
Abstract
In vitro and in vivo studies have recently reported significant chemopreventive effects of green tea-derived polyphenols in different diseases. However, it remains unclear how such effects could be triggered. In order to elucidate the effects of epicatechin gallate (ECG) in C6 cells, both by itself and against H2O2-induced genotoxicity, measurements of DNA strand breaks and chromosome loss were performed. DNA damage was measured by comet and micronucleus assays. The present study shows for the first time how ECG, the major green tea-derived polyphenol, is able to exert dose-dependent genoprotective effects in an H2O2-induced toxicity model of C6 astroglial cells. We demonstrate that doses of ECG in a range from 0.1 to 1 μM were able to completely prevent H2O2-induced genotoxicity in vitro. In contrast, considerably higher concentrations of ECG (10 μM) were able to reverse previous positive effects in a dose- and time-dependent manner. The same results were confirmed by both comet (F 3,9 = 336,148; P < .001) and micronucleus (F 3,9 = 23,228; P < .001) methods. Together, our data show ECG as a dose-dependent genoprotective compound in C6 astroglial cells. This indicates that small doses of polyphenols included in our diet could have beneficial effects on neural cells, contributing to prevention of oxidative stress-associated brain pathologies. In addition, our data highlight the importance of strictly modulating doses and/or consumption of antioxidant-fortified foods or additional supplements containing such beneficial molecules.
Introduction
S
Green tea, from Camellia sinensis, is obtained as a result of minimal oxidation during the production process. It is widely consumed throughout the world and has received a great deal of attention since tea polyphenols were shown to be strong antioxidants. 2
Green tea-derived polyphenols, such as catechins, may be responsible for reactive oxygen species quenching. 3 The chemopreventive effects of green tea have been attributed to biochemical activities induced by its polyphenolic constituents such as (-)-epicatechin gallate (ECG), (-)-epicatechin, (-)-epigallocatechin 3-gallate, and (-)-epigallocatechin. 4,5
The capability of flavonoids to act as an antioxidant depends on their molecular structure, the position of their hydroxyl groups, and other possible chemical substitutions in their organic structure. 6 The relative antioxidant activity of green tea-derived catechins is (-)-epigallocatechin 3-gallate = ECG > (-)-epigallocatechin > (-)-epicatechin. 7 Some studies have demonstrated that excessive production of free radicals and reactive oxygen species, such as hydrogen peroxide (H2O2), is indeed harmful, promotes aging, and can strongly influence the development of many diseases such as cancer 8 as well as cardiovascular and neurodegenerative disorders. 9 –11 Tea polyphenols are retained in the brain and may exert neuroprotective effects whenever their consumption is controlled. 12 In fact, prolonged green tea ingestion is able to significantly protect hippocampal proteins and lipids against oxidative damage. 13
Previous works have demonstrated that intracellular reactive oxygen species and reactive nitrogen species such as superoxide anion, hydroxyl radicals, H2O2, lipid peroxyl radicals, nitric oxide, and peroxynitrite can lead to destruction of different cellular components, including lipids, proteins, and DNA. 6,14 However, cells can still survive thanks to antioxidant-dependent repair mechanisms. 15 Characteristic properties of polyphenols such as penetration in brain tissue as well as their antioxidant and iron-chelating capabilities may make such compounds putative natural drugs for the treatment of oxidative-related neurodegenerative diseases. 16
Glial cells, particularly astrocytes, are known to extensively interact with neuronal elements in the brain, influencing their activity and exerting a prominent role in both protection and repair of nervous tissue after damage. 17 The C6 cell line was originally derived from rat tumors induced by N-nitrosomethylurea, 18 and it is widely used as an astrocyte-like cell line. 19 –25
The main aims of our study were to elucidate the effects of ECG in C6 cells, both by itself and against H2O2-induced genotoxicity by measuring DNA strand breaks and chromosome loss.
Experimental Procedures
Materials
Ethidium bromide, material for cell culture, and ECG were purchased from Sigma (St. Louis, MO, USA). 4′,6′-Diamidino-2-phenylindole was from Calbiochem (La Jolla, CA, USA). Dulbecco's modified Eagle's medium was purchased from Gibco BRL (Carlsbad, CA, USA), and fetal bovine serum was purchased from Cultilab (Campinas, SP, Brazil). All other chemicals were obtained from regular commercial suppliers.
Cell culture
C6 astroglial cells were cultured as previously described. 25 In our preparations, more than 95% of the cells exhibited positive immunoreactivity to glial fibrillary acidic protein. Late passages of cells (100 passages, minimum) were seeded in flasks and cultured in Dulbecco's modified Eagle's medium (pH 7.4) supplemented with 5% fetal bovine serum, 2.5 mg/mL amphotericin B (Fungizone®, Bristol-Myers Squibb, Princeton, NJ, USA), and 100 U/L gentamicin. Exponentially growing cells were incubated for 1, 6, 12, and 24 hours at 37°C in an atmosphere of 5% CO2/95% air in Dulbecco's modified Eagle's medium (pH 7.4) without serum in the absence or presence of ECG (0.1, 1, or 10 μM). The concentrations of ECG used in these experiments were obtained from previous determinations. 26
H2O2 treatment
In order to investigate the genoprotective effects of ECG against H2O2-induced oxidative stress, cells were preincubated with different concentrations of ECG (0.1, 1, and 10 μM) for 1 hour at 37°C in an atmosphere of 5% CO2/95% air in Dulbecco's modified Eagle's medium (pH 7.4) without serum. After this time, the medium was maintained, and 1 mM H2O2 was added. 27 Cells were incubated in the same conditions for an additional 30 minutes.
Nuclear morphology assay
C6 cells were cultured on circular glass coverslips and treated with or without 1 and 10 μM ECG for 1, 6, 12, or 24 hours. Cells were fixed for 20 minutes with 4% paraformaldehyde in phosphate-buffered saline, stained with 0.2 μg/mL 4′,6′-diamidino-2-phenylindole for 1 hour, and visualized under a fluorescent microscope (Nikon [Tokyo, Japan] inverted microscope using a TE-FM epi-fluorescence accessory). Apoptotic cells were morphologically identified by nuclear shrinkage and chromatin condensation and/or fragmentation.
Cytokinesis-block micronucleus assay
Micronuclei are DNA-containing structures that result from chromosomal loss during mitosis. They represent a subgroup of all chromosomal aberrations. This makes the micronucleus frequency test a widely accepted method for investigating both in vitro and in vivo genotoxicity in human biomonitoring studies. 28 The cytokinesis-block micronucleus technique was performed as previously described 29,30 with minor modifications. After treatment, cells were incubated with 2 μg/mL cytochalasin B for 24 hours, fixed with chilled methanol/glacial acetic acid (3:1 vol/vol) for 5 minutes, and stained with Giemsa. Each data point represents the mean of eight independent experiments. In each experiment, 1,000 binucleated cells/sample were analyzed.
Comet assay
After different treatments as described above, C6 cells were detached by incubating in the presence of trypsin/EDTA (0.05%). During trypsinization, cells were carefully manipulated to avoid mechanical stress. Comet assay (single-cell gel electrophoresis) was performed as previously described. 27 In brief, slides were prepared by mixing 30 μL of C6 cell suspension with 70 μL of low-melting -point agarose (0.75%). Following electrophoresis, slides were incubated with 5 μg/mL ethidium bromide and left in the dark for 20 minutes to stain the DNA. Images of 100 randomly selected nuclei (50 nuclei from two replicated slides) were analyzed for each treatment. Nuclei were scored visually for comet tail size based on an arbitrary scale of 0–4, i.e., ranging from no damage to extensive damage of DNA. Therefore, the damage index scale could range from 0 (all nuclei without tail, 100 cells × 0) to 400 (all nuclei with maximally elongated tails, 100 cells × 4). Slides were viewed on a Nikon inverted microscope using a TE-FM epifluorescence accessory, and images were transferred to a computer with a digital camera (Sound Vision Inc., Wayland, MA, USA).
Statistical analysis
To verify the dose- and time course-dependent effect of ECG, we used one-way analysis of variance for repeated measures, followed by a post hoc analysis (Tukey's test). ECG effects × different doses of H2O2 were analyzed statistically by two-way analysis of variance followed by a post hoc analysis (Tukey's test). Data are mean ± SEM values. Values with P < .05 were considered to be significant. All analyses were carried out in a PC-compatible computer using the Statistical Package for Social Sciences (SPSS) software (SPSS Inc., Chicago, IL, USA).
Results
Effect of ECG on chromosome loss in C6 cells
Micronucleus frequency (Table 1) was increased by ECG, compared to their respective control values (F 3,48 = 262,381; P < .001). The analysis also indicate a significant interaction between concentration and time (F 3,9 = 23,228; P < .001).
Cells were incubated with epicatechin gallate (ECG) (0.1, 1.0, and 10 μM) for 1, 6, 12, or 24 hours. Data are mean ± SEM values for micronucleus frequency among 1,000 binucleated cells, from eight independent experiments performed in duplicate. To verify the main effect of time course and different doses of ECG, repeated-measures one-way analysis of variance was used, followed by post hoc analysis with Tukey's test.
Values that do not share a common letter differ significantly at P < .001.
Effect of ECG on nuclear morphology and DNA strand breaks in C6 cells
To assess the direct effect of ECG on DNA integrity, cells were incubated in the presence of 0.1, 1, and 10 μM ECG for 1, 6, 12, or 24 hours. As shown in Figure 1, ECG induced DNA damage (F 3,64 = 1946,747; P < .001). This effect was concentration and time dependent (F 3,9 = 336,148; P < .001). Only 2% of cells exposed to 10 μM ECG after 24 hours presented nuclear fragmentation by 4′,6′-diamidino-2-phenylindole staining assay (data not shown).

In vitro time- and dose-dependent effects of ECG on DNA damage. C6 astroglial cells were exposed to different concentrations of ECG (0.1, 1, and 10 μM) for different time intervals (1–24 hours). The extent of damage to DNA was determined by the comet assay. The index of DNA damage was calculated as described in Materials and Methods. Data are mean ± SEM values of 16 experimental determinations performed in duplicates. To verify the dose- and time course-dependent effect of ECG, repeated-measures analysis of variance was performed, followed by post hoc analysis with Tukey's test. abcdefColumns that do not share a common letter differ significantly at P < .001.
Genoprotective effects of ECG against H2O2-induced DNA damage in C6 cells
To investigate the effect of ECG on DNA damage induced by H2O2, cells were preincubated with different concentrations of ECG (0.1, 1, and 10 μM) for 1 hour. After this time, 1 mM H2O2 was added and maintained for 30 minutes (Fig. 2A). Such an assay was performed at the 1-hour time point taking into account that at this time ECG by itself had minimal influence on DNA integrity (Fig. 1). ECG-induced genoprotective effects after treatment are shown in Figure 2B. The index of DNA damage observed when cells were incubated in the presence of H2O2 in the absence of ECG was 42.5 ± 3.9. In this context, ECG at 0.1 and 1.0 μM (20.0 ± 1.3 and 22.0 ± 1.5, respectively) (F = 47,529; P < .001) was able to significantly prevent the H2O2-induced genotoxicity in vitro.
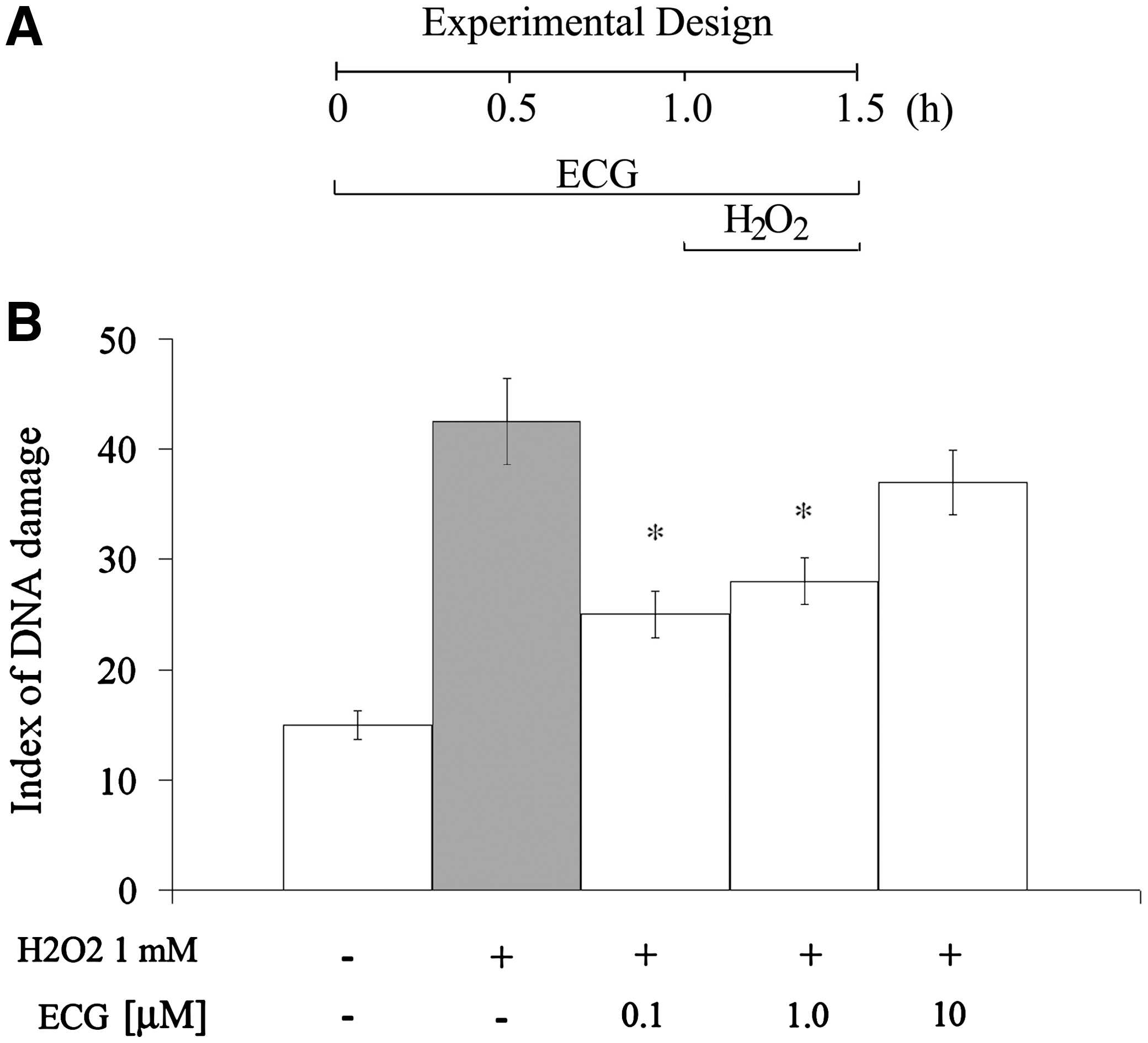
Protective effect of ECG against H2O2-induced DNA damage in C6 astroglial cells. Cells were preincubated for 1 hour in the presence of ECG (0.1, 1, and 10 μM) before exposure to H2O2. Culture medium was maintained, and then 0.1 mM H2O2 was added. Cells were incubated in the same conditions for 30 minutes longer. The extent of damage to DNA was determined by the comet assay. The index of DNA damage was calculated as described in Materials and Methods. Data are mean ± SEM values. Differences were statistically analyzed by two-way analysis of variance followed by Tukey's test. *Significant differences from control values (P < .001).
Discussion
It is very well documented that several redox-active compounds could have a dualistic effect, either beneficial or toxic, depending on the concentration used. 25,27,31,32 The ability of polyphenols to scavenge reactive oxygen species depends on their chemical structures. 33 In addition, antioxidant/pro-oxidant activity of different redox-active compounds, such as polyphenols, largely depends on the levels consumed within the diet and may potentially cause DNA damage, 34 probably triggered by direct binding of polyphenol to DNA. 35
As previously reported by our group, 26 10 μM ECG induces morphological alterations in the C6 astroglial cell line (process-bearing cells) with a small increase in propidium iodide incorporation after 24 hours of incubation (up to 4%). In the present work, we observed that, at least up to 24 hours, ECG did not induce any indicative alteration in nuclear morphology related to apoptotic process (data not shown). DNA strand breaks in individual cells result from events such as direct scission of the DNA backbone.
In the present in vitro experimental model (1 hour in the presence of ECG, before exposure to H2O2 for 30 minutes), ECG was not cytotoxic to C6 cells. However, it is important to mention that index values below 30 are obtained from less prominent DNA strand breaks. In such context, the first 6 hours of ECG incubation resulted in mild DNA damage. Thereafter, ECG clearly induced a time- and dose-dependent genotoxicity.
The degree of chromosome loss, analyzed by the micronucleus frequency test (Table 1), reflects the capacity of the cells to resist oxidative stress and repair single-strand breaks. 34 As ECG by itself induced DNA damage at higher doses, we decided to investigate if this genotoxic effect could impair DNA repair. Actually, micronuclei frequency values from ECG exposure were significantly higher than control values. The highest dose of ECG induced an increase (about 60%) in all incubation time points (Fig. 1). It indicates that ECG genotoxicity at higher doses affects the repair system, probably via a pro-oxidant effect. 32
In order to investigate possible genoprotective effects of ECG against DNA damage induced by oxidant conditions, we used an experimental procedure previously established in our group. 27 In this model, cells were preincubated with antioxidant for 1 hour before H2O2-induced insult. H2O2 is particularly attractive as an oxidant model because its cellular actions and fate are well understood. It readily crosses the cellular membrane and gives rise to highly reactive hydroxyl radicals, which have the ability to react with different macromolecules, including DNA, proteins, and lipids, and to ultimately damage a cell. 36
Following the observation that the 1-hour time point of ECG exposure did not induce genotoxic effects by itself in C6 cells, we decided to keep the same time point for ECG preincubation, just before the H2O2 pulse at 30 minutes, which is known to be sufficient and enough to induce genotoxic properties. 27 We found that H2O2-induced genotoxicity was significantly prevented by 0.1 and 1 μM ECG, suggesting that, at these concentrations, ECG plays a beneficial effect probably due to its antioxidant properties. The reaction between ECG and hydroxyl radicals is particularly important for preventing oxidative injury because hydroxyl radicals have been shown to be highly responsible for several pathogeneses related to a wide range of diseases.
Despite the fact that our in vitro data demonstrated a significant astroglial genoprotection upon ECG treatment, some limitations should be under consideration regarding the difficulty of correlating in vitro experiments with in vivo treatments. Plasma ECG concentrations in rats range from 0.2 to 135 nM after 7.3 mg/kg ECG administration via the tail vein. 37 The pharmacokinetics of catechins in rat brain fetuses after pregnant ingestion of green tea extract could result in a ECG range from 10 to 80 pmol/g. 38 Moreover, under in vitro experiments, drug concentrations need to be usually higher than chronic in vivo treatments in order to mimic long-term physiological cell activities and their functions. 27
Conclusions
Our results clearly demonstrated that ECG can affect DNA in our cell model, but, most importantly, it is able to protect against H2O2-induced DNA damage, showing a statistically significant dose-dependent genoprotective effect. This indicates that small doses of polyphenols included in our daily diet could play genoprotective effects in neural cells and, therefore, could ameliorate oxidative stress-associated brain pathologies. Nevertheless, the dualistic genotoxic effect exerted by high doses of ECG suggests that caution is recommended when large quantities of antioxidants and/or supplements in foods are consumed.
Footnotes
Acknowledgments
We would like to thank Dr. Richard Rodnight for critical reading and helpful discussion of the manuscript. This work was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, FINEP/Rede IBN grant 01.06.0842-00, and the National Institute of Science and Technology for Excitotoxicity and Neuroprotection.
Author Disclosure Statement
The authors declare that no competing financial interests exist.