
Abstract
The underlying causes of denervation of the neuromuscular junction and eventual motor neuron death in amyotrophic lateral sclerosis (ALS) have not been resolved. The superoxide dismutase 1 (SOD1)G93A mutant mouse is a frequently used animal model of ALS. We hypothesized that resveratrol (RSV), a polyphenolic molecule that enhances mammalian NAD+-dependent SIRT1 deacetylases and may increase life span, would improve motor function and survival in the SOD1 mouse model via modulation of p53 acetylation. Data were collected for mean survival times, neuromuscular performance on the ROTOR-ROD™ (San Diego Instruments, San Diego, CA, USA), body weight, and p53 acetylation. Mean survival times were not statistically different (P = .23) between control and experimental (RSV-fed) groups (mean ± SD, control [n = 11] 138 ± 6 days vs. experimental [n = 10] 135 ± 8 days). Performance was not significantly different between groups at time points corresponding to 50%, 80%, and 90% mean life span (P = .46), nor did RSV treatment attenuate body weight loss. Thus although manipulation of SIRT1 deacetylase activity has effects at the protein level in healthy aging organisms, we conclude that RSV treatment does not lead to functional improvement or increased longevity in a mouse model of ALS. We speculate that RSV-mediated modulation of p53 acetylation is either incapable of increasing or insufficient to increase motor performance and longevity in this model of ALS.
Introduction
R
Materials and Methods
Animals
All animal experiments were approved by the Wake Forest University School of Medicine Animal Care and Use Committee. SOD1G93A mutant mice were obtained from Jackson Laboratory (Bar Harbor, ME, USA). Once the experimental diet was introduced, all mice were caged individually to monitor daily intake. Body weights of mice were obtained several times weekly. End stage was defined as the time at which mice had lost a minimum of 10% of their 11-week body weight and were unable to right themselves within 30 seconds. 10
Diet
Administration of a soft control diet (n = 11, eight females) and soft RSV diet (n = 10, seven females) was initiated at postnatal day 47, prior to overt symptom onset. 11 RSV diet was prepared as described. 7 In brief, 625 mg of RSV (Orchid Pharmaceuticals, Chennai, India) was mixed to homogeneity into 10 kg of control diet, such that a 25-g mouse consuming 10 g of diet daily ingested 25 mg/kg RSV. The diets were stored frozen and thawed before use to maintain bioavailability. 12 In order to better mimic the conditions in which RSV was found to have significant effects in healthy mice, 7 RSV was administered via diet (rather than other means), at the higher of two doses described.
ROTOR-ROD
Motor performance was assessed by the ROTOR-ROD as described. 13 Animals began familiarization to the apparatus on postnatal days 36–38. Familiarization continued once per week until p70, when animals were tested three times per week. At each test, mice were given three trials to complete 120 seconds on the apparatus that gradually accelerated to a terminal speed of 11.5 rpm. The longest time of the three trials was recorded. Performance testing was terminated when mice could no longer maintain balance on a stationary rod.
Isolation of nuclear protein fraction
At end stage, mice were humanely sacrificed. Spinal cord tissue was quickly harvested from the lumbar vertebral column and homogenized in a glass Dounce mortar-pestle homogenizer containing 0.5 mL of cold homogenization buffer plus 1:20 NaF, 1:1,000 SO4 3−, and 1:100 protease inhibitor solution (Sigma, St. Louis, MO, USA). ll further processing occurred at 4°C. Nuclei were isolated by a 10-minute centrifugation at 500 g. Lysis buffer (250 μL), containing 1:20 NaF, 1:1,000 SO4 3−, and protease inhibitor, was then added to the pellet. The tube was vortex-mixed to disrupt nuclear membranes, allowing fractionation of nuclear protein 14 following further high-speed centrifugation.
Western immunoblotting
Protein concentration was estimated using the BCA protein assay kit and NanoDrop software (Thermo Scientific, Rockford, IL, USA). An equal amount (50 μg) of protein from spinal cord extracts, positive control, and negative control was added to sample buffer such that the final volume was 35 μL. Positive control (p53, catalog number MS-104-PCL) and negative control protein (albumin, catalog number 23209) were obtained from Thermo Scientific (Waltham, MA, USA). Samples were boiled for 5 minutes. Thirty-microliter samples were loaded onto a 10-well 10% Tris-HCl sodium dodecyl sulfate-polyacrylamide gel (Bio-Rad, Hercules, CA, USA). After electrophoresis, proteins were transferred to a polyvinylidene difluoride Immobilon™ membrane (Millipore, Billerica, MA, USA). Acetylated p53 (ac-p53) was detected using a previously validated 15 anti-acetyl-p53 (Upstate, Billerica) at 1:1,000 dilution, in conjunction with an enhanced chemiluminescent detection system (Thermo Scientific) per the manufacturer's instructions. Following the p53 probe, antibody was stripped, and the membrane was reprobed for β-actin (Calbiochem, La Jolla, CA, USA), an internal control for protein loading. Intensity of p53 bands was normalized to the intensity of β-actin bands using MultiGuage version 3.0 (Fujifilm, Norcross, GA, USA) software.
Statistical analysis
Using GraphPad Prism software (version 4.03, GraphPad Software, Inc., La Jolla) Student's t test was used to compare the RSV-treated versus control mice sample means for body weight, life span, and normalized p53 band pixel density, and analysis of variance was used to compare ROTOR-ROD performance of groups over time. Significance was accepted at P ≤ .05.
Results
Body weight and life span
To measure effects of RSV treatment on body weight in SOD1G93A mice we monitored body weights of experimental and control mice throughout the experiment. The body weights of experimental (n = 10) and control (n = 11) mice were not statistically different at postnatal day 71, 108, or 122. These time points correspond to 50%, 80%, and 90% mean life span. Furthermore, mean life spans were not significantly different (experimental, 135 ± 8 days; control, 138 ± 6 days), nor was probability of survival, expressed using a Kaplan-Meier survival curve (Fig. 1A). No mice in either group died prematurely, suggesting that RSV (25 mg/kg) is not toxic to mice at this dose. A pilot study showed that, as the disease progressed, mice in both groups had difficulty raising their heads to consume food and water. We therefore provided food and water in shallow dishes, which were placed at the bottom of the cage. By postnatal day 90 it was obvious that not all animals were consuming their entire daily diet; however, this observation was true for both groups.
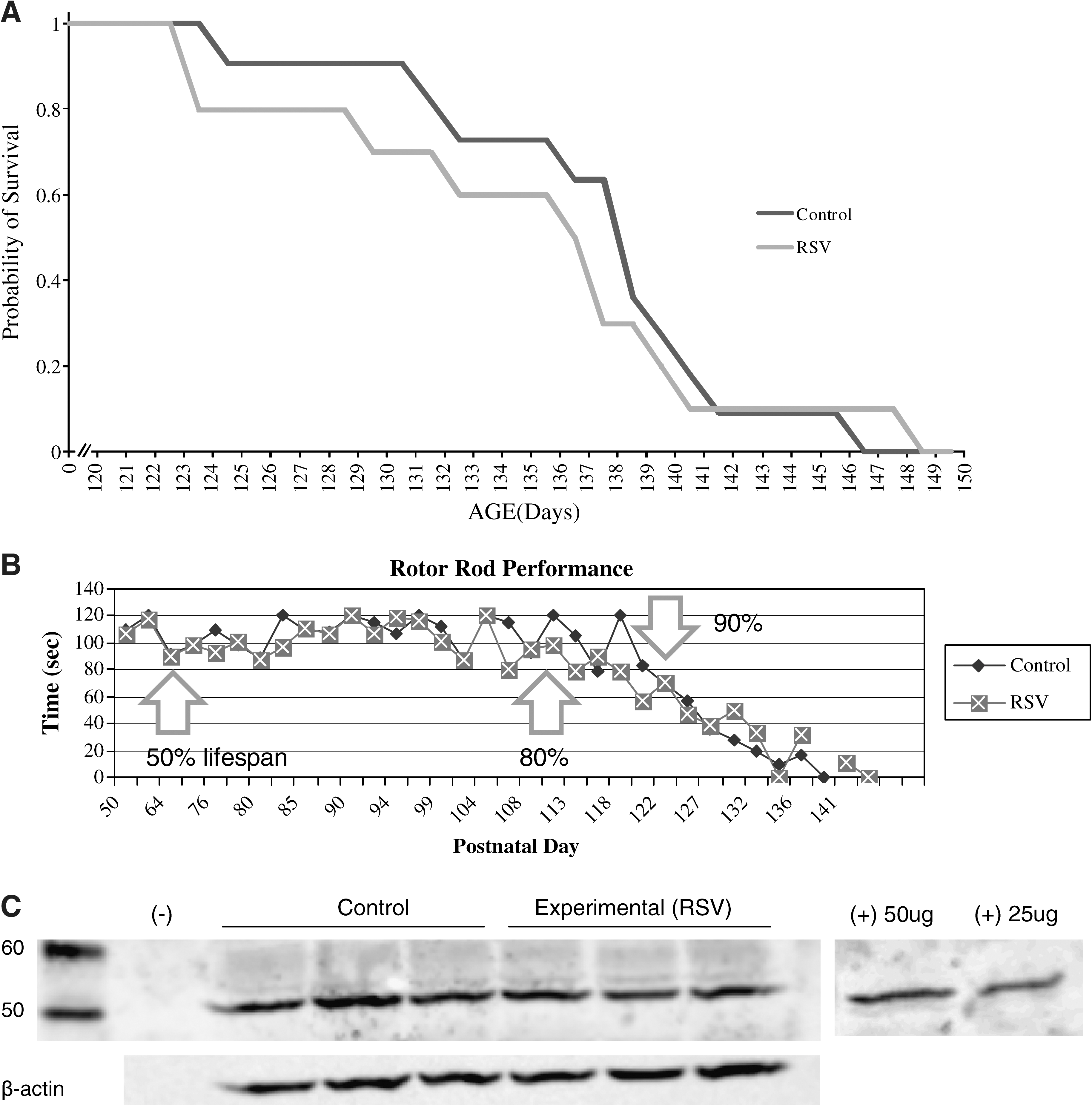
(
ROTOR-ROD performance
To measure effects of RSV treatment on motor performance in SOD1G93A mice, we monitored latency to fall time between experimental and control mice. ROTOR-ROD performance is a commonly used test of the neuromuscular system in mouse models of disease. 13,16 There was no difference in motor performance or functional decline between the RSV and control diet groups (Fig. 1B).
Immunoblot analyses
To measure effects of RSV treatment on the acetylation status of p53, spinal cord lysates from experimental and control mice were subjected to western blot analysis. Following normalization to protein load, intensities of bands for ac-p53 were not significantly different between groups (Fig. 1C).
Discussion
The main finding from the present study indicates that chronic oral administration of RSV, at a dose approximating that previously studied 7 in healthy mice (25 mg/kg/day), does not mitigate the progressive neurodegenerative effects of ALS as measured by the ROTOR-ROD test, body weight, or life span in an ALS mouse model. Furthermore, daily dietary intake of RSV at this dose does not appear to alter the acetylation of the tumor suppressor factor, p53, as measured by western blot analysis. Reasons for the lack of beneficial effect of RSV in the present study are not clear but may include posttranslational modifications of p53, 17 insufficient RSV dose, or bioavailability of the compound. To address these concerns, pharmacokinetic studies of various dosages of RSV might be worthy of further study; however, the validity of even the most widely accepted methods for determining RSV and SIRT1 activity in vivo has been questioned. 18 Thus, the ostensibly simple notion of measuring dietary absorption of RSV is limited by current assays. As a surrogate assay for our hypothesis testing, guided by review of the literature, we performed western blots to evaluate the effects of RSV on ac-p53. At the single dose studied, not one of our outcome measures indicated a biological effect of dietary RSV.
Although the dosage of RSV used in our study was approximately the same as the dosage previously described as having physiological effects, 7 an important difference between the studies is the duration of treatment. Because of the premature deaths of SOD1 mutant mice, the RSV treatment only lasted approximately 90 days. Treatment of healthy mice with RSV for over a year produced beneficial effects, mimicking caloric restriction. 7 Future studies that investigate the chronic effects of various dosages of RSV over the truncated life span of SOD1 mice might yield different results. Importantly, previous research has suggested that the SIRT1 pathway plays a role in neurodegenerative diseases. 1
Previous reports 8,9 have considered the effects of caloric restriction in SOD1 mice. However, both of these reports found that death was accelerated. If caloric restriction and RSV similarly affect SIRT1, it would be reasonable to speculate that RSV might also accelerate death. In our study, daily dietary administration of 25 mg/kg RSV did not accelerate death; however, RSV did not prolong life or improve neuromuscular performance in SOD1 mice.
It is possible that RSV acts in vivo to prolong life span through molecular pathways that are independent of p53. Evidence 7 indicates that healthy rodents fed a hypercaloric diet do not experience metabolic disease associated with obesity and aging if they are concurrently fed 22.4 mg/kg RSV daily. Thus, it may again be reasonable to speculate that life span is modulated by caloric restriction and RSV via insulin-dependent pathways. These pathways may include insulin-like growth factor-1, 5′-AMP-activated protein kinase, peroxisome proliferator-activated receptor γ coactivator-1α, 7 and heat shock proteins. 9,13 Future studies may consider whether these pathways, and the potential clinical benefits of modulating these pathways, are mediated at the nucleus or at the mitochondria. 19,20
Conclusion
Chronic daily administration of RSV (25 mg/kg) does not mitigate progression of neurodegeneration in the SOD1G93A mouse model of ALS. This finding is consistent with literature 8,9 showing that caloric restriction is not protective in the SOD1G93A mouse model of ALS.
Footnotes
Acknowledgments
We thank the R.W. Oppenheim laboratory for helpful discussions and pilot studies. This study was funded by laboratory startup funds for M.K.C.
Author Disclosure Statement
All authors declare that no competing financial interests exist.