
Abstract
Red ginger (Zingiber officinale var. Rubra) has been prescribed as an analgesic for arthritis pain in Indonesian traditional medicine. The surface color of the rhizome is purple because of the anthocyanidins in its peel. We prepared 40% ethanolic extract from dried red ginger (red ginger extract [RGE]) and evaluated its anti-inflammatory activity using acute and chronic inflammation models. In an acetic acid-induced mouse writhing model, RGE (10–100 mg/kg) suppressed both the frequency of writhing and the increase in permeability of abdominal capillaries. On the other hand, continuous treatment with RGE (10 mg/kg) significantly (P < .05) suppressed footpad edema in a rat adjuvant arthritis model. To clarify the anti-inflammatory mechanism of RGE, we examined the effect on prostaglandin (PG) and nitric oxide (NO) production from mouse leukemic monocytes (RAW264 cells) stimulated by lipopolysaccharide. RGE (3 and 10 μg/mL) significantly (P < .05) suppressed PGE2 production, while it also suppressed NO production at 100 μg/mL. After bioassay-guided separation of RGE, we found that [6]-shogaol and gingerdiols suppressed NO production. Red dye fractions presumed to be proanthocyanidins also suppressed NO production at 100 μg/mL. Consequently, we found a potent suppressive effect of RGE on acute and chronic inflammation, and inhibition of macrophage activation seems to be involved in this anti-inflammatory effect. [6]-Shogaol, gingerdiols, and proanthocyanidins were identified as constituents that inhibited NO production.
Introduction
G
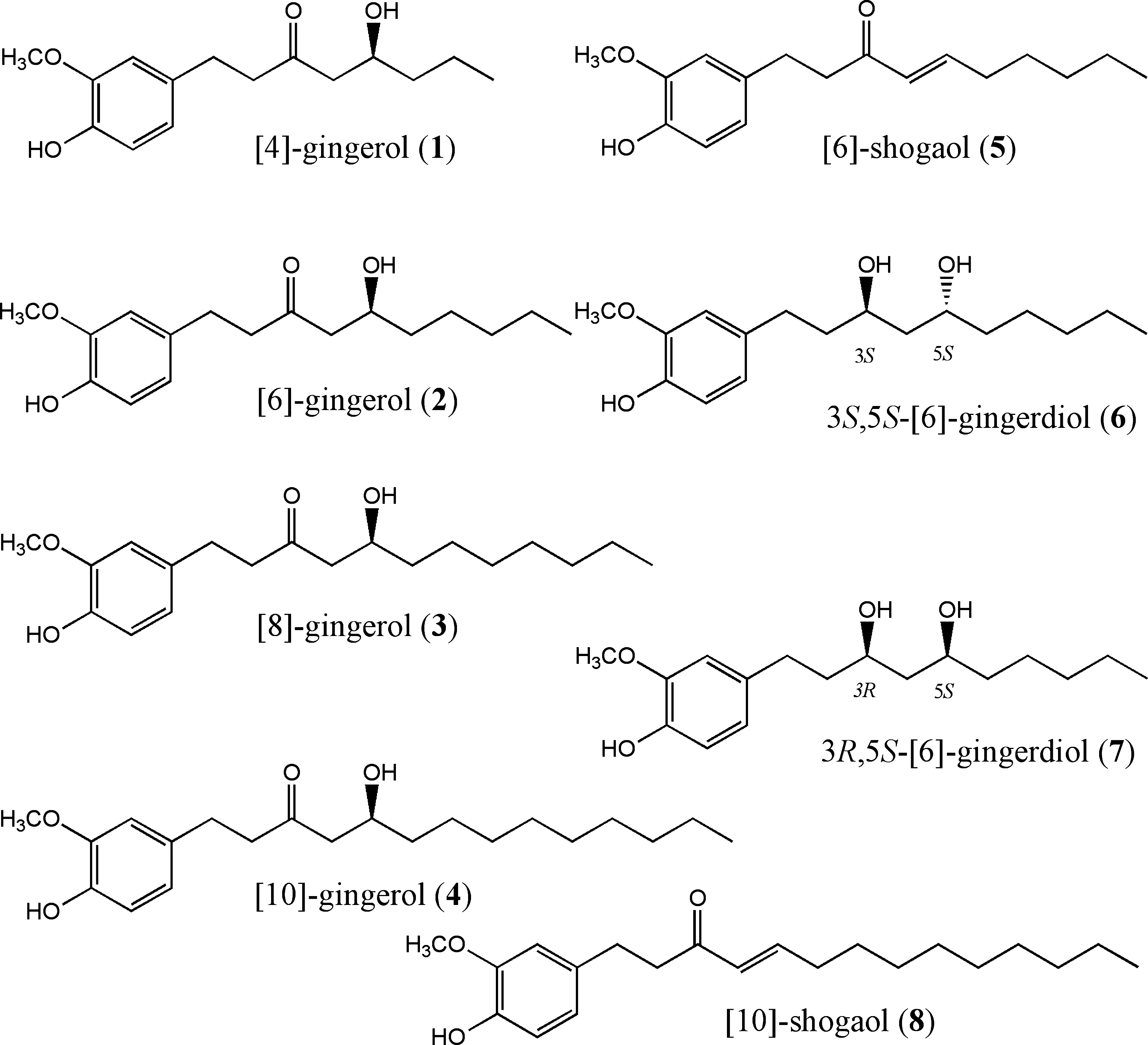
Structures of compounds isolated from red ginger.
Materials And Methods
Animals and cells
Male ddY mice (5 weeks old) were obtained from Japan SLC, Inc. (Shizuoka, Japan). The mice were housed in an air-conditioned room (23 ± 3°C, 50 ± 10% relative humidity) and were fed a standard nonpurified diet (CE-2, Clea Japan, Inc., Shizuoka) and tap water ad libitum. The experiments were performed in accordance with the 1987 Guidelines for Animal Experimentation of the Japan Association for Laboratory Animal Science. Male Sprague-Dawley rats (8 weeks old) were purchased from Orient Bio Inc. (Seongnam, Korea). The rats were housed in an air-conditioned room (23 ± 3°C, 50 ± 10% relative humidity) and fed a standard experimental diet (PMI Nutrition International, Brentwood, MO, USA) and sterilized tap water ad libitum. All rats used in this study were cared for in accordance with the principles outlined by the National Institutes of Health in the Guide for the Care and Use of Laboratory Animals. The study described in this protocol does not unnecessarily duplicate previous experiments (Association for Assessment and Accreditation of Laboratory Animal Care International accredited, 1998).
Mouse leukemic monocytes, RAW264 cells (number RCB0535), were obtained from Riken Bio Resource Center (Tsukuba, Japan).
Preparation of RGE
Sliced and dried red ginger (10 kg), cultivated on Java Island in Indonesia, was powdered and defatted with n-hexane (30 L, 40°C) followed by extraction (80°C, 2 hours) with 40% ethanol (50 L), and the solvent was evaporated. The extract was powdered by spray-drying. The yield of RGE was 7.4%. The contents of principal gingerol derivatives determined by high-performance liquid chromatography were as follows: [4]-gingerol (
Isolation and identification of RGE constituents
RGE (100 g) was suspended in H2O (500 mL) and partitioned with ethyl acetate (500 mL × 3). The solvent was evaporated to obtain the ethyl acetate portion (20.0 g). The ethyl acetate portion (1 g) was subjected to reversed-phase high-performance liquid chromatography on a 250- × 10-mm (inner diameter) octadecylsilanized silica gel column (GL Science, Tokyo, Japan) in 80% methanol at 4 mL/minute to obtain [4]-gingerol (
Reagents
Acetic acid and dimethyl sulfoxide were obtained from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). Pontamine sky blue 6BX was purchased from Alfa Aesar (Ward Hill, MA, USA). Freund's incomplete adjuvant and desiccated Mycobacterium butyricum were purchased from Difco BRL (Kansas City, MO, USA). Indomethacin, Eagle's minimum essential medium, fetal calf serum, nonessential amino acids solution, penicillin and streptomycin mixture solution, and LPS (from Escherichia coli, serotype O127:B8) were obtained from Sigma-Aldrich (St. Louis, MO, USA). The prostaglandin (PG) E2 enzyme immunoassay kit monoclonal (Cayman Chemical Co.) was purchased from GE Healthcare (Pittsburgh, PA, USA). The Griess reagent kit (NO2/NO3 assay kit-CII) and methylthiazole tetrazolium (MTT) were purchased from Dojindo Laboratories (Kumamoto, Japan).
Acetic acid–induced inflammation in mice
According to the method of Matsuda et al., 12 RGE suspended in 5% acacia solution was given orally to fasted (20-hour) mice (6 weeks old). Pontamine sky blue solution (2%) was injected (10 mL/kg) from the tail vein 55 minutes later, and then acetic acid (1%) was injected (10 mL/kg) into the abdominal cavity 5 minutes after the injection. The writhing frequency was counted for 15 minutes from 5 minutes after the acetic acid injection. The mouse was sacrificed under ether anesthesia, and the inside of the abdominal cavity containing pontamine sky blue was flushed gently with saline (8 mL). Leaked dye solution was filled with saline to a volume of 10 mL, and the absorbance was measured at 590 nm.
Adjuvant arthritis in rats
Freund's incomplete adjuvant (0.1 mL) containing desiccated M. butyricum (1 mg) was injected subcutaneously into the right hind paw of rats (8 weeks old). RGE suspended in 0.5% sodium carboxymethyl cellulose solution was given daily after immunization, and the increase in edema in the right hind paw was measured by a digital plethysmometer (LE 7500, Panlab SL, Barcelona, Spain).
Determination of PGE2 and NO production from RAW264 cells
According to the method of Yun et al., 13 RAW264 cells were suspended in Eagle's minimum essential medium containing 0.1 mM nonessential amino acids, 10% fetal calf serum, penicillin (100 units/mL), and streptomycin (100 μg/mL) at a concentration of 1 × 106 cells/mL. The cell suspension (200 μL) was seeded in a 48-well culture plate and cultured for 24 hours. The cells were rinsed with fetal calf serum-free medium (200 μL), and fetal calf serum-free medium (170 μL) was added to each well. Ten microliters of LPS (200 μg/mL) solution and 20 μL of sample were added, and the cells were cultured for 20 hours. The supernatant was collected to measure PGE2 and NO using the PGE2 enzyme immunoassay kit monoclonal and Griess reagent kit.
Statistics
The in vivo results were expressed as means and SE values, and the cell study results were given as mean and SD values. Significance of the differences was examined by one-way analysis of variance or two-way analysis of variance followed by Dunnett's test. Differences of P < .05 were considered significant.
Results And Discussion
The anti-inflammatory activity of RGE on acute inflammation was examined using a mouse writhing model induced by intraperitoneal injection of acetic acid. Figure 2 shows the increase in writhing behavior and dye leakage into the abdominal cavity caused by enhancement of capillary permeability. Oral treatment with RGE (10 –100 mg/kg) suppressed the writhing frequency and dye leakage in a dose-dependent manner. RGE was found to exhibit analgesic and anti-inflammatory effects against acute inflammation by oral treatment. Young et al.
5
reported that [6]-gingerol (
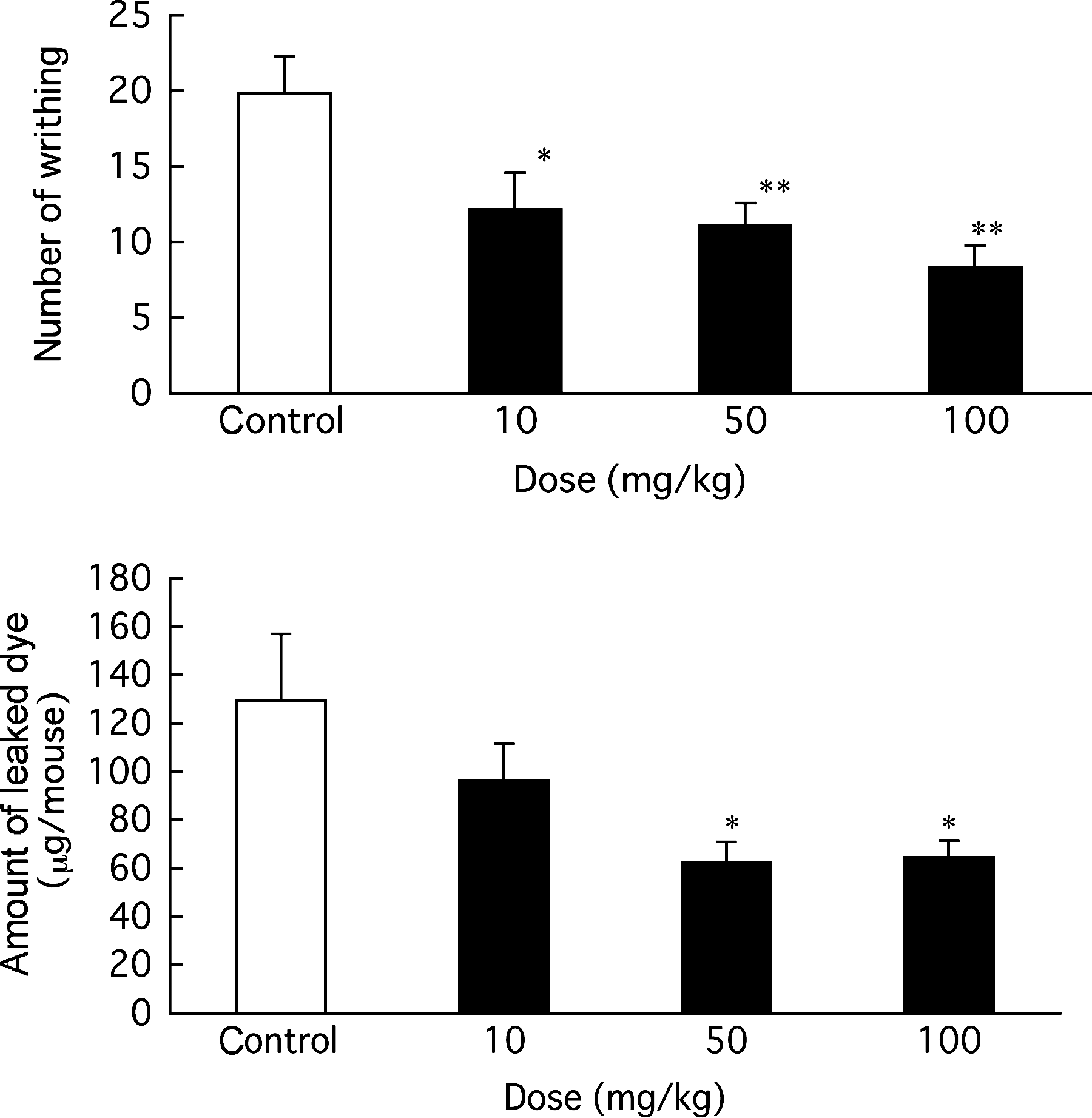
Effect of RGE on acetic acid-induced writhing and inflammation in mice. RGE was given orally to fasted mice, and pontamine sky blue solution was injected (200 mg/kg) intravenously 55 minutes later. Then, acetic acid (100 mg/kg) was injected intraperitoneally 5 minutes after the injection. The writhing frequency was counted for 15 minutes from 5 minutes after the acetic acid injection. The mouse was sacrificed, and the leaked dye in the abdominal cavity was washed with saline. The absorbance of collected dye solution was measured at 590 nm. Data are mean ± SE values of 12 animals. Significance of differences was examined by one-way analysis of variance followed by Dunnett's test. Asterisks denote significant differences from the control: *P < .05, **P < .01.
Subsequently, we examined the effect of RGE on chronic inflammation using a rat adjuvant arthritis model. As shown in Figure 3, continuous oral treatment with RGE (10 mg/kg/day) for 13 days significantly suppressed foot paw edema. Sharma et al.
14
reported that daily oral treatment with ginger oil (33 mg/kg) prepared from the dried rhizome of Z. officinale Roscoe by steam distillation suppressed adjuvant arthritis in rats. However, the oil is not thought to contain [6]-gingerol (

Effect of RGE on adjuvant arthritis in rats. Adjuvant arthritis in the rat foot was induced by subcutaneous injection of a mixture of Freund's incomplete adjuvant (0.1 mL) and M. butyricum (1 mg) into the hind paw. RGE was given daily after adjuvant injection, and the increase in edema in the hind paw was measured. Data are mean ± SE values of seven animals. Significance of differences was examined by two-way analysis of variance followed by Dunnett's test. Asterisks denote significant differences from the control: *P < .05, **P < .01.
To investigate the inhibitory mechanism of RGE on acute and chronic inflammation, LPS-induced PGE2 and NO productions in RAW264 cells were evaluated. RAW264 cells are macrophage-like cells that are frequently used to evaluate the effect of samples on the release of cytokines and inflammatory mediators induced by inflammatory stimuli.
17,18
RGE (1–10 μg/mL) suppressed PGE2 production in a concentration-dependent manner (Fig. 4). The 50% inhibitory concentration (IC50) value was 10 μg/mL. The suppressive effects of ginger constituents on PGE2 production have been reported for Chinese white ginger and Japanese yellow ginger.
19,20
Inhibition of PGE2 production is thought to be the principal mechanism of the anti-inflammatory action of ginger. In this experiment, we confirmed that red ginger also suppresses LPS-induced PGE2 production in RAW264 cells. Inhibition of COX-2 activity by ginger was reported to be involved in the mechanism for suppression of PGE2 production.
21
Among the constituents of ginger, [8]-paradol, [8]-shogaol, and [8]-gingerol (
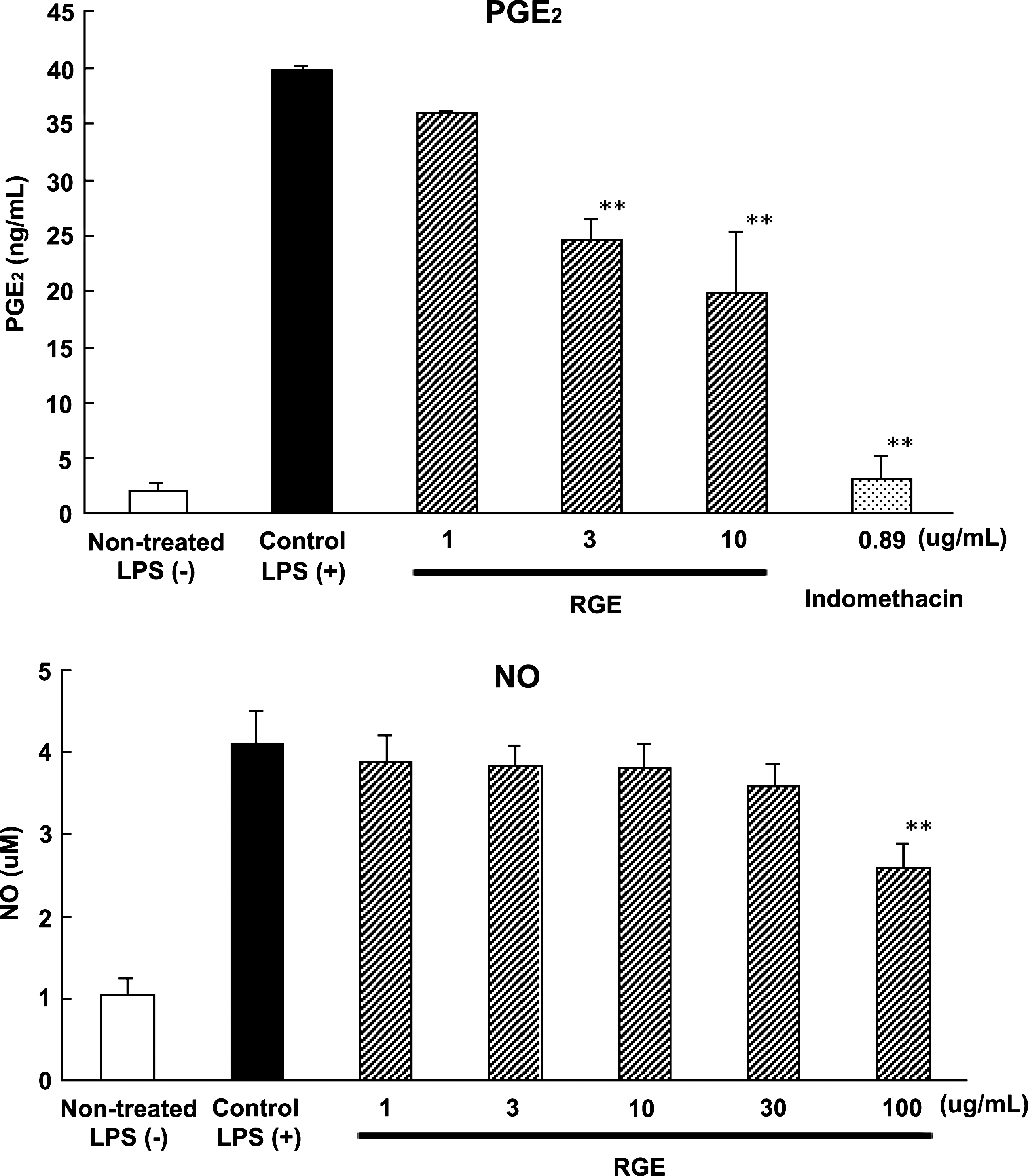
Effect of RGE on PGE2 and NO production in RAW264 cells induced by LPS. RAW264 cells (2 × 105 cells) in 200 μL of medium were precultured for 24 hours. The medium was replaced with fetal calf serum-free medium, and each test sample solution and LPS (10 μg/mL) were added. The cells were cultured for 20 hours, and the supernatant was collected. Data are mean ± SD values of four to six experiments. Significance of differences was examined by one-way analysis of variance followed by Dunnett's test. Asterisks denote significant differences from the control: **P < .01.
Excessive NO production by inducible NO synthase is closely related to rat adjuvant arthritis.
26,27
Inhibitors of NO synthase were reported to potently suppress adjuvant arthritis,
28
so we examined the effect of RGE on NO production in LPS-stimulated RAW264 cells. High concentrations of RGE significantly suppressed LPS-stimulated NO production in RAW264 cells (Fig. 4). MTT assay revealed no cytotoxicity up to this high concentration. Imanishi et al.
29
reported that an extract prepared from common ginger (Z. officinale Roscoe) suppressed NO production in RAW264.7 cells at 100 μg/mL. Our data obtained from red ginger (Z. officinale var. Rubra) were similar to their results. Regarding the suppressive effect of ginger constituents on NO production, only [6]-gingerol (
RAW264 cells (2 × 105 cells in 200 μL of medium) were precultured for 24 hours. The medium was replaced with fetal calf serum-free medium, and each test sample solution and LPS (10 μg/mL) were added. The cells were cultured for 20 hours, and the supernatant was collected. NO concentration represents mean ± SD values from six experiments.
Significance of the differences was examined by one-way analysis of variance followed by Dunnett's test. Asterisks denote significant differences from the control group [LPS (+)]: **P < .01.
We also investigated RDF containing proanthocyanidins. RDF suppressed NO production by 35.4%. With purified fractions (RDF-1– 4), all fractions exhibited suppressive effects at 100 μg/mL. Among them, RDF-3 and - 4 potently suppressed NO production. Recently, several reports relevant to the inhibitory effect of food-derived proanthocyanidins on NO production have been reported, including those from longan flowers, 34 persimmon peel, 35 and grape seeds. 36 Inhibition of inducible NO synthase is thought to be involved in the mechanism. 37 Therefore, it is possible for RDF to inhibit NO production by inhibition of NO synthase.
In conclusion, RGE was found to suppress acute and chronic inflammation. Inhibition of PGE2 by gingerol derivatives is considered to be involved in the major anti-inflammatory mechanism. Additionally, inhibition of NO production by various gingerol derivatives and proanthocyanidins appears to be a minor contributor to the anti-inflammatory effect of RGE.
Footnotes
Author Disclosure Statement
H.S., S.-J.S., and J.T. are employees of Oryza Oil & Fat Chemical Co., Ltd. A.S. is an employee of Hamuri Co., Ltd. No competing financial interests exist for J.-W.S., N.K., S.T., Y.K., and N.M.