
Abstract
The prognosis for patients with metastasized prostate cancer is still poor, despite conventional aggressive therapeutic modalities. Several in vitro studies together with animal models and epidemiological studies have indicated that phytochemicals can be antitumorigenic and may be protective against human cancers. However, the potential antitumor effects of genistein isoflavone, a widely studied nutrient phytochemical, have been equivocal. In this study, we investigated the effects of genistein-selenium (Gn-Se) combination on chemosensitivity and matrix metalloproteinase-2 (MMP-2) expression levels in PC3 (hormone-independent) and LNCaP (hormone-dependent) prostate cancer cells. 3-(4,5-Dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium staining and ATP bioassay showed that genistein, selenium, and Gn-Se combination significantly inhibited growth of LNCaP and PC3 cells in a dose- and time-dependent manner, independent of hormonal status, and with no significant differences in chemosensitivity between LNCaP and PC3. Gn-Se combination induced significantly the greatest growth inhibition in both cell lines. Growth inhibition was through apoptosis induction. The treatment-induced apoptotic cascades are caspase-dependent, with evidence of an alternative non-caspase pathway(s). Treatment also induced a dose- and time-dependent decrease in MMP-2 expression levels in PC3 and LNCaP with no significant differences between the two cells. Gn-Se combination induced the greatest depression in MMP-2. Overall, none of the treatment modalities had any significant inhibitory effect in normal prostate epithelial cells. The data obtained from the present study indicate that Gn-Se combination may have chemopreventive value and/or may be adjuvant to standard therapy for prostate tumors independent of hormonal status. MMP-2 expression in cancer cells has been associated with active invasion and metastasis.
Introduction
T
In this study our focus was on dietary nutrient strategies: the phytochemical genistein (Gn) isoflavone and the mineral selenium (Se). We examined the potential effects of Gn-Se combination on two human malignant prostate cancer cell lines, PC3 (hormone-independent) and LNCaP (hormone-dependent) phenotypes. These nutrients are known to induce apoptosis in various tumor cells via different mechanisms of action.
Gn (4′,5′,7-trihydroxyisoflavone) is a major isoflavone constituent of soy and soy products. 9,10 Because of its structural similarity to estrogen, Gn has the ability to bind estrogenic receptors and can therefore regulate cell proliferation and prevent hormone-dependent growth of cancer cells. 11,12 Gn also exhibits several pharmacological effects, including cell cycle inhibition by arresting the cell at the G2/M phase, 12 –14 inhibition of tyrosine kinases, DNA topoisomerases, and anti-angiogenesis, 11,12,15 decreasing oxidative DNA damage, 16 and lowering prostate cancer risk. 17 Research in animal models has revealed that dietary phytoestrogens can inhibit growth and decrease androgen receptor expression in rat prostates 18 as well as reduce tumor cell proliferation, increase apoptosis, and inhibit angiogenesis in mice. 19 Gn has also been shown to alter growth factor signaling in rat prostates. 15,20 Unlike other isoflavonoids, the concentration at which Gn exerts its toxicity is far above that at which it exerts biological and pharmacological effects. This property makes Gn highly favored as a potential chemopreventive agent. 12
Se is of nutritional significance as an essential trace element in human nutrition. Broad scientific and several anecdotal evidence from various health centers has revealed the importance of Se in human health. A number of animal models and epidemiological studies indicate that Se can have a protective effect against human cancer. 21 –25 Epidemiological studies by the National Cancer Institute and long-term observational studies indicated/revealed the potential of selenium in significant reduction of prostate cancer risk and proliferation 26,27 in humans. The mechanism of the anticarcinogenic activity of Se has not been fully elucidated, but there is evidence that induction of apoptosis, regulation of p53, and modulation of androgen-regulated genes are among the possible mechanisms. 25,28 –30 Furthermore, Se is a major component of many antioxidant enzymes. Such observations strongly indicate the potential chemopreventive benefits of Se and the justification for further research.
In studying the molecular and cellular effects of Gn and Se on prostate tumor cells, a group of investigators reported synergistic effects of Gn-Se combination on apoptosis induction and associated signaling pathways (AKT phosphorylation, p21, Bax) in p53-expressing LNCaP and p53-null PC3 cells. 14 In preliminary studies in our laboratory, the 50% inhibitory concentration (IC50) for Gn-Se combination in both PC3 and LNCaP cells was significantly lower (P < .01) than the IC50 for either Gn or Se. From these observations, we believed that the combination could offer a greater and safer chemoprevention efficacy with minimum cytotoxicity. In this study, we investigated if Gn-Se combination induces growth inhibition in LNCaP (hormone-dependent) and/or PC3 (hormone-independent) prostate cancer cell lines and the potential mechanism of growth inhibition mediated by the intrinsic pathway(s) of treatment-induced apoptosis. The effect of treatment on matrix metalloproteinase-2 (MMP-2) expression was also determined. MMP-2 expression in cancer cells has been associated with active invasion and metastasis.
Materials and Methods
Cell lines and chemicals
The prostate cancer cell lines PC3 (hormone-independent) and LNCaP (hormone-sensitive) and NPtEC normal prostate epithelial cells were maintained in complete RPMI-1640 medium (Sigma-Aldrich Chemical Co., St. Louis, MO, USA), supplemented with 10% fetal bovine serum. Gn (Indofine Chemical Co. Inc., Somerville, NJ, USA) was dissolved in dimethyl sulfoxide as a 100 μg/L stock solution and diluted in RPMI 1640 culture medium at a final dimethyl sulfoxide concentration of 0.2% (vol/vol). Gn concentrations of 0, 10, 20, 30, 40, 50, 60, and 100 μM (Gn10–100) were prepared and stored at −35°C. Dimethyl sulfoxide alone was found to be nontoxic to these cells at 0.5%. Dimethyl sulfoxide and [3-(4, 5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) were purchased from Sigma-Aldrich. ATP and MTS assay kits were from Oncogene Research (San Diego, CA, USA). A.H. contributed the selenite, the form of Se used in this study.
Cell culture
Treatment-induced growth inhibition
The cells were cultured and treated with Gn, Se, or Gn-Se combination. For Gn treatment, cells were treated in triplicates with varying concentrations of Gn: 0, 10, 20, 30, 40, 50, and 60 μM (Gn0–60). For Se treatment, cells were treated in triplicates with 0, 1, 2, 3, 5, 10, and 20 μM (Se0–20). For combination treatment (Gn-Se), the cells were treated with Gn0–60-Se0–20 doses at a 1:1 (vol/vol) ratio (Table 1). The plates were cultured for 24 and 48 hours, respectively, and subjected to ATP and MTS bioassay to determine chemosensitivity of the cells to the treatments.
Gn, genistein; Se, selenium.
ATP assay
Treatment-induced cytotoxicity in the cells was determined using the ATP assay. ATP is a source of energy in viable cells and is required for many cellular processes, including apoptosis. Cells that are dying are unable to perform normal metabolic functions and can no longer manufacture ATP. Therefore, a decrease in ATP levels can indicate cell death, whereas an increase will indicate cell proliferation. The ATP assay is based on bioluminescent detection of ATP levels within the cell and offers a sensitive determination of even small amounts of ATP. The assay uses
The assay is optimized for fast determination of low levels of ATP. Concentrations down to 0.1 pmol can be accurately determined using the luminescent signal of the luciferase reaction.
In brief, each cell type (PC3, LNCaP, or NPtEC) was plated at a density of 1.5 × 104 cells per well in 96-well plates and incubated for 24–48 hours to achieve 80–90% confluence. The supernatants were discarded, and the adherent cells were treated in triplicates with varying concentrations of Gn (Gn0–60), Se (Se0–20), or Gn-Se combination or with 1% Triton X-100 as a positive control. The cells were then incubated for 24–48 hours under standard conditions of 5% CO2 and 37°C in a humidified atmosphere. After incubation, 50 μL of lysis solution was immediately added to each well and left for 5 minutes. Then 50 μL was transferred from each well of the 96-well plate to its corresponding well in a white plate, after which 50 μL of luciferin/luciferase solution was added to each well. Luminescence was determined after 5 minutes using a luminometer. ATP concentration was determined by comparison of luminescent values against a standard curve. Percentage inhibition was calculated using the following formula, and the IC50 was determined from the graph:
where Lum c is the luminescence of the control sample and Lum s is the luminescence of the treated sample.
MTS assay
To supplement/confirm the ATP results, MTS assay was performed to determine treatment-induced cytotoxicity in the cells. This assay uses MTS to determine cell viability. In the presence of viable cells, the substrate is reduced to a colored formazan product. In active mitochondria, the tetrazolium ring is cleaved in the presence of dehydrogenase enzymes, producing a product that is soluble in tissue culture medium. Therefore, the color intensity is directly proportional to the number of viable cells. Relative cell numbers can therefore be determined based on optical absorbance from each well of microtiter plate. In brief, cells were plated at a density of 1.5 × 104 cells per well in a 96-well plate and incubated for 24 hours to achieve confluence. After incubation, the cells were treated with varying concentrations of Gn, Se, or Gn-Se combination with or 1% Triton X-100 as a positive control. After a 24–48-hour incubation, 10 μL of MTS reagent was added to each well of the 96-well plate. The plates were then incubated at 37°C and 5% CO2 for 4 hours, and the absorbance was read at 490 nm using a plate reader. Percentage inhibition was calculated using the following formula:
where Ab c is the absorbance of the control sample and Ab s is the absorbance of the treated sample.
Treatment-induced apoptosis
Apoptosis is characterized by morphological changes within the cells such as fragmentation of nuclear chromatin and cell shrinkage. These changes can be detected by exposing the cells to certain dyes that when taken up by the cells cause fluorescence, which indicates the level of cell death. Apoptosis induction was determined using the Vybrant® apoptosis assay kit (Molecular Probes, Division of Invitrogen, Eugene, OR, USA). This assay uses two dyes: propidium iodide and YO-PRO®-1. The green fluorescent dye YO-PRO-1 can enter early apoptotic cells, whereas the red fluorescent dye propidium iodide cannot. Therefore, use of both dyes provides a sensitive indication of apoptosis. After staining cells with both dyes, live cells show little or no fluorescence, early apoptotic cells will show green fluorescence, and dead cells (membrane-compromised cells) will show both red and green fluorescence, which could be detected by fluorescence microscopy or flow cytometry. In brief, the cells were cultured to achieve confluence and then were treated as previously described with Gn0–60, Se0–20, Gn-Se combination, or topotecan (50 μM) as a positive inducer of apoptosis. After a 24–48-hour incubation, cells were harvested and washed with cold phosphate-buffered saline, and cell density was adjusted to 1 × 106 cells/mL in phosphate-buffered saline. One microliter of YO-PRO-1 stock solution (Component A) was added to each well, followed by 1 μL of propidium iodide stock solution (Component B), to each 1 mL of cell suspension. The cells were incubated on ice for 20–30 minutes and examined with fluorescence microscopy.
Treatment-induced caspase expression in prostate cells
Caspase proteases have been implicated in the molecular pathways of apoptosis induction in a number of cancer cells. In this study the cells were cultured in triplicates and treated as described previously. The involvement of caspase-3 in the molecular pathway of treatment- induced apoptosis was determined by using a caspase-3 colorimetric activity assay. The assay involves the caspase-mediated cleavage of a labeled peptide substrate acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA):
The p-nitroanilide (pNA) product can be quantified using a spectrophotometer or plate reader at 405 nm. This gives a direct indication of caspase-3 activity. For caspase-3 determination, the cells were treated with varying concentrations of Gn, Se, Gn-Se, or topotecan (50 μM) for the positive control, as previously described. The cells were counted and collected by centrifugation at 10,000 g for 5 minutes, after which 2–4 × 106 cells were resuspended in 200 μL of lysis buffer, incubated on ice for 20 minutes, and centrifuged for 5 minutes at 10,000 g, and the supernatant collected. The BCA protein assay kit (Pierce, Rockford, IL, USA) was used to determine the protein concentration of the samples. Thirty micrograms of protein lysate from each sample was then assayed for caspase activity. Percentage increase in caspase-3 activity was calculated by comparing the absorbance values of the treated samples to that of the control sample.
Treatment-induced MMP-2 expression in the cells
MMP-2 expression in cancer cells has been associated with active invasion and metastasis of many human cancer cells. In the present study, the influence of Gn, Se, and Gn-Se combination on MMP-2 expression level in LNCaP and PC3 cells was investigated. In brief, LNCaP and PC3 cells were cultured and treated as previously above. The cells were cultured for 24 and 48 hours (three identical treatment set-ups). At each interval, the growth medium was removed and further centrifuged (1,000 g, 5 minutes, 4°C), and the supernatant was removed for MMP-2 determination or stored at −35°C until tested/assayed for MMP-2 expression, using a human enzyme-linked immunosorbent immunoassay specific for MMP-2, following the protocol of the manufacturer (Oncogene Research). The absorbance was read at the given wavelength on a Multiskan® enzyme-linked immunosorbent assay plate reader (Thermo Scientific, Waltham, MA, USA). A dose–response graph was drawn by plotting the concentration against the MMP-2 values (in nmol/mL). The experiment was repeated two or three times with the same treatment doses or adjusted dose ranges to reassess the inhibitory activity of the treatments on MMP-2 expression in the cells.
Statistical analysis
The experiments were performed in duplicates to confirm similar results. Significance of the differences in mean values was determined using Student's t test and considering P < .05 to be statistically significant.
Results
Treatment-induced growth inhibition in prostate cells
The cells were incubated with Gn or Se in single and combination treatments at varying cytotoxic dose ranges for 24 and 48 hours, respectively. The results obtained in both MTS and ATP assays indicated that inhibition of cell growth and proliferation was time- and dose-dependent in both PC3 and LNCaP cells (PC3 results shown in Figs. 1 and 2). The Se-treated cells showed significantly higher growth inhibition than the Gn-exposed cells (P < .05; Fig. 2). In all the treatment groups, there were no significant differences in response to treatment between PC3 and LNCaP cells (P < .05), implying that chemosensitivity to treatment was not influenced by hormonal status of the cancer cells. The combination treatment induced the greatest growth inhibition than either of the single treatments (P < .05) in both cells; there were no significant differences in chemosensitivity of the two cell types to Se-Gn combination treatment (P > .05). The pattern of cell response to the treatments was identical in both LNCaP and PC3 cells; hence no graph is shown for LNCaP. In all the treatment groups, growth inhibition was increased concurrently with increasing treatment dosages (Figs. 1 and 2). Neither of the single treatments (Gn or Se) nor the Gn-Se combination had any significant inhibitory effect in normal prostate epithelial NPtEC cells (Fig. 1C), although at higher concentrations (Gn60-Se20), treatment induced some cytotoxicity in the normal cells. The IC50 in LNCaP and PC3 cells at 48 hours of incubation was determined from the data obtained and was found to be about 50–60 μM, 45–50 μM, and 30–40 μM, respectively, with Gn, Se, and Se-Gn combination treatment. The data obtained from the ATP analysis was complementary (similar) to the MTS results, thus confirming the accuracy/validity of the data.
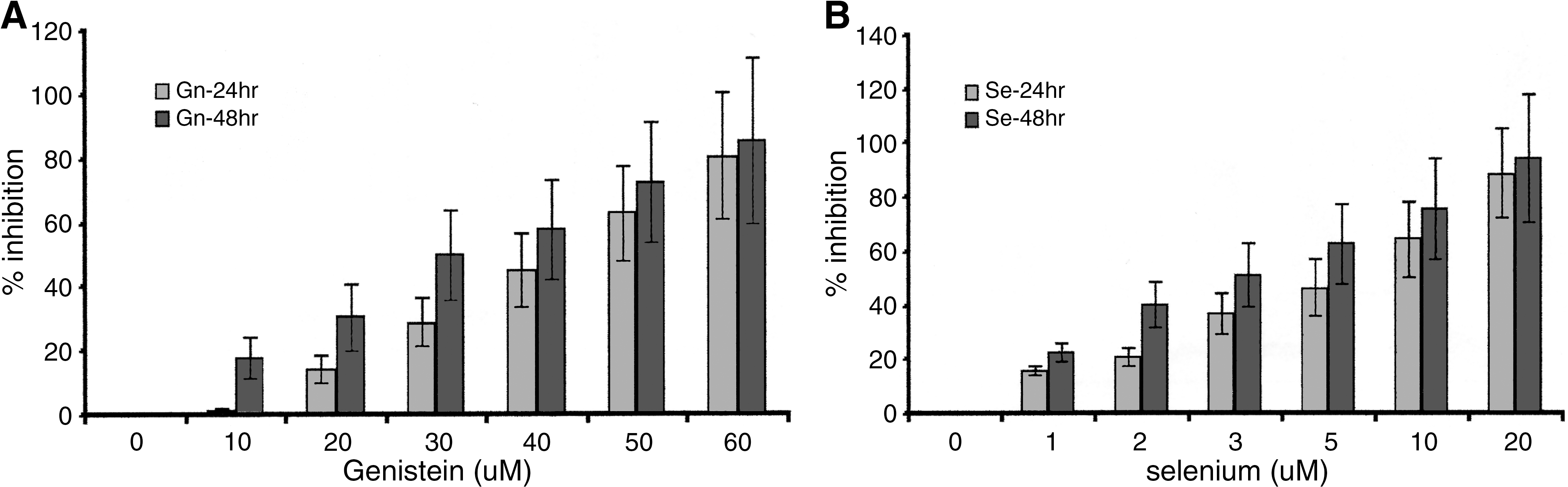
Treatment-induced growth inhibition of PC3 cells. Cells (PC3 and LNCaP [data not shown]) were treated with varying concentrations of Gn, Se, or Gn-Se combination as described in Materials and Methods, and percentage inhibition (shown relative to control untreated cells) was determined using the ATP assay. A dose- and time-dependent result was observed in both (


Treatment-induced growth inhibition of PC3 cells. Cells (PC3 and LNCaP [data not shown]) were treated with varying concentrations of Gn, Se, or Gn-Se combination as described in Materials and Methods, and percentage inhibition was determined using the ATP assay. (
Treatment-induced apoptosis in prostate cells
Treatment-induced apoptosis and percentage apoptosis in PC3 and LNCaP cells were investigated, using the fluorescent dyes YO-PRO-1 and propidium iodide stain according the manufacturer's instructions. The data indicated significant dose- and time-dependent treatment-induced apoptosis in both LNCaP and PC3 cells in all the treatment groups, with no significant differences between LNCaP and PC3 cells. The results also revealed that at identical dose levels, the percentage of apoptosis was significantly higher in the combination-treated (Gn-Se) cells than in cells given either of the single treatments (Gn or Se) (P < .05). This implies a significant therapeutic potential of the combination treatment over either Gn or Se treatment alone. Figure 3 summarizes the data for the treatment-induced apoptosis in PC3 cells. A similar response pattern was observed/obtained for LNCaP cells (data not shown).
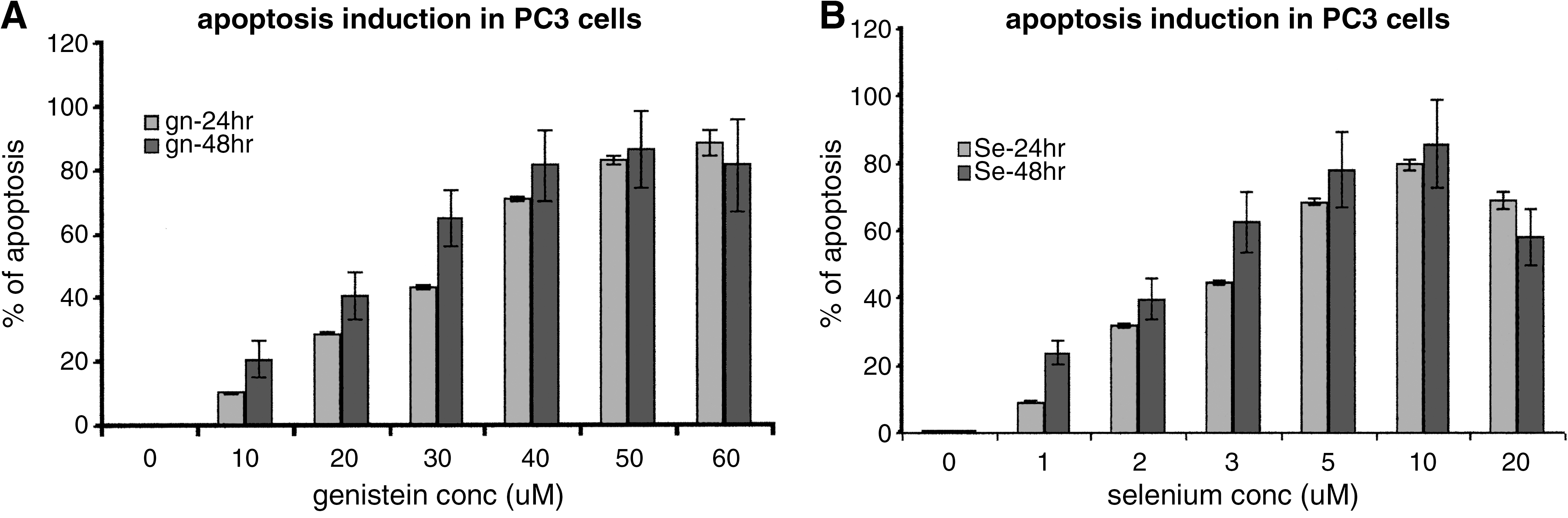
Treatment-induced apoptosis in PC3 cells. Cells (PC3 and LNCaP [data not shown]) were treated with varying concentrations of (

Caspase protease analysis in treated cells
PC3 and LNCaP cells were treated as described above, and post-treatment apoptosis induction was investigated using the caspase-3 colorimetric assay, according to the manufacturer's protocol. The experiments were performed in the presence and absence of a caspase-3 inhibitor. The results showed that there was a significant difference in caspase-3 activity among the three treatment groups (Gn, Se, and Gn-Se). In all cases, the data indicated that caspase-3 induction increased in a time- and dose-dependent manner, as summarized for PC3 cells in Figure 4. The results were similar in both PC3 and LNCaP cells, with no significant difference in caspase-3 activity between the two types of cells (P < .05). Caspase-3 activity was highest in the combination-treated cells relative to the single treatment groups (Gn or Se) (P < .05) (Fig. 4). These results correlate with the data obtained in the apoptosis assay and indicate that Gn-induced cell death involves the activation of the caspase-3 signaling pathway. The results also indicate/reveal that although the caspase protease appears to be the major molecular/signaling pathway by which apoptosis was induced in all the treatment groups, there could be an alternate non-caspase signaling pathway, which appears to be of less significance.
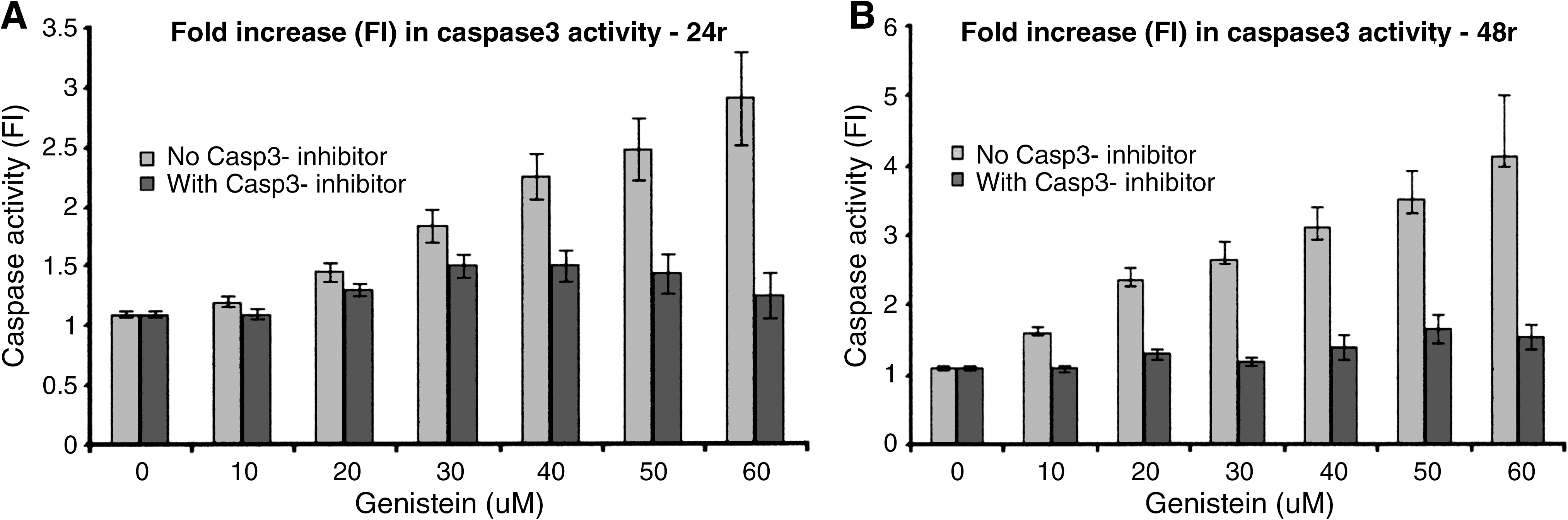
Effect of treatment on caspase-3 (Casp3) activity in PC3 cells. Cells (PC3 and LNCaP [data not shown]) were treated with varying concentrations of (
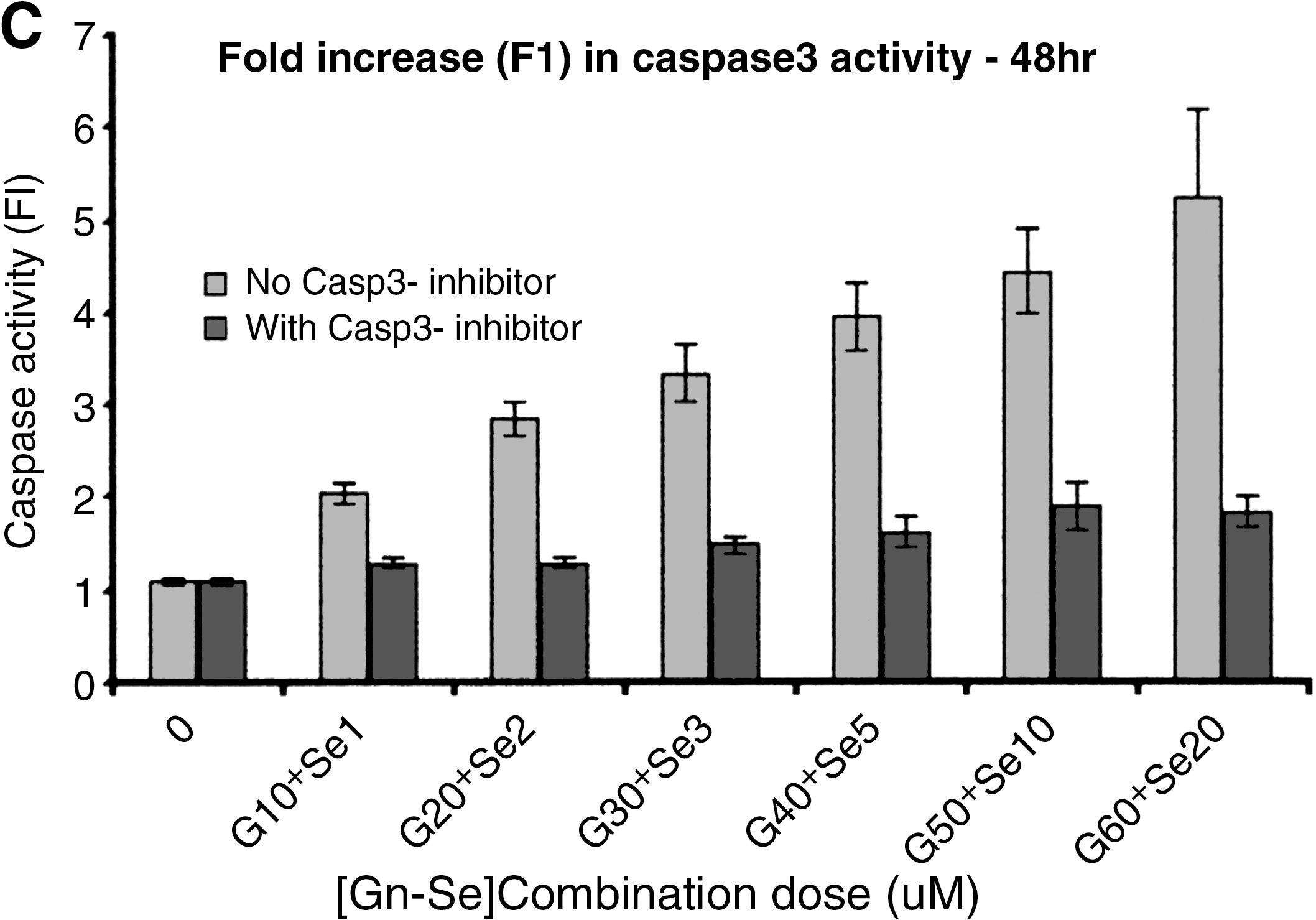
MMP-2 expression levels in treated cells
The absorbance of the sample is directly proportional to the concentration of MMP-2. The data showed a dose- and time-dependent effect of the single and combination treatments on MMP-2 expression levels in both PC3 and LNCaP cells. MMP-2 expression levels in Gn-, Se-, and Gn-Se combination-treated cells decreased progressively with increasing doses. The absolute activity of MMP-2 decreased significantly more in Gn-Se combination-treated cells than in Gn- or Se-treated cells, with significant differences in the decline of expression levels (P < .05) among the treatment groups and with the greatest decline (least expression levels of MMP-2) in the Gn-Se combination-treated cells (Gn > diethylstilbestrol > Gn-diethylstilbestrol). There was a significant negative correlation among the treatment (Gn, Se, and Gn-Se) concentrations and MMP-2 expression levels in both PC3 and LNCaP. This correlation peaked in both PC3 and LNCaP cells treated with the Gn-Se combination: PC3, r = –0.81, P = .031; LNCaP, r = –0.85, P = .04. However, the expression levels of MMP-2 between treated PC3 and LNCaP cells were not significantly different at any dose level (P > .05), implying that hormonal status did not influence MMP-2 expression.
Discussion
At the forefront of chemotherapeutic investigations is the search for treatment regimens with the least cytotoxicity concurrent with minimal side effects in the patient. The focus includes a search into phytochemicals with known potential anticancer properties. This study was done to primarily investigate the potential chemotherapeutic efficacy (growth inhibition) of Gn-Se combination on androgen-insensitive PC3 and androgen-sensitive LNCaP human prostate cancer cells and to determine the potential mechanism of action of the combined treatment. The results indicated that Gn, Se, and Gn-Se combination significantly inhibited growth and proliferation of both prostate cancer cells with no significant difference in cytotoxicity between the two cell lines. Both the MTS and ATP assays revealed a dose- and time-dependent growth inhibitory effect in LNCaP and PC3 cells by all the treatment groups (single and combination). These results were partially in conformity with the findings in previous studies that had reported Gn-induced growth inhibition in prostate and breast cancer cells via apoptosis. 11,19,31 –35 The present data revealed that Gn-Se combination induced significantly greater growth inhibition of both PC3 and LNCaP cells than the Gn and Se single treatments and with no regard to hormonal status. Other studies have shown that Gn-induced apoptosis in prostate cancer cells is independent of the hormonal status of the cells. 33,35 It should be noted that the human PC3 and LNCaP prostate cancer cells exhibit different hormonal phenotypes: PC3 is androgen-insensitive, and LNCaP is androgen-sensitive. It appears that addition of Se (mineral nutrient) to the Gn (phytoestrogenic nutrient) altered/increased the sensitivity of both cells to the Gn isoflavone, which was particularly obvious at higher combination doses. In other studies, Azrak et al. 36 reported the efficacy of Se (methylselenol-cysteine) in increasing the therapeutic index of irinotecan against cancer. The structural composition of the two nutrients, Gn and Se, could partly explain the increased cytotoxicity/efficacy manifested in the combination-treated PC3 and LNCaP cancer cells. It is not certain at this point in the investigation why the chemosensitivity of the two cells with different hormonal status is not significantly different. However, other studies have shown that Gn-induced apoptosis in prostate cancer cells is independent of the hormonal status of the cells. 33,35
Our results indicate that the growth inhibition/antitumor effect of Gn, Se, and/or Gn-Se combination was mainly a result of apoptosis induction in the cells. At similar treatment concentrations, percentage apoptosis was significantly highest in the combination-treated cells than in the Gn- or Se-treated cells. This observation is of therapeutic significance with regard to treatment-induced cytotoxicity in the cells. A time- and dose-dependent induction of apoptosis by Gn and Se has been previously observed in some cancer cells. 32,33,35 However, to our knowledge, the additive effect of Se in the Gn-Se combination in PC3 and LNCaP prostate cancer cells has not been previously reported. Caspase protease signaling/expression was implicated in the apoptosis-induction pathway in both cell types in all the treatment groups, implying that the Gn-Se signaling pathway in apoptosis induction may be independent of hormonal status of the prostate cancer cells. However, the fact that blocking of caspase activity (presence of caspase inhibitors) did not completely abrogate apoptosis in either cell type implies a possible alternate non-caspase pathway for growth inhibition via treatment-induced apoptosis in all the treatment groups. In previous studies we observed that the Gn-induced apoptosis cascade in hormone-dependent LNCaP and hormone-independent PC3 cells was due to activation of caspase-3 and up-regulation of Bax protein in both cells. 11,33,37,38
The overall data obtained in the present study revealed a dose- and time-dependent decrease in MMP-2 expression levels in PC3 and LNCaP cells in all the treatment groups. The greatest depression in MMP-2 expression was seen in the combination-treated cells. Gn-induced decrease in MMP-2 expression levels in cancer cells has been reported previously. 39 MMP-2 expression in cancer cells has been associated with active invasion and metastasis of many human cancer cells. This indicates the potential chemopreventive significance of Gn-Se combination treatment as observed in the present studies, especially if this could be extrapolated into in vivo studies in PC3/LNCaP animal models. A synergistic effect of Gn-Se combination on apoptosis induction in prostate tumors has been recently reported. 14
In conclusion, Gn-Se combination induces the highest cytotoxicity in both LNCaP and PC3 cells independent of hormonal status, relative to single Gn and Se treatment alone. Treatment did not induce significant growth inhibition in normal prostate epithelial cells after a 48-hour treatment. Growth inhibition was induced by apoptotic cell death in all the treatment groups, and the apoptotic cascades induced by the treatments are largely caspase dependent, with evidence of some alternative non-caspase signaling pathway(s). The latter was evidenced by the induction of some degree of apoptosis even in the presence of caspase inhibitors. These results further suggest that Gn-Se combination may have potential chemopreventive value against both hormone-independent and hormone-dependent prostate tumors through induction of apoptosis. Further research is in progress to explore the potential chemopreventive efficacy and clinical significance of Gn-Se combination in animal models of PC3 and LNCaP cells and to determine if Gn-Se combination would be a suitable/useful adjuvant to conventional chemo- and/or radiotherapy.
Footnotes
Acknowledgments
The authors acknowledge the technical staff in our research laboratory for their assistance. Florida Atlantic University is acknowledged for providing the facilities, and Rambaugh-Goodwin Cancer Research Institute at Plantation City, FL, USA, is also acknowledged for allowing the use of its facilities.
Author Disclosure Statement
The authors declare that there are no existing competing financial interests with respect to this research. J.K.-D. conceived the idea, designed the experiment, and contributed to the execution of the experiment, analysis of the data, and writing of the manuscript. K.M. contributed to the culturing and bioassays, and reviewing of the manuscript draft. A.H. contributed a reagent, design of the project, some calculations, and helpful discussions of the results. V.H. contributed to the culturing, writing, and proofreading the drafted manuscript for submission. M.J. contributed to the culturing, data analysis, and proofreading of the draft.