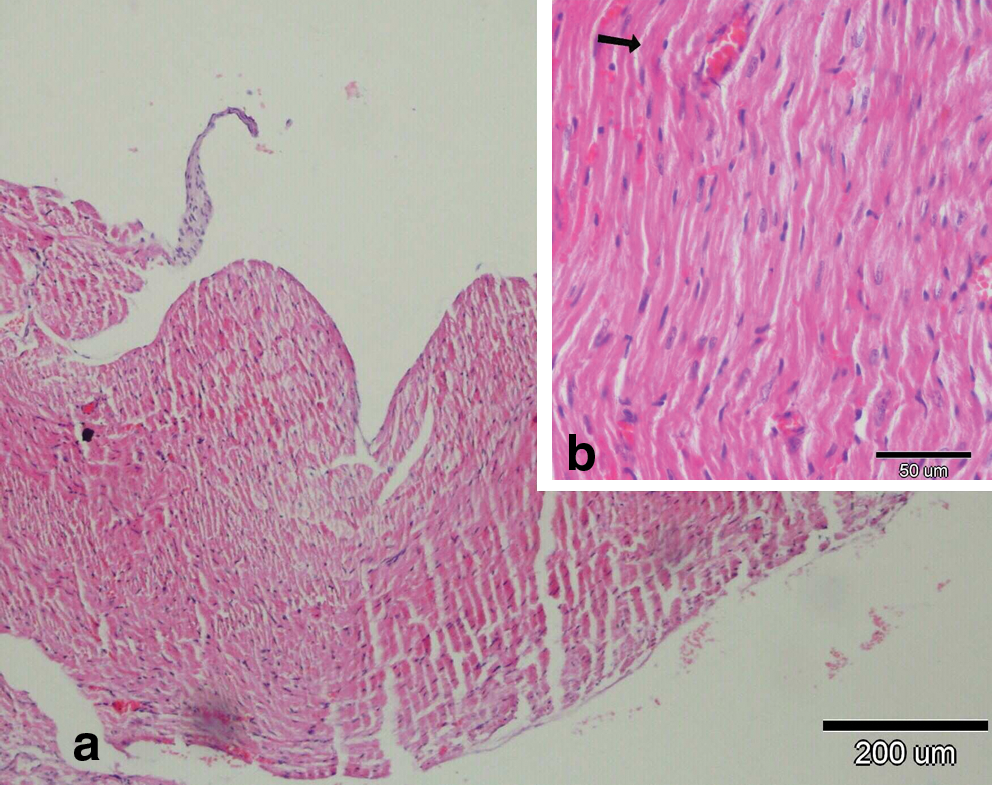
(
Changes were scored on the following scale: (−), no changes; (+), mild changes; (++), moderate changes; (+++), severe changes.
Discussion
In recent years, genetic monitoring of populations exposed to potential mutagens is an early warning system for genetic diseases such as cancer. The most frequently used genetic end points are CAs and MN frequency. 27,28 The CA and MN tests are reliable techniques for the evaluation of genotoxic effects induced by chemical pollutants, radiation, and drugs. 29 –31
In the present study, we confirmed the importance of CA and MN tests for evaluating of genotoxicity in mice exposed to a cumulative dose of DOX of 15 mg/kg of body weight. Our data showed that DOX administration induced an increase in the frequency of CAs and abnormal metaphases in bone marrow metaphases. DOX also significantly decreased the value of MI. As a result of exposure to 15 mg/kg of body weight DOX, a significant stimulation in the frequency of CAs such as chromatid breaks, dicentrics, acentric fragments, and chromatid gaps in bone marrow cells was observed. The maximum level of CAs incidence was observed in Group II, exposed to only DOX, and the lowest frequencies were observed in GSE-supplemented Group VI. Compared to DOX-treated mice, the number of CAs was always less in GSE + DOX-treated groups. In other words, the combined effect of GSE and DOX resulted in a significant decrease of CAs and abnormal metaphase rates, and again an increase in MI was observed. The 50 and 150 mg/kg of body weight doses of GSE decreased the number of DOX-induced structural CAs significantly, and the effect was dose-related. This resulted in decreases of 32% and 53%, respectively, in the frequency of induced CAs, particularly as chromatid breaks. It is considered that DOX cytotoxicity may be mediated by free radicals derived from this drug. These reactive oxygen species may then oxidize proteins, lipids, and nucleic acids and potentially cause DNA strand scission. In this way, DOX could induce mutations and CAs in normal and tumor cells. 6 These results support the findings of other researchers. Takeuchi et al. 32 showed the presence of chromosomal damage induced by the antitumor agent DOX in Wistar rats. In a similar study, a significant increase in the frequency of CAs and abnormal metaphases in Wistar rats treated with DOX alone was determined. 6 Gülkac et al. 7 observed that a cumulative dose of DOX of 90 mg/kg of body weight caused an increase in the frequency of CAs such as gaps, chromatid breaks, isochromatid breaks, exchanges, triradial figures, acentric fragments, and translocations.
MNs are cytoplasmic chromatin masses with the appearance of small nuclei that originate from chromosome fragments or whole chromosomes that are not included in the main daughter nuclei during metaphase or anaphase phase of cell division, and they can be easily recognized in the cytoplasm of erythrocytes. 33 In the present study, the frequency of MNs was increased in erythrocytes of DOX-treated mice for 6 consecutive days. The pretreatment of animals with GSE doses caused a significant reduction in the frequency of MNs compared to the group treated with DOX alone. There was a clear relationship between the MN frequency and dose of GSE. The MN frequency decreased with the increase of GSE dose. Although the values decreased, the number of MNs was still higher than that of the control and GSE alone-treated groups. In previous studies, a significant increase in MN frequency in erythrocytes after exposure to a high DOX dose was determined. Venkatesh et al. 34 determined a dose-dependent elevation in the frequency of micronucleated polychromatic cells in bone marrow erythrocytes of mice treated with different concentrations of DOX. In a similar study, Resende et al. 35 observed a significant increase in the MN frequency in mice treated with a cumulative dose of DOX of 90 mg/kg of body weight. The MN test in vivo was found to be easy to perform, and its results were mainly in accordance with results from CA tests in vivo. Besides, there was a linear correlation between CA and MN levels.
In cells, GSH serves to remove reactive oxygen species such as H2O2 produced because of either cellular respiration or metabolism of toxic substances and protects cells from oxidative injuries. 36 A change in the GSH ratio is considered as an indicator of the oxidative state of the cell. 37 In the present study, there was a significant reduction in GSH levels of heart tissues, implicating the presence of oxidative tissue damage. GSE, as an antioxidant agent, ameliorated oxidative injury in the tissues and functional deteriorations. In our study, GSH levels of heart tissues were significantly decreased by DOX toxicity, and because of its antioxidant activity, GSE treatment reduced the DOX-induced oxidative injury and restored GSH levels significantly.
MDA is a good indicator of the degree of lipid peroxidation. 38 MDA is a product generated during the oxidative breakdown of certain macromolecules such as lipids and is found either in free form or bound to certain tissue structures. Endoperoxides are generated as a result of changes that occur in the molecular structure of fatty acids during their breakdown, and MDA is generated during the breakdown of endoperoxides. MDA is considered to be the most significant indicator of membrane lipid peroxidation, arising from the interaction of reactive oxygen species with cell membranes. Because of the high susceptibility of lipid membranes to peroxidation, the free radicals easily peroxidated the lipid membranes, and MDA was generated as a final product of peroxidation. MDA also causes peroxidation itself and accelerates peroxidation by the means of synergy with free radicals. 39 In the present study, we found a significant increase in MDA levels during DOX toxicity, which is in agreement with previous studies. The increase in MDA levels in heart tissues suggests enhanced lipid peroxidation leading to tissue damage and failure of antioxidant defense mechanisms. In histological sections of the heart we have observed that DOX induced severe hyperemia, myofibril degeneration, and hemorrhage in myocardium. There was a significant decrease in the severity of hemorrhage in GSE-treated groups. However, a limited decrease was seen in the severity of myocardial degeneration in Group IV. Our results also show that GSE treatment significantly inhibits MDA production, implying a reduction in lipid peroxidation and cellular injury that protect the tissues against DOX-induced oxidative damage. The effect of DOX on GSH and MDA has also been described by other authors. For example, Reddy et al., 2 Öz and Ilhan, 40 and Narin et al. 41 reported a significant decrease in GSH levels and increase in MDA levels of the kidney, lung, liver, brain, and heart tissues.
In conclusion, the present study demonstrated that administration of GSE greatly attenuated the DOX-elicited changes in the levels of lipid peroxidation and oxidative stress biomarkers (MDA and GSH) in myocardial tissues and significantly reduced the frequency of CAs and MNs in bone marrow and erythrocyte cells. The protective effect of GSE on toxicity induced by DOX may be explainable with the antioxidant capacity of GSE, which is a vegetable product. Previous studies have shown that proanthocyanidin-rich extracts from grape seeds contain several type of polyphenolic components such as dimers, trimers, tetramers, and oligomers and monomeric flavanols like catechins and epicatechin derivatives that have antioxidant, anticancerogenic, and anti-inflammatory properties. 16 Also, many investigators have demonstrated the efficacy of GSE as an inhibitor of lipid peroxidation and as a powerful free radical scavenger in vitro as well as in vivo. Similarly, GSE was found to reduce the oxidation of polyunsaturated fatty acids in mouse liver microsomes and provide protection against lipid peroxidation and DNA fragmentation in mice. 14 Zhao et al. 42 suggested that dimerization and trimerization of monomers (catechins and epicatechins) could lead to anticarcinogenic effects in skin tumorigenesis possibly because of increase in their antioxidant activity. In a similar study, Arii et al. 43 have shown that oral administration of GSE in diet to mice inhibits APC mutation-associated intestinal adenoma formation. Raina et al. 15 showed that GSE inhibits in vivo growth of DU145 xenograft in nude mice via an inhibition of cell proliferation and an induction of apoptosis.
Because of all the above-mentioned reasons, antioxidant activity of GSE might be responsible for the protection against cardiotoxicity and genotoxicity induced by DOX. Therefore, the antioxidant role of GSE may be used as a “toxicity-limiting agent” to reduce side effects of cancer pills in the near future.
Footnotes
Acknowledgment
This study was supported by grants from the Giresun University Scientific Research Projects Department.
Author Disclosure Statement
No competing financial interests exist.