
Abstract
Regular consumption of fruits and vegetables is strongly associated with a reduced risk of cardiovascular disease (CVD). This effect occurs, in part, because of the plethora of bioactive agents in foods and their subsequent function as antioxidants. Ergothioneine (ERT), a novel antioxidant, is present in edible mushrooms and is not synthesized, but is accumulated, by humans through diet. In this study, we tested whether ERT, a bioactive agent, could interrupt pro-inflammatory induction of adhesion molecule expression associated with atherogenesis. Human aortic endothelial cells (HAECs) were incubated with increasing concentrations of ERT (0.01–10.0 mM) overnight (16 hours) followed by incubation with medium alone or with the pro-inflammatory cytokine interleukin (IL)-1β (5 ng/mL) for 6 hours to induce expression of vascular cell adhesion molecule-1 (VCAM-1), intercellular adhesion molecule-1 (ICAM-1), and endothelial-leukocyte adhesion molecule (ELAM-1 or E-selectin). ERT at 0.1–0.3 mM significantly (P < .05) reduced VCAM-1, ICAM-1, and E-selectin expression up to 41%. VCAM-1 was suppressed to the greatest extent followed by E-selectin and then ICAM-1. We next tested if binding of preloaded U937 human monocytes to HAECs would be inhibited. U937 binding to HAECs was significantly reduced in IL-1β-stimulated HAECs preincubated with 1 and 3 mM ERT. Unstimulated monolayers demonstrated marginal, but significant, reductions. ERT was not toxic to HAECs at any concentration used. These data provide evidence that ERT found in commonly consumed dietary mushrooms can protect against events observed in atherogenesis, suggesting increased dietary intake of edible mushrooms would be a prudent medicinal means of reducing CVD risk.
Introduction
E
CVD is one of the leading causes of death in the United States, with the annual death rate exceeding 1 million. 2 A hallmark of CVD is the formation of atheromas, or fatty deposits within aortic blood vessels, collectively termed atherogenesis. Atherogenesis is complex but clearly involves inflammation and oxidative stress, occurring primarily in the inner layer of blood vessels of the aortic endothelium. 8,9 Pro-inflammatory cytokines such as interleukin (IL)-1β, chemokines, and adhesion molecules (intercellular adhesion molecule-1 [ICAM-1], vascular cell adhesion molecule-1 [VCAM-1], and endothelial-leukocyte adhesion molecule-1 [ELAM-1 or E-selectin]) actively advance the disease by initiating an interaction between the vascular endothelium and pro-inflammatory immune cells such as monocytes. 10 This cellular interaction leads to a cascade of events including increased expression of adhesion molecules, migration of monocytes into the subendothelium, and, ultimately, foam cell formation leading to atheromas and atherosclerotic plaque. 9,11
Dietary antioxidants can protect against oxidative stress, inflammation, and cellular damage, which are all linked to diseases such as cancer and CVD. 5,10,12 Ergothioneine (ERT), a novel antioxidant, is found in edible mushrooms and may exert protection against many diseases, including CVD. 13 –15 ERT is an amino acid derivative of histidine (Fig. 1) and is not synthesized in humans, but levels increase in human blood as a result of dietary consumption at concentrations as low as 0.44 μM. 16 ERT has been reported in human tissues at concentrations of 1–4 mg/mL in blood (46–183 μM) and has been detected in the liver, kidney, seminal vesicles, bone marrow, optic lens, and central nervous system. 16 ERT concentrations have been quantified by high-performance liquid chromatography-mass spectrometry in both commonly consumed and specialty mushrooms in the range of 0.21–2.6 mg/g dry weight, with white button at the lower concentration and oyster at the higher level. 17,18 ERT functions as an antioxidant and inactivates numerous diverse free radicals, making it potentially one of the most effective hydrophilic bioactive molecules. Through its antioxidant activity in a pro-oxidant, pro-inflammatory environment, ERT can potentially reduce binding of monocytes to endothelial cells, lowering the risk for CVD. 19,20

Chemical structure of ergothionine.
Although research continues on elucidation of an ERT transporter and ERT's role as an antiinflammatory agent, little attention has been focused on the potential protective role of ERT against atherogenesis in a biological context or determination of a specific underlying mechanism. This is the first study to determine whether preincubation of human aortic endothelial cells (HAECs) with ERT could reduce pro-inflammatory-induced adhesion molecule expression. After analyzing VCAM-1, ICAM-1, and E-selectin expression, we assessed whether this translated into a significant reduction in binding of human monocytes to aortic endothelium in a pro-inflammatory environment.
Materials and Methods
Chemicals
Dimethyl sulfoxide, formaldehyde, and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were obtained from Sigma Chemical Co. (St. Louis, MO, USA). Plasticware (plates and flasks) was purchased from Corning, Inc. (Corning, NY, USA). All other chemicals and cell culture reagents were obtained from Sigma Chemical Co.
Cell culture
HAECs were purchased from Clonetics Laboratories (San Diego, CA, USA) and cultured in endothelial cell basal medium (Clonetics). The medium contained 10% fetal bovine serum, hydrocortisone, vascular endothelial growth factor, insulin-like growth factor, fibroblast growth factor, gentamicin, epidermal growth factor, heparin, and ascorbic acid at concentrations provided as a kit by the manufacturer (Clonetics). HAECs were seeded in 1% gelatin (Sigma)-coated T-75 flasks (Corning) and 24- and 96-well plates (Corning). Medium was changed every other day, and cells were subcultured through trypsinization and reseeded in flasks to propagate. For experiments, cells grew to confluence, and each assay was replicated three or four times in quadruplicate using passages 6–8. To deliver, water-soluble ERT (Sigma) was dissolved in endothelial cell basal medium and filtered (filter disk; pore size, 0.22 μm) to produce final working concentrations (0.01–10 mM). U937 monocytes (ATCC, Rockville, MD, USA) were grown in suspension culture in RPMI-1640 medium (Sigma) supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin. Medium was exchanged every 3 days, and cells were subcultured weekly.
Cytotoxicity of ERT
Monolayers of HAECs were incubated overnight with increasing concentrations of ERT and then washed twice with phosphate-buffered saline (PBS) before addition of phenol red-free medium containing MTT (5 mg/mL). Cultures were incubated for 4 hours at 37°C with subsequent centrifugation of the plate for 10 minutes (1,500 g at 25°C) to pellet reduced MTT formazan crystals. Medium was aspirated, dimethyl sulfoxide (0.1 mL per well) was added, samples were mixed (15 minutes at 25°C) to solubilize MTT, and solutions were analyzed at 560 nm. Lack of cytotoxicity was corroborated by light microscopy.
Adhesion molecule expression
HAECs were cultured in 24-well plates until confluent and were incubated overnight with increasing concentrations of ERT dissolved in cell culture medium. After incubation, monolayers were washed twice with PBS (0.5 mL per well) and further incubated with medium alone or with IL-1β (5 ng/mL) for 6 hours at 37°C. Medium was removed, and monolayers were fixed with 1% formaldehyde at 25°C for 30 minutes. Monolayers were washed with PBS and blocked with 10% fetal bovine serum in PBS for 1 hour to reduce nonspecific binding and background. Monoclonal antibodies against human VCAM-1, ICAM-1, and E-selectin (BD Pharmingen, San Diego) were added at 5, 1, and 10 μg/mL, respectively, in PBS with 10% fetal bovine serum for 1 hour at 25°C. The secondary antibody, horseradish-conjugated anti-mouse immunoglobulin G (Santa Cruz Biotechnology, Santa Cruz, CA), was added at a 1:400 dilution and incubated at 25°C for 2 hours. Subsequently, after washing with PBS, horseradish peroxidase substrate was added, and cells were incubated for 1 hour to develop the colorimetric end point (Bio-Rad, Hercules, CA, USA). The absorbances were read at 405 nm using a SpectraFluor multiwell plate reader (Tecan, Research Triangle Park, NC, USA).
Fluorescent labeling of monocytes
U937 cells, a human monocyte cell line, was used to test adherence to HAEC monolayers. 21 Prior to co-incubation, U937 cells were fluorescently labeled by incubating cells (2 × 106 cells/mL) with 5 μM 2′,7′-bis-(2-carboxyethyl)-5 (and 6)-carboxy-fluorescent acetoxymethyl ester (Molecular Probes, Eugene, OR, USA) in endothelial cell basal medium for 30 minutes at 37°C and 5% CO2 as described previously. 7 2′,7′-Bis-(2-carboxyethyl)-5 (and 6)-carboxy-fluorescent acetoxymethyl ester, a nonfluorescent lipophilic compound, readily enters cells where it is modified to become fluorescent and retained by cells. 2′,7′-Bis-(2-carboxyethyl)-5 (and 6)-carboxy-fluorescent acetoxymethyl ester was prepared as a 1 mg/mL stock in dimethyl sulfoxide and stored at −80°C. After labeling, cells were washed twice with PBS containing 1% fetal bovine serum to remove excess dye. U937 cells were then resuspended in medium (5 × 105 cells/mL) and added (0.5 mL per well) to HAEC monolayers as described below.
U937 monocyte binding to HAECs
HAECs were cultured in gel-coated 24-well plates until confluent and then incubated overnight with increasing concentrations of ERT dissolved in cell culture medium. After incubation, monolayers were washed twice with PBS (0.5 mL per well) and further incubated with IL-1β (5 ng/mL) for 6 hours at 37°C. U937 cells, labeled as described above, were incubated with HAECs for 30 minutes at 37°C and 5% CO2. After incubation, nonadherent cells were removed by washing twice with PBS containing 1% fetal bovine serum. The attached cells were lysed with 0.5 mL of 50 mM Tris buffer (pH 7.6) containing 1% sodium dodecyl sulfate. The fluorescent intensity was measured at excitation and emission wavelengths of 485 and 535 nm, respectively, with a Tecan Spectrafluor fluorescence multiwell plate reader.
Statistical analysis
Data were analyzed using Student's t test to compare individual treatment groups with the respective controls. Results are given as mean ± SEM values, and P < .05 is considered significant.
Results
We first tested whether increasing concentrations of ERT would cause cytotoxicity and/or contribute to reduced cell viability. Using the MTT tetrazolium assay, we did not observe toxicity or reduced viability at concentrations up to 1 mM compared to control cultures (Fig. 2). We also did not note any morphological differences between control and treated cells when monolayers were examined by microscopy.
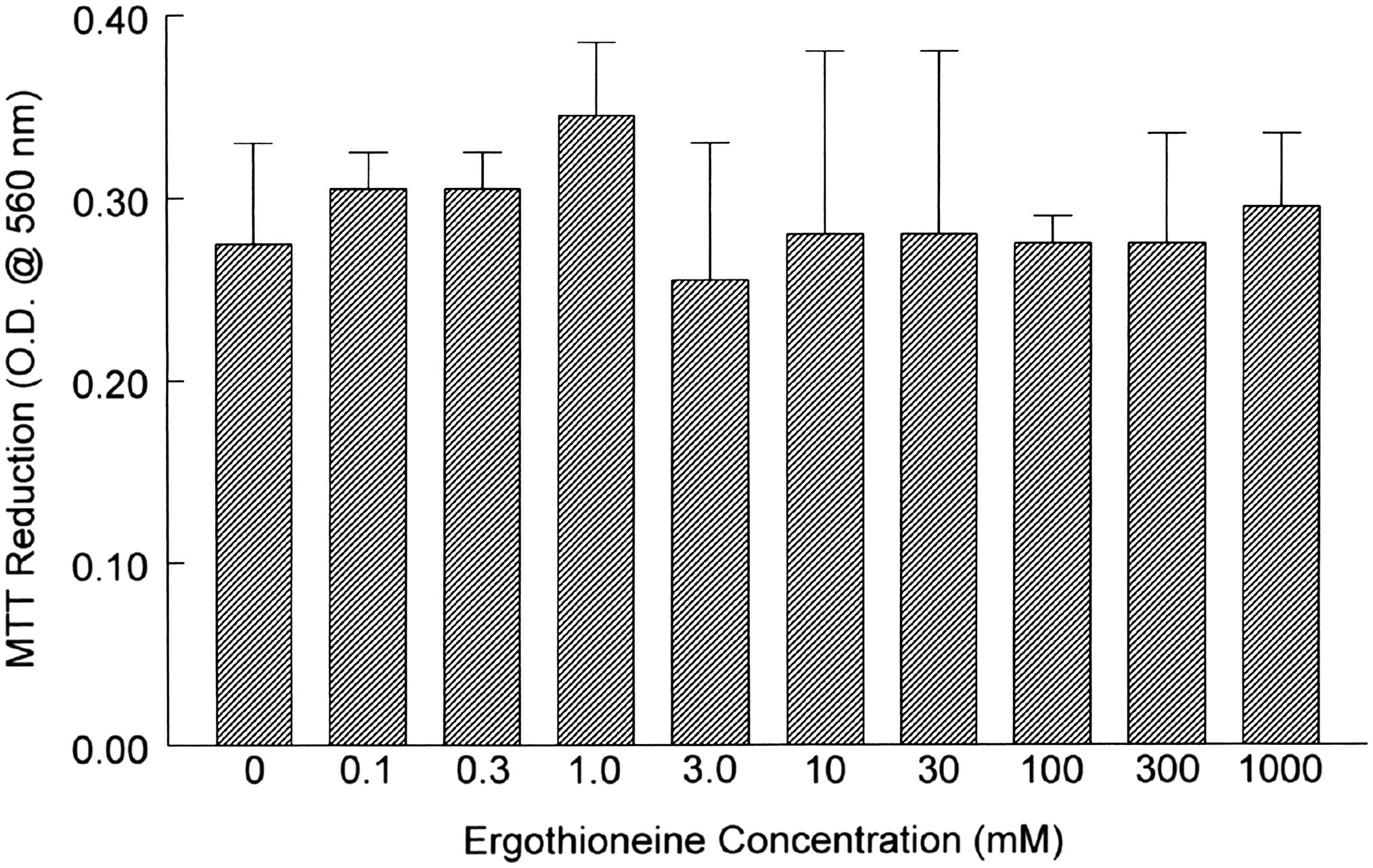
Viability of human aortic endothelial cells incubated overnight with increasing concentrations of ergothioneine. Monolayers were incubated overnight, washed with phosphate-buffered saline, and further incubated for 4 hours with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) tetrazolium reagent. At harvest, plates were centrifuged, phosphate-buffered saline was aspirated, and pelleted MTT-formazan crystals were solubilized in dimethyl sulfoxide before analyzing at 540 nm. Data are mean ± SEM values for optical density (O.D.) and are representative of three or four experiments, each performed in quadruplicate.
Increased expression of adhesion molecules and binding of monocytes to HAECs are critical events in atherogenesis. As a result, we first analyzed the expression of VCAM-1, a pivotal and commonly measured adhesion molecule, after preincubation with medium alone or containing increasing concentrations of ERT. After stimulation with IL-1β for 6 hours, VCAM-1 expression increased 2.4-fold (Fig. 3). After preincubation with ERT, VCAM-1 expression was marginally, but significantly, reduced by 10% at 0.3 mM. VCAM-1 expression was further suppressed in a dose-dependent manner up to 41% after preincubation with 10 mM ERT with no observed cytotoxicity. Basal expression of VCAM-1 was also significantly reduced after preincubation with 3 and 10 mM ERT by 7% and 39%, respectively.

Vascular cell adhesion molecule-1 expression in human aortic endothelial cells. Monolayers were incubated overnight with increasing concentrations of ergothioneine. Cells were then incubated with medium alone or interleukin (IL)-1β (5 ng/mL) for 6 hours. After harvesting, vascular cell adhesion molecule-1 expression was determined by enzyme-linked immunosorbent assay. Data are mean ± SEM values for O.D. and are representative of three or four experiments, each performed in quadruplicate. *P < .05 versus the respective control (CTL).
We next analyzed the expression of ICAM-1. Basal ICAM-1 expression was higher than either VCAM-1 or E-selectin, and incubation with IL-1β further increased expression by twofold, as expected (Fig. 4). After preincubation with ERT, ICAM-1 expression was marginally, but significantly, decreased at concentrations as low as 0.1 mM. Maximal suppression was significant (P < .05), although marginal, at 10% using 1 mM ERT. Basal ICAM-1 expression was not significantly modulated by ERT alone.
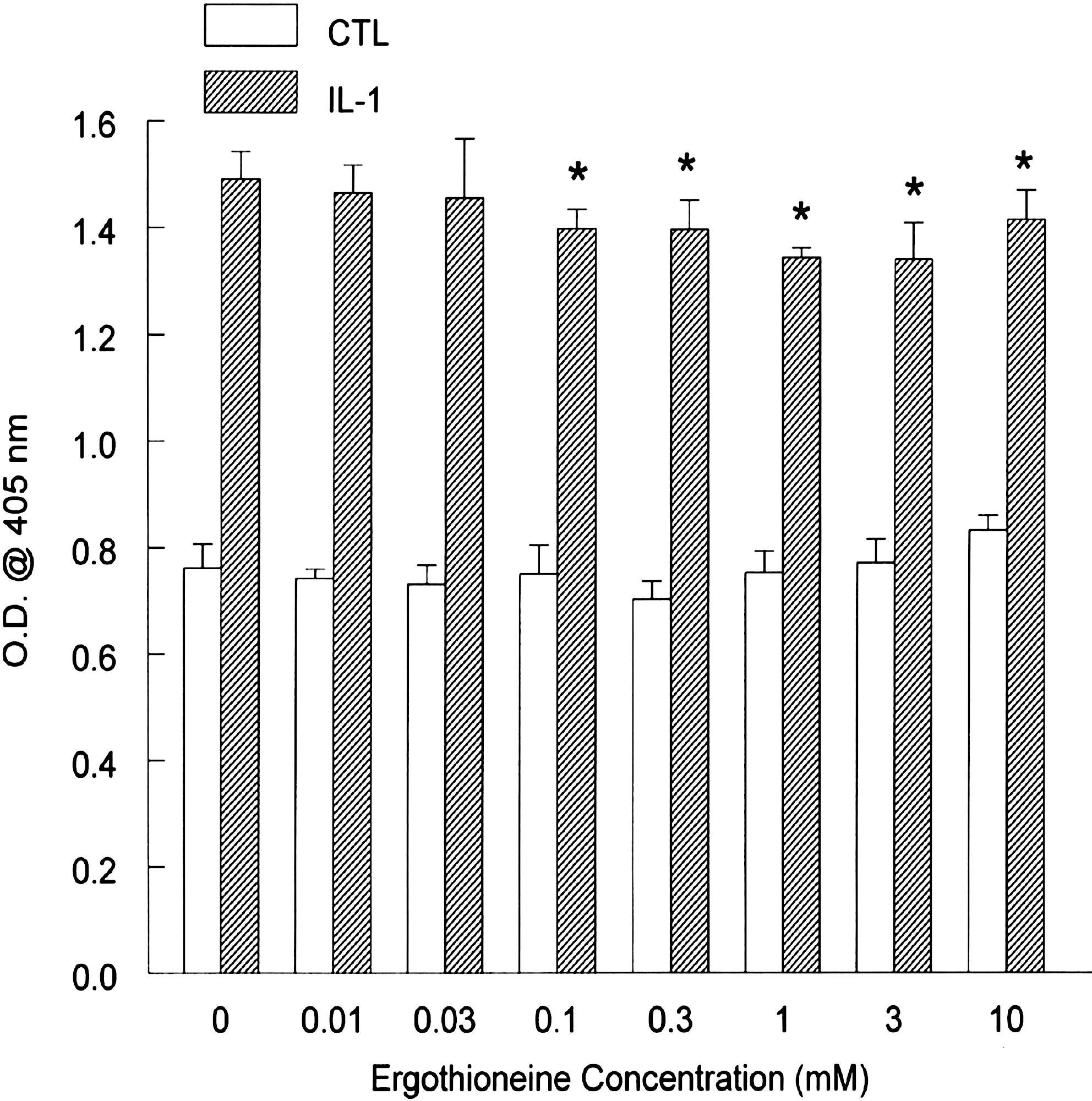
Intercellular adhesion molecule-1 expression in human aortic endothelial cells. Monolayers were incubated overnight with increasing concentrations of ergothioneine. Cells were then incubated with medium alone or IL-1β (5 ng/mL) for 6 hours. After harvesting, intercellular adhesion molecule-1 expression was determined by enzyme-linked immunosorbent assay. Data are mean ± SEM values for O.D. and are representative of three or four experiments, each performed in quadruplicate. *P < .05 versus the respective CTL.
We next analyzed the expression of the adhesion molecule E-selectin. Basal expression remained low and unchanged in cultures until preincubation of cells with ERT at 1 and 10 mM, where reduction was significantly reduced up to 31% (Fig. 5). After incubation with IL-1β, E-selectin expression increased by eightfold. After preincubation with ERT, E-selectin expression was significantly reduced by > 10% at concentrations as low as 0.1 mM. Peak suppression occurred at 1 mM with a reduction of 26%, which remained constant up to 10 mM.
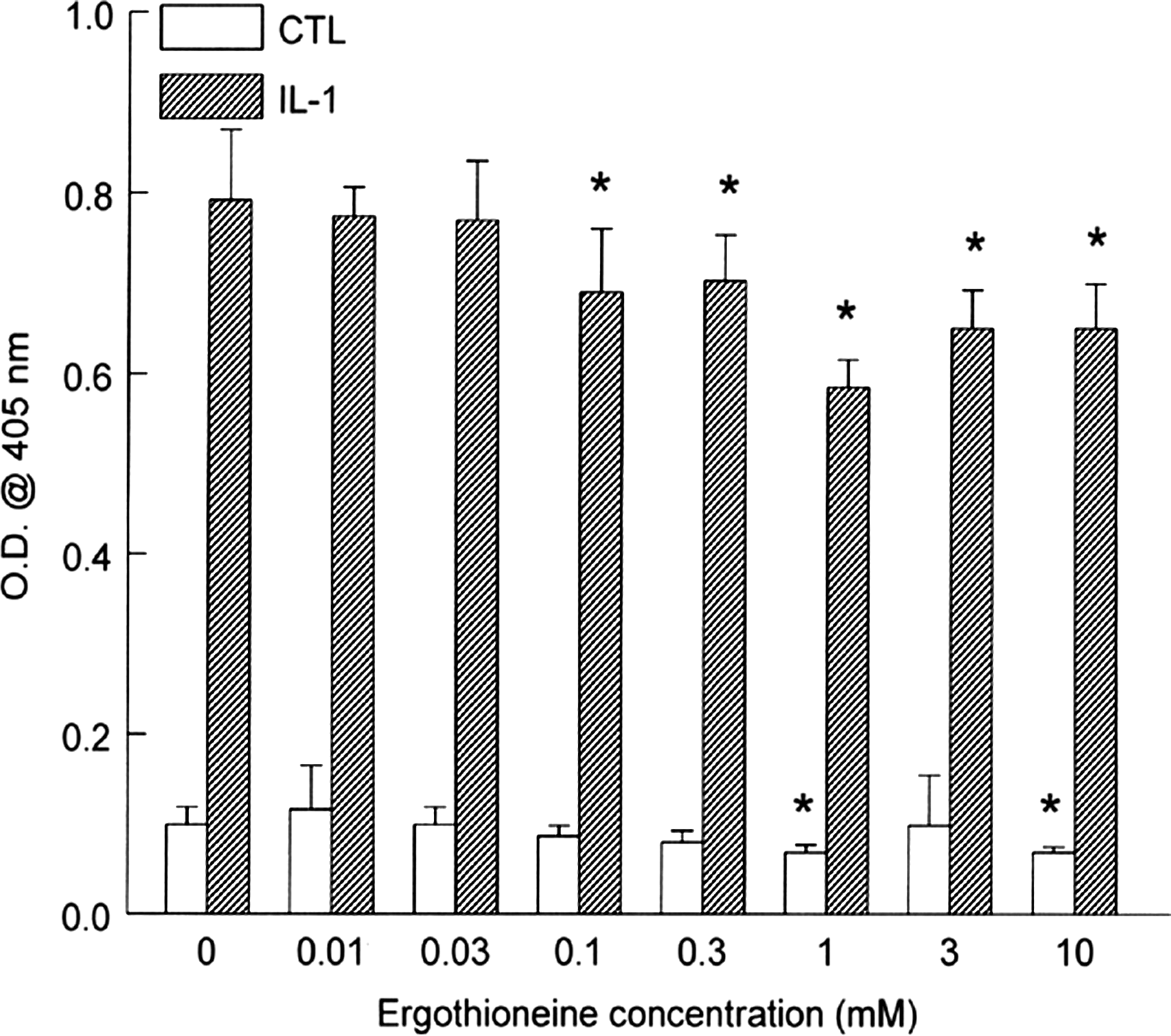
E-selectin adhesion molecule expression in human aortic endothelial cells. Monolayers were incubated overnight with increasing concentrations of ergothioneine. Cells were then incubated with medium alone or IL-1β (5 ng/mL) for 6 hours. After harvesting, E-selectin expression was determined by enzyme-linked immunosorbent assay. Data are mean ± SEM values for O.D. and are representative of three or four experiments, each performed in quadruplicate. *P < .05 versus the respective CTL.
We next analyzed the capacity of ERT to prevent or attenuate the binding of preloaded U937 monocytes to the endothelium, viz., HAECs. Based on the previous dose–response curves and reported physiological levels, we selected 1 and 3 mM as test concentrations of ERT for further study. Preincubation with ERT reduced adhesion of monocytes to unstimulated HAECs significantly by 15% at 3 mM but not at 1 mM ERT (Fig. 6). Incubation with IL-1β increased monocyte adherence to HAECs by 2.5-fold. Preincubation with 1 and 3 mM ERT significantly reduced U937 adhesion to HAECs by 12% and 27%, respectively.
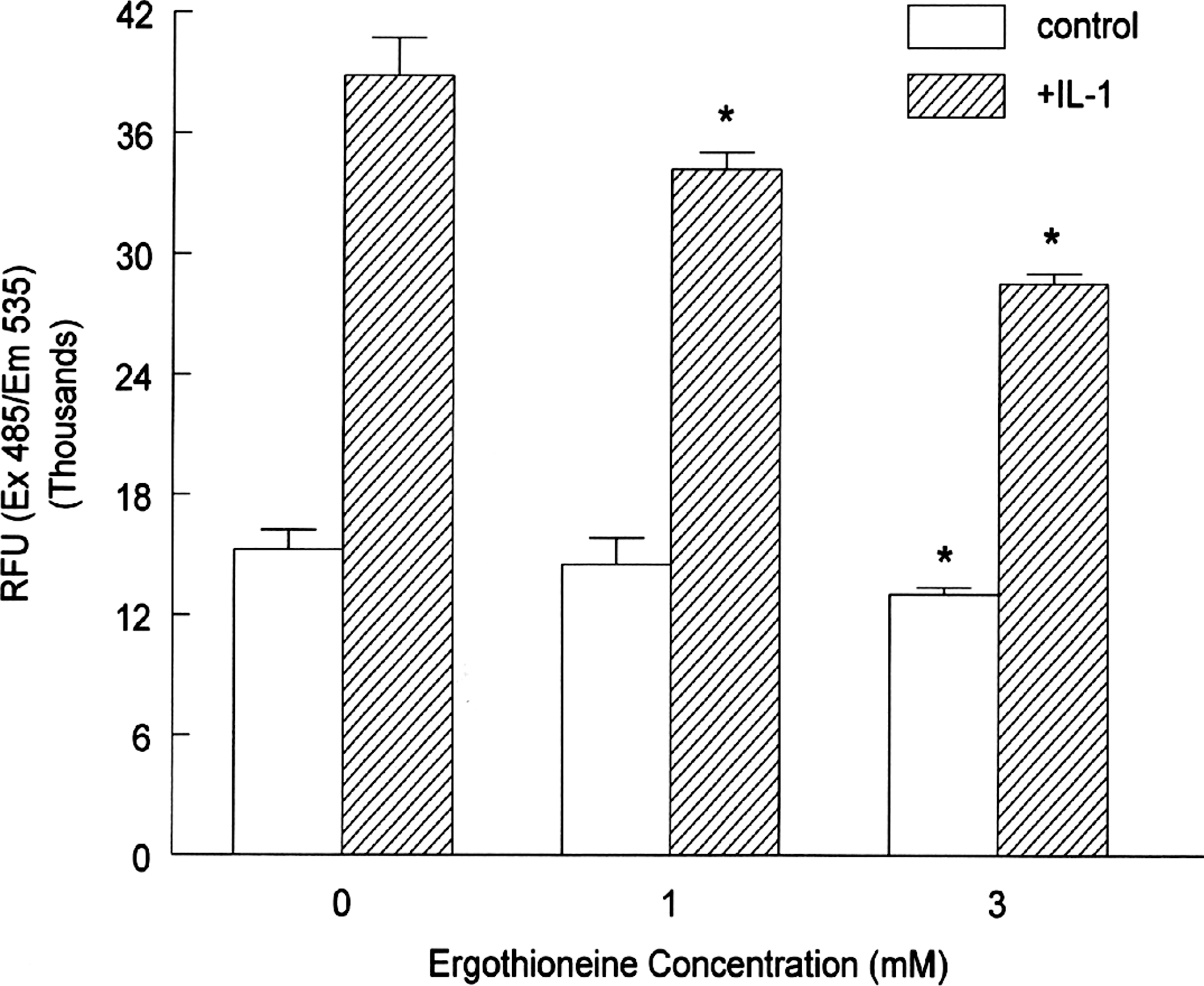
Binding of U937 human monocytes to human aortic endothelial cells preincubated with ergothioneine. Human aortic endothelial cells were incubated overnight with the indicated concentrations of ergothioneine followed by incubation with medium alone or with IL-1β (5 ng/mL) for 6 hours. Fluorescently labeled U937 cells were co-incubated with human aortic endothelial cells for 30 minutes at 37°C. After washing, cells were harvested, and fluorescence was quantitated to determine binding. Data are mean ± SEM values for relative fluorescent units (RFU) and are representative of three or four experiments, each performed in triplicate. *P < .05 versus the respective control. Ex, excitation; Em, emission.
Discussion
This study is the first to demonstrate the capacity of ERT, a bioactive antioxidant in edible mushrooms, to reduce adhesion molecule expression with subsequent reductions in monocyte adhesion to HAECs. In this study, we have demonstrated that ERT significantly reduced cell surface expression of VCAM-1, ICAM-1, and E-selectin. Furthermore, there was a significant reduction in adhesion of human monocytes to unstimulated and pro-inflammatory, cytokine-stimulated HAECs. The health implications are that ERT can be protective against CVD by interfering with events that contribute to atherogenesis. This would support the use of ERT-rich dietary mushrooms as functional foods and medicinal agents.
As a molecule, ERT has received little attention regarding its potential role in protecting against heart disease. However, its clear capacity to function as an antioxidant coupled with the well-accepted role of oxidative stress in the etiology of CVD supports such a notion. Furthermore, intracellular pro-atherogenic events and cell adhesion expression are modulated by the thiol/disulfide redox state. 22 Thus, it is intriguing that ERT differs from other thiols in that it exists primarily as a thione under physiological conditions, which intuitively would seem to limit its antioxidant activity and potential efficacy. The thione/thiol tautomerism exhibited by ERT does confer differences in reactivity toward other molecules compared to other biological thiols such as glutathione. 16 However, studies have shown that ERT possesses antioxidant activity with a Trolox equivalent antioxidant capacity value of 0.87 mmol of Trolox/L, a ferric reducing antioxidant power value of 0.89 mmol of Fe/L, 20 and a hypochlorite (HOCl) scavenging capacity value of 70.68 μM (50% inhibitory concentration). 23 In chemical studies, ERT scavenged superoxide anion radical and singlet oxygen in ultraviolet B-irradiated fibroblasts. 24 Additionally, ERT reacts with the hydroxyl radical in a biologically relevant manner and cooperates with ascorbic acid to protect biological systems against reactive oxygen species. 25,26
The capacity of bioactive agents, and particularly antioxidants, to reduce expression of adhesion molecules has been reported in HAECs, a nontransformed primary endothelial cell line derived from humans. In this study, clear reductions in adhesion molecule expression of up to 41%, 10%, and 26% are demonstrated for VCAM-1, ICAM-1, and E-selectin, respectively, in HAECs. In other studies, the antioxidant vitamin E, viz.,
The capacity of diverse antioxidants to suppress adhesion molecule expression and monocyte binding has been reported in other cell models with varying degrees of specificity and magnitude. For example, N-acetyl-
In this study, concentrations of ERT as low as 0.1–0.3 mM (22.9–68.7 μg/mL) reduced adhesion molecule expression, but monocyte adhesion was reduced significantly at a higher concentration of 1 mM (229.3 μg/mL). This has also been demonstrated in HAECs using vitamin E, where
The data also suggest the presence of an ERT transporter in HAECs. The ERT transporter functions in many cell types to accumulate ERT to high levels and promote cellular retention. 37 Animals and humans store ERT as a free form in tissues that are known to be exposed to oxidative stress, including red blood cells, seminal fluid, ocular lens, liver, and kidneys, at levels that can reach the millimolar range comparable to the range used in this study. 16 It seems intuitive that endothelial cells, a site of pro-atherogenic and inflammatory oxidative stress, would also accumulate ERT. In these studies, it has been clearly demonstrated that preincubation of HAECs with ERT confers anti-atherogenic bioactivity, presumably due to cellular accumulation.
In this report, dietary ERT, found predominantly in edible mushrooms, interfered with processes clearly linked to atherogenesis and CVD. This is the first report to demonstrate these effects in an in vitro model of atherogenesis. The finding that dietary bioactive agents such as ERT are protective against human diseases, i.e., CVD, further supports the notion that consumption of fruits and vegetables, and particularly ERT-rich mushrooms, is an important and efficacious cardioprotective approach.
Footnotes
Acknowledgments
This work was funded by The Mushroom Council and the Australian Mushroom Growers Association.
Author Disclosure Statement
The author affirms that there are no commercial associations that might create a conflict of interest in association with this manuscript submission. No competing financial interests exist.