
Abstract
Although the immunomodulatory effects of many herbs have been extensively studied, research related to possible immunomodulatory effects of various spices is relatively scarce. Here, the potential immunomodulatory effects of black pepper and cardamom are investigated. Our data show that black pepper and cardamom aqueous extracts significantly enhance splenocyte proliferation in a dose-dependent, synergistic fashion. Enzyme-linked immunosorbent assay experiments reveal that black pepper and cardamom significantly enhance and suppress, respectively, T helper (Th)1 cytokine release by splenocytes. Conversely, Th2 cytokine release by splenocytes is significantly suppressed and enhanced by black pepper and cardamom, respectively. Experimental evidence suggests that black pepper and cardamom extracts exert pro-inflammatory and anti-inflammatory roles, respectively. Consistently, nitric oxide production by macrophages is significantly augmented and reduced by black pepper and cardamom, respectively. Remarkably, it is evident that black pepper and cardamom extracts significantly enhance the cytotoxic activity of natural killer cells, indicating their potential anti-cancer effects. Our findings strongly suggest that black pepper and cardamom exert immunomodulatory roles and antitumor activities, and hence they manifest themselves as natural agents that can promote the maintenance of a healthy immune system. We anticipate that black pepper and cardamom constituents can be used as potential therapeutic tools to regulate inflammatory responses and prevent/attenuate carcinogenesis.
Introduction
S
A wide range of natural products isolated from herbs and spices have been shown to possess immunomodulatory effects that can be very beneficial in fighting many diseases. 9 –12 The great potential that some spices possess in terms of preventing and treating various diseases, including cancer, cannot be underestimated. 13 –15 However, relatively little is known about the potential immunomodulatory and anticancer effects of a wide range of commonly used spices, and the molecular mechanisms underlying such effects are either poorly understood or largely unidentified. In this study, we focus on investigating the potential immunomodulatory and anti-cancer effects of black pepper (Piper nigrum) and cardamom (Elettaria cardamomum). We also attempt to shed light on the possible molecular mechanisms by which black pepper and cardamom extracts exert their immunomodulatory and anti-cancer effects.
Many studies have previously demonstrated that black pepper, its extracts, and its major constituents have diverse physiological effects in the gastrointestinal tract, kidney, and liver. 16 Black pepper extracts have also been shown to exert antimicrobial activities. 17 –19 Moreover, some studies have demonstrated that black pepper extracts and its major constituents possess anti-cancer properties in vitro and in vivo. 20 –24 Indeed, the ability of black pepper to alter the metabolic activity of various enzymes has been proposed as the mechanism of action by which black pepper inhibits carcinogenesis. 22,23,25,26
Cardamom was shown to play a wide range of health-promoting roles against various conditions such as constipation, colic, diarrhea, dyspepsia, vomiting, headache, epilepsy, and cardiovascular diseases. 27,28 Recently, cardamom was reported to exhibit spasmogenic, spasmolytic, blood pressure-lowering, vasodilator, diuretic, and sedative activities. 29 Antimicrobial properties of cardamom extracts have been documented. 30 –33 Experimental evidence suggests that cardamom extracts display anti-cancer activities. 26,34 –37 Enzymatic modulation potential 26,34 –37 and anti-inflammatory, antiproliferative, and pro-apoptotic activities 36 have been proposed as mechanisms underlying the anti-cancer properties of cardamom.
Our findings suggest that black pepper and cardamom are potential immunomodulators of splenocyte proliferation and Th1/Th2 cytokine profiles. Furthermore, our findings also suggest that the aqueous extracts of black pepper and cardamom exert pro-inflammatory and anti-inflammatory roles, respectively. Finally, our study also suggests that the previously identified anti-cancer activities of black pepper and cardamom extracts may be mediated via the profound potential of such extracts to provoke the cytotoxic activity of natural killer (NK) cells. Hence, our study provides compelling evidence suggesting that black pepper and cardamom extracts may serve as potential immunoregulators of inflammatory responses, Th1/Th2 immune responses, and carcinogenesis. We anticipate that black pepper and cardamom constituents may serve as natural therapeutic agents to prevent/treat diverse inflammatory conditions and various types of cancer.
Materials and Methods
Mice
Age-matched BALB/c and C57/BL6 mice (6–8 weeks old) were purchased from Jackson Laboratories (Bar Harbor, ME, USA) and were kept on a 12-hour light/dark cycle in the Carleton Animal Care Facility at Dalhousie University, Halifax, NS, Canada. Mice were fed chow diet and were sacrificed by cervical dislocation for splenocyte or macrophage isolation.
Preparation of aqueous extracts of the spices studied
Whole-seed black pepper and cardamom were washed, dried, and ground in liquid nitrogen. After complete evaporation of liquid nitrogen, 10 mL of double distilled H2O was added to the 20 g of ground spice and stirred overnight with magnet stirrer to allow extraction. The crude spice extracts were centrifuged at 10,000 g for 15 minutes at room temperature. Subsequently, the supernatants were harvested and subjected to rotatory evaporation. After complete evaporation, a stock concentration of 20 mg/mL of each extract was prepared, and the extracts were sterilized by filtration using Nalgene filters (Thermo Fisher Scientific, Rochester, NY, USA) (pore size, 0.22 μm).
Reagents and materials
Thioglycollate broth medium was purchased from DIFCO (Detroit, MI, USA). Lipopolysaccharide (LPS) isolated from Escherichia coli O55:B5 and concanavalin A (ConA) were purchased from Sigma-Aldrich (St. Louis, MO, USA). RPMI-1640 medium, fetal bovine serum, penicillin-streptomycin, and
Isolation of splenocytes
Splenocyte isolation was performed as previously described. 38 In brief, spleens were isolated from BALB/c mice, cut into several pieces, and gently crushed. Clumps were further dispersed by passing the suspension through a 19-gauge needle. Subsequently, cell suspension was filtered through a 200-μm mesh nylon screen, and cells were collected by centrifugation. Erythrocytes were lysed using ACK lysis buffer (0.15 M NH4Cl, 1 mM KHCO3, and 0.1 mM disodium EDTA), and splenocytes were finally washed and resuspended in RPMI-1640 medium supplemented with 10% heat-inactivated fetal bovine serum, 1% penicillin-streptomycin, 10 mM HEPES, and 50 μM β-mercaptoethanol. By trypan blue exclusion, cell counting revealed >98% viability.
Peritoneal macrophage isolation and culture
Thioglycollate-elicited peritoneal macrophages were isolated from BALB/c mice as previously described. 39 In brief, mice were injected intraperitoneally with 3 mL of sterile 3% Brewer's thioglycollate broth solution (Sigma-Aldrich). Five days later, mice were sacrificed by cervical dislocation, and peritoneal exudate cells were isolated by peritoneal lavage. Peritoneal exudate cells were obtained by centrifugation and resuspended in ACK lysis buffer for erythrocyte lysis. Subsequently, cells were centrifuged, resuspended, and cultured in RPMI-1640 medium supplemented with 10% heat-inactivated fetal bovine serum, 1% penicillin-streptomycin, 10 mM HEPES, and 50 μM β-mercaptoethanol. By trypan blue exclusion, cell counting revealed >98% viability.
In vitro splenocyte proliferation assay
Splenocyte proliferation was assayed as previously described. 40 In brief, 2 × 105 splenocytes were cultured for 48 and 72 hours in medium supplemented with vehicle, 10 ng/mL LPS, 1 μg/mL ConA, and aqueous extracts of black pepper and cardamom. Subsequently, cultured splenocytes were pulsed with [3H]thymidine (1 μCi per well) for 16 hours before cell harvest. Cells were harvested using a semiautomated multiwell harvester, and cell lysates were transferred onto fiberglass filter paper (Skatron Instruments, Lier, Norway). The dried filter paper was subsequently transferred to a vial containing 1.5 mL of scintillation fluid (Beckman, Fullerton, CA, USA). Incorporation of [3H]thymidine was determined using the 1211 Rackbeta scintillation counter (LKB Wallac, Turku, Finland).
Measurement of nitric oxide (NO) production by macrophages (Griess assay)
NO production by macrophages was assessed by the colorimetric Griess reaction as previously described. 41 In brief, 2 × 105 macrophages were cultured in medium supplemented with vehicle, 10 ng/mL LPS, 2 U/mL IFNγ, a combination of LPS and IFNγ, or aqueous extracts of black pepper and cardamom in the presence or absence of IFNγ and/or LPS for 48 hours. Subsequently, 100 μL of supernatant and serial dilutions of NaNO2 standard solution were placed in 96-well microtiter plates and then mixed with Griess reagent containing 1% sulfanilamide, 0.1% naphthylethylenediamide dihydrochloride, and 2.5% H3PO4. The optical density was measured at 550 nm using an Emax® precision microplate reader (Molecular Devices, Sunnyvale, CA, USA), and the amount of accumulated nitrite in the samples was quantified according to the standard curve.
NK activity assessment by JAM assay
The cytotoxic activity of NK cells was assessed by JAM assay as previously described. 42 In brief, YAC-1 tumor cells were cultured for 4 hours in medium containing 5 μCi/mL [3H]thymidine for labeling. Subsequently, labeled YAC-1 tumor cells were cultured in 96-well V-bottom culture plates in the presence or absence of splenocytes (containing NK cells) isolated from C57/BL6 mice at effector:target ratios (E:T ratios) of 200:1, 100:1, and 50:1. YAC-1 tumor cells cultured in the presence or absence of splenocytes were treated with vehicle or aqueous extracts of black pepper and cardamom. At 4 hours post-incubation, YAC-1 tumor cells were harvested using a semiautomated multiwell harvester, and radioactivity was measured using the 1211 Rackbeta scintillation counter (LKB Wallac). Percentage cytotoxicity was determined as follows: % cytotoxicity = ([vehicle-treated YAC-1 tumor cells − targeted-YAC-1 tumor cells]/vehicle-treated YAC-1 tumor cells) × 100.
Assessment of cytokine secretion by ELISA
For assessment of interleukin (IL)-4, IL-10, and IFNγ release, 2 × 105 splenocytes were treated with vehicle, 10 ng/mL LPS, 1 μg/mL ConA, and aqueous extracts of black pepper and cardamom in the presence or absence of 1 μg/mL ConA for 48 hours. For assessment of IL-6 and tumor necrosis factor-α (TNFα) release, 2 × 105 macrophages were treated with vehicle, 10 ng/mL LPS, 2 U/mL IFNγ, a combination of LPS and IFNγ, and aqueous extracts of black pepper and in the presence and absence of LPS plus IFNγ for 48 hours and 12 hours (IL-6 and TNFα, respectively). Subsequently, supernatants were harvested, and cytokine concentration was determined using BD OptEIA ELISA kits and the Emax precision microplate reader.
Statistical analysis
Data are mean ± SEM values of the indicated number of experiments. Statistical significance was determined using Student's t test for unpaired observations; *P < .05, **P < .01, and ***P < .001 were considered statistically significant.
Results
Assessment of splenocyte proliferation in the presence of aqueous extracts of black pepper and cardamom
We hypothesized that aqueous extracts of black pepper and cardamom can potentially enhance splenocyte proliferation. To test this hypothesis, BALB/c splenocytes were cultured in medium supplemented with vehicle, ConA (T lymphocyte mitogen), LPS (B lymphocyte mitogen), or aqueous extracts of black pepper or cardamom at four doses (1, 10, 50, and 100 μg/mL) for 48 and 72 hours. Subsequently, cultured splenocytes were subjected to an in vitro proliferation assay using [3H]thymidine incorporation to assess the potential modulatory effects of the aqueous extracts on splenocyte proliferation. As shown in Figure 1A and B, the proliferation of splenocytes was enhanced in a dose-dependent manner in the presence of aqueous extracts of black pepper and cardamom at 72 hours. Notably, all doses (except 1 μg/mL) of black pepper aqueous extract led to significant enhancement of splenocyte proliferation, but significant enhancement by of splenocyte proliferation by cardamom was only observed at 100 μg/mL. Noteworthy is that significant enhancement of splenocyte proliferation at 48 hours was only observed with black pepper aqueous extract at 50 and 100 μg/mL (data not shown).
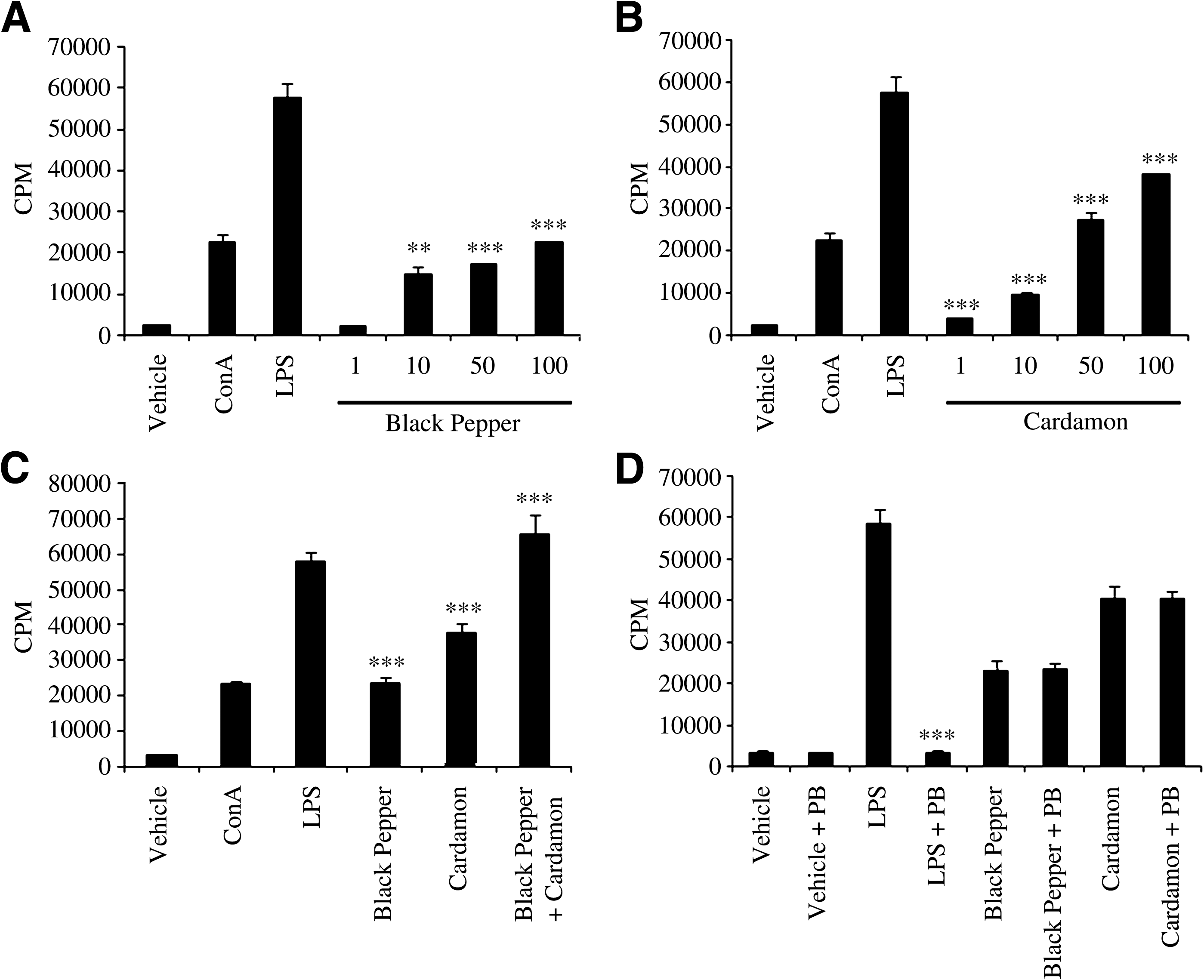
Dose-responsive effect of aqueous extracts of (
Synergistic stimulatory effect of aqueous extracts of black pepper and cardamom on splenocyte proliferation
To determine whether the aqueous extracts of black pepper and cardamom have any synergistic effect on splenocyte proliferation, BALB/c splenocytes were cultured in medium supplemented with 100 μg/mL aqueous extract of black pepper, cardamom, or a combination of both. Vehicle, LPS, and ConA were used as experimental controls. At 72 hours post-incubation, cultured splenocytes were subjected to the in vitro splenocyte proliferation assay. As shown in Figure 1C, splenocyte proliferation was significantly greater in the presence of aqueous extracts of both black pepper and cardamom in combination compared to the effect of each extract separately. Indeed, the enhancement of splenocyte proliferation in the presence of the aqueous extracts of both black pepper and cardamom in combination is comparable to that caused by LPS (P = .23). A significant synergistic effect of the aqueous extracts of black pepper and cardamom on splenocyte proliferation was also observed at lower doses (1, 10, and 50 μg/mL) of the extracts (data not shown), in which the stimulatory action of one extract is enhanced and magnified in the presence of the other extract. Notably, the aqueous extract of cardamom displayed a significantly greater potential to enhance splenocyte proliferation compared to the aqueous extract of black pepper (P = .004) (Fig. 1C). Together, these data suggest that not only do aqueous extracts of black pepper and cardamom significantly enhance the proliferation of splenocytes, but they also interact cooperatively to further augment splenocyte proliferation.
Polymyxin B (PB), which is known to potentially inactivate LPS, was used to determine whether the aqueous extracts of black pepper and cardamom were LPS-contaminated. To this end, BALB/c splenocytes were cultured in medium supplemented with aqueous extracts of black pepper and cardamom at 100 μg/mL in the presence or absence of 1 μg/mL PB and subsequently subjected to the in vitro splenocyte proliferation assay. Clearly, PB significantly reduced the ability of LPS to promote splenocyte proliferation, whereas the ability of aqueous extracts of black pepper or cardamom to enhance splenocyte proliferation was not altered in presence of PB (Fig. 1D). Collectively, these results clearly rule out the possibility that the aqueous extracts of black pepper and cardamom enhance splenocyte proliferation because of possible LPS contamination.
Aqueous extracts of black pepper and cardamom modulate cytokine release by splenocytes and macrophages
To further evaluate the immunomodulatory effects of the aqueous extracts of black pepper and cardamom, the production of IL-4, IL-10, and IFNγ by lymphocytes and the production of IL-6 and TNFα by macrophages were assessed. To assess IL-4, IL-10, and IFNγ secretion, BALB/c splenocytes were cultured in medium supplemented with vehicle, LPS, ConA, and aqueous extracts of black pepper and cardamom (1, 10, 50, and 100 μg/mL) in the presence or absence of ConA. As shown in Figure 2A and C, the aqueous extract of black pepper had no significant effect on IL-4 and IL-10 release by splenocytes at any of the doses compared to vehicle-treated splenocytes. However, dose-dependent inhibition of IL-4 and IL-10 release was observed when splenocytes were treated with the aqueous extract of black pepper in the presence of ConA (Fig. 2A and C). With regard to cardamom, IL-4 and IL-10 release was significantly enhanced when splenocytes were treated with 50 and 100 μg/mL aqueous extract of cardamom compared to vehicle-treated splenocytes (Fig. 2B and D). Likewise, a dose-dependent increase in IL-4 and IL-10 release was observed when splenocytes were treated with aqueous extract of cardamom in the presence of ConA compared to ConA-treated splenocytes (Fig. 2B and D).

Effect of aqueous extracts of (
Noticeably, treatment of splenocytes with the aqueous extract of black pepper was accompanied by a dose-dependent increase in IFNγ release in the presence and absence of ConA compared to ConA-treated and vehicle-treated splenocytes, respectively (Fig. 2E). As shown in Figure 2F, the aqueous extract of cardamom had no significant effect on IFNγ release by splenocytes at any of the doses compared to vehicle-treated splenocytes. However, dose-dependent inhibition of IFNγ release was observed when splenocytes were treated with the aqueous extract of cardamom in the presence of ConA (Fig. 2F).
To assess IL-6 and TNFα secretion, BALB/c macrophages were cultured in medium supplemented with vehicle, LPS, IFNγ, combination of LPS and IFNγ, and aqueous extracts of black pepper and cardamom (1, 10, 50, and 100 μg/mL) in the presence or absence of LPS plus IFNγ. At 48 and 12 hours post-incubation, supernatants were harvested and subjected to ELISA to measure the concentration of IL-6 and TNFα, respectively. As shown in Figure 3A and C, IL-6 and TNFα release by macrophages was significantly enhanced by the aqueous extract of black pepper in the presence and absence of LPS and IFNγ compared to macrophages treated with LPS and IFNγ and vehicle-treated macrophages, respectively. Clearly, the aqueous extract of cardamom alone had no significant effect on IL-6 and TNFα release by macrophages at any of the doses (Fig. 3B and D). However, dose-dependent inhibition of IL-6 and TNFα release was observed when macrophages were treated with the aqueous extract of cardamom in the presence of LPS and IFNγ (Fig. 3B and D). Collectively, these findings suggest that black pepper and cardamom possess immunomodulatory functions with regard to cytokine release profile in splenocytes and macrophages.
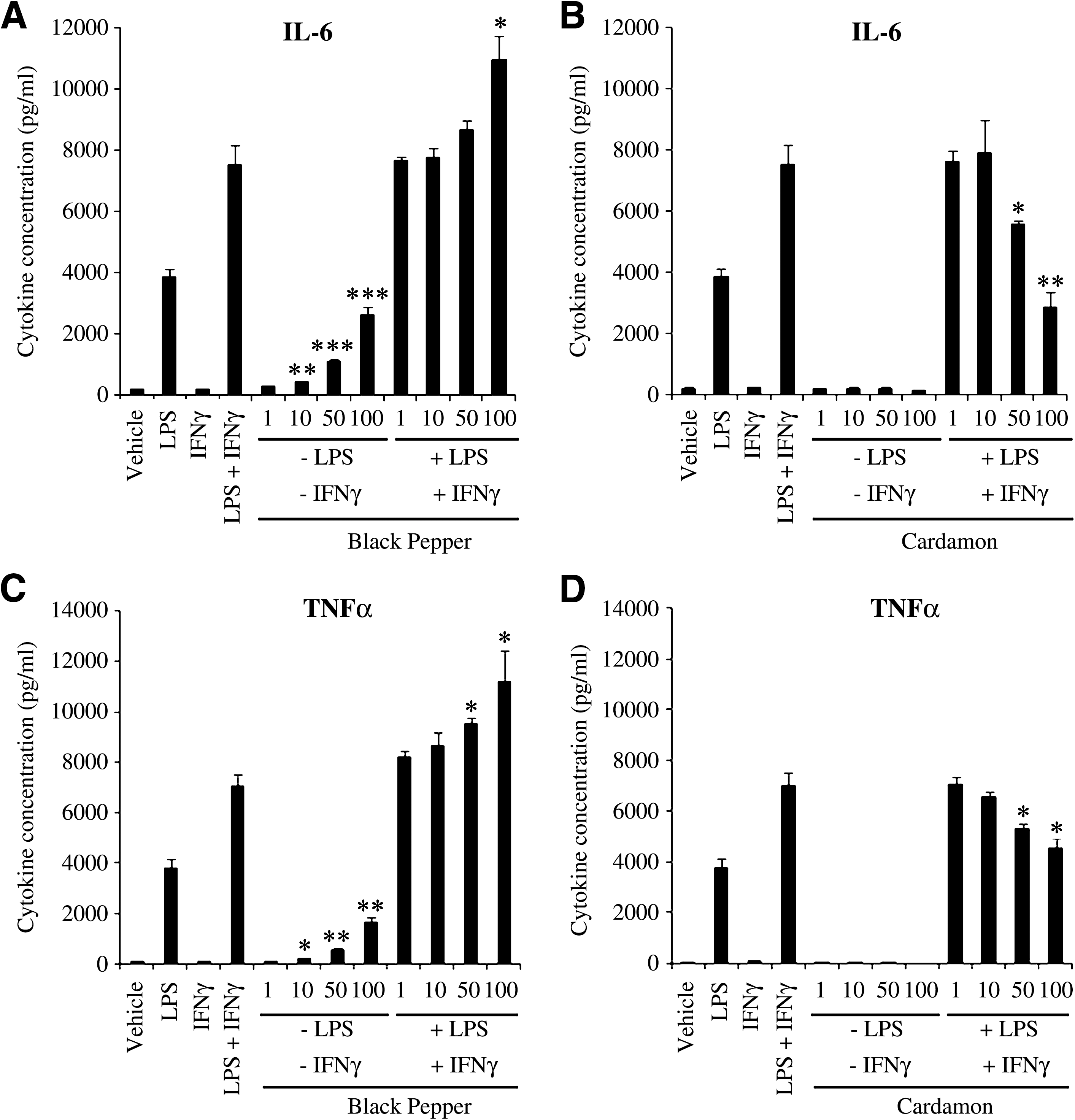
Effect of aqueous extracts of (
Assessment of NO production by macrophages in the presence of aqueous extracts of black pepper and cardamom
To assess the potential ability of aqueous extracts of black pepper and cardamom to modulate NO production by macrophages, BALB/c macrophages were cultured in medium supplemented with vehicle, IFNγ, LPS, or aqueous extracts of black pepper or cardamom at four different doses (1, 10, 50, and 100 μg/mL) in the presence or absence of IFNγ. Subsequently, cultured macrophages were subjected to Griess assay to measure NaNO2 production. As shown in Figure 4, LPS, IFNγ, and a combination of both caused macrophages to produce about 9, 40, and 99 μM NaNO2, respectively, compared to 6 μM NaNO2 in vehicle-treated peritoneal macrophages. Whereas the aqueous extract of cardamom had no significant effect on NO production by macrophages at any of the doses under unstimulatory conditions (Fig. 4B), the aqueous extract of black pepper at 100 μg/mL significantly enhanced NO production by macrophages compared to vehicle-treated macrophages under unstimulatory conditions (Fig. 4A). In the presence of IFNγ, the aqueous extract of black pepper significantly enhanced NO production by macrophages at 50 and 100 μg/mL compared to IFNγ-treated macrophages (Fig. 4A). Likewise, treatment of macrophages with 50 or 100 μg/mL black pepper extract in the presence of LPS and IFNγ led to a significant increase in NO production compared to macrophages treated with LPS and IFNγ (Fig. 4A). These findings suggest that the aqueous extract of black pepper mimics the LPS potential to prime macrophages. Interestingly, however, the aqueous extract of cardamom significantly suppressed NO production at 50 and 100 μg/mL doses in the presence of IFN compared to IFNγ-treated macrophages (approximately twofold suppression) (Fig. 4B). In the presence of both LPS and IFNγ, however, 100 μg/mL cardamom extract was the only dose capable of causing significant suppression of NO production by macrophages (Fig. 4B).

Dose-responsive effect of aqueous extracts of (
The aqueous extract of cardamom is capable of potently suppressing NO production by IFNγ-treated macrophages as well as IFNγ- and LPS-treated macrophages even in presence of the aqueous extract of black pepper (Fig. 4C). Indeed, the combined effect of the aqueous extracts of black pepper and cardamom on NO production is very comparable to the effect of the aqueous extract of cardamom alone in unstimulated, IFNγ-treated, and IFNγ- and LPS-treated macrophages (P = .22, P = .35, and P = .96, respectively) (Fig. 4C). This observation indicates that cardamom is a much more potent modulator of NO production than black pepper.
Evaluation of the cytotoxic activity of NK cells in the presence of aqueous extracts of black pepper and cardamom
To further assess the immunostimulatory effects of black pepper and cardamom, the cytotoxic activity of NK cells against YAC-1 tumor cells was evaluated in the presence of aqueous extracts of black pepper and cardamom. YAC-1 tumor cells were cultured in medium supplemented with vehicle, 100 μg/mL aqueous extract of black pepper or cardamom, and effector cells (NK cells) at E:T ratios of 200:1, 100:1, and 50:1. In addition, YAC-1 tumor cells were treated with 1, 10, 50, and 100 μg/mL aqueous extracts of black pepper and cardamom in the presence of effector cells at the 200:1 E:T ratio. As shown in Figure 5A, the aqueous extract of black pepper significantly enhances the cytotoxic activity of NK cells at 50 and 100 μg/mL doses. With regard to cardamom, significant dose-dependent stimulation of the cytotoxic activity of NK cells is observed at all doses (1, 10, 50, and 100 μg/mL) (Fig. 5B). Importantly, the aqueous extracts of black pepper and cardamom possess no direct cytotoxic activity against YAC-1 tumor cells (Fig. 5A and B). As expected, the enhanced cytotoxic activity of NK cells against YAC-1 tumor cells by aqueous extracts of black pepper and cardamom is proportional to the E:T ratio (data not shown). Interestingly, the aqueous extracts of black pepper and cardamom display synergistic stimulatory effect on the cytotoxic activity of NK cells against YAC-1 tumor cells (Fig. 5C), in which NK cytotoxic activity is much greater in the presence of both extracts compared to the separate action of each extract. Notably, the aqueous extract of cardamom displays a more potent stimulatory effect on the cytotoxic activity of NK cells compared to that of black pepper (Fig. 5C), but this differential ability to enhance the cytotoxic activity of NK cells did not reach statistical significance (P = .13). Taken together, these data strongly suggest that black pepper and cardamom have the potential to markedly enhance the anti-cancer activity of NK cells.
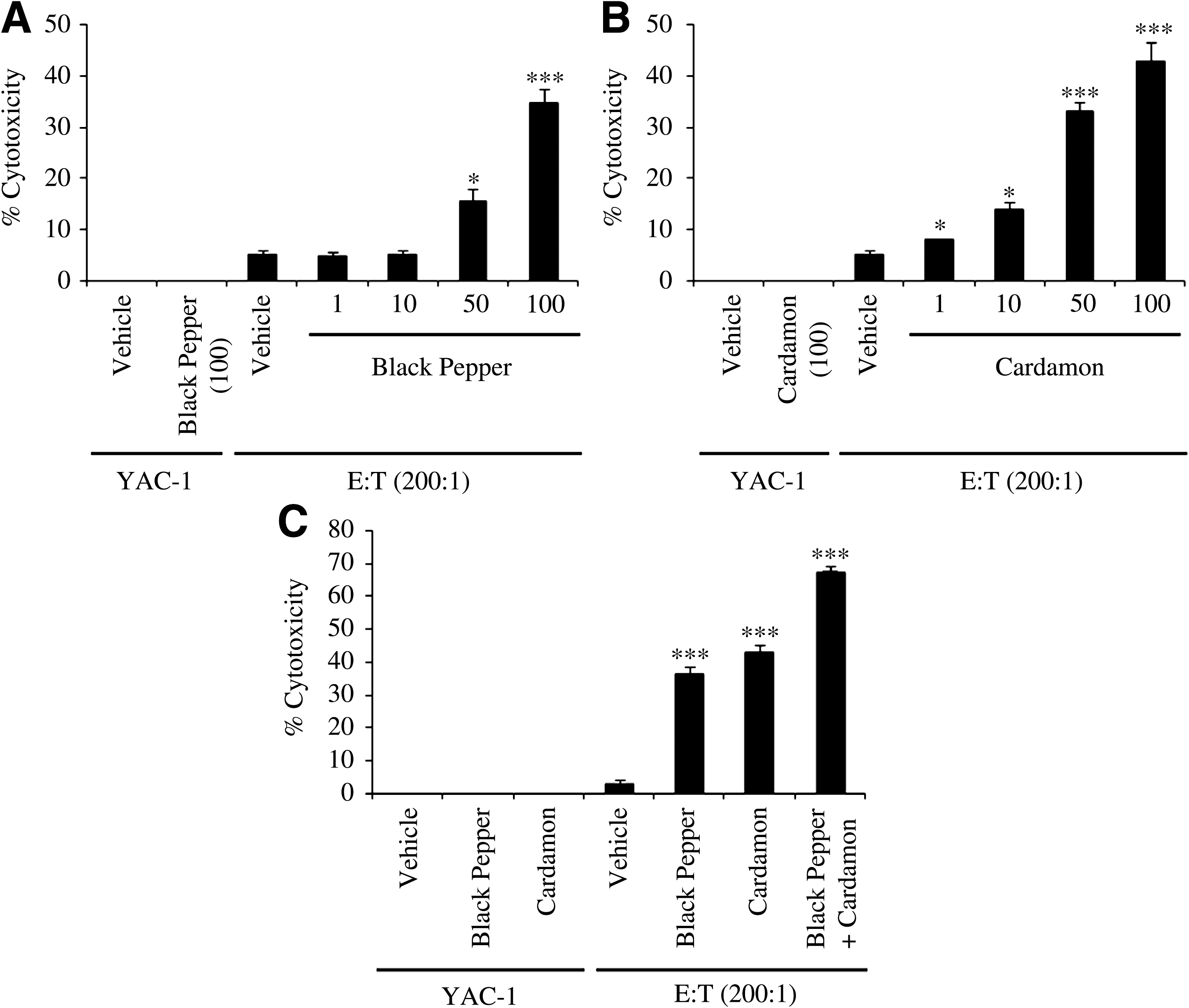
Dose-responsive effect of aqueous extracts of (
Discussion
A search for new drugs designed for promoting optimal immune function is the main focus of many researchers worldwide, and natural products used in traditional medicines have been the source of many medically beneficial drugs. 43,44 In search of such natural products, we investigated the potential immunomodulatory functions of black pepper and cardamom. In this study, we provide experimental evidence demonstrating that aqueous extracts of black pepper and cardamom are potentially capable of modulating the function of various immune cells. Indeed, the proliferation of splenocytes is significantly enhanced in the presence of aqueous extracts of black pepper and cardamom in a dose-dependent (Fig. 1A and B) and cooperative (Fig. 1C) fashion. This suggests that the aqueous extracts of black pepper and cardamom contain constituents that are capable of promoting the proliferative signaling pathways in splenocytes. Consistently, piperine, an active alkaloid component of black pepper, was shown to enhance murine splenocyte proliferation. 45 Similarly, eugenol, an active component of cardamom, has been reported to significantly enhance cell-mediated lymphocyte proliferation in vitro. 46
The immunomodulatory effects of spices on the release of major cytokines by splenocytes and macrophages have not been widely investigated. In our in vitro studies using primary splenocytes, the release of the Th2 cytokines IL-4 and IL-10 has been shown to be suppressed by black pepper extract and enhanced by cardamom extract (Fig. 2). Conversely, the release of the Th1 cytokine IFNγ has been shown to be enhanced and suppressed by aqueous extracts of black pepper and cardamom, respectively (Fig. 2). Consistently, piperine was shown to induce the secretion of Th1 cytokines IL-2 and IFNγ from splenocytes 45 while inhibiting Th2 cytokine secretion. 47 Moreover, and in agreement with our findings suggesting that the cardamom extract inhibits Th1 cytokine secretion and enhances Th2 cytokine secretion in splenocytes, eugenol was shown to be a potent inhibitor of the major Th1 cytokine IL-2. 48 Because Th2 cells are responsible for type-I hypersensitivity responses (i.e., allergic reactions), it is conceivable that the aqueous extract of cardamom, but not black pepper, may have the potential to induce type-I hypersensitivity reactions, development of Th2 cells, and establishment of Th2 immune responses. Interestingly, two studies have suggested that cardamom powder may be associated with contact and systemic contact-type dermatitis. 49 –51
It is evident that the release of the pro-inflammatory cytokines IL-6 and TNFα by macrophages is enhanced by the aqueous extract of black pepper (Fig. 3), indicating that black pepper promotes macrophage pro-inflammatory responsiveness. It is noteworthy that our data clearly indicate that the aqueous extract of black pepper significantly enhances NO production by macrophages only in the presence of IFNγ (Fig. 4A). This suggests that black pepper extract contains constituents that mimic the ability of LPS to prime macrophages for enhanced NO production. Conversely, the aqueous extract of cardamom impedes macrophage pro-inflammatory responsiveness, as reflected by the suppressive effects of the cardamom extract on IL-6, TNFα, and NO release by macrophages (Fig. 3). In agreement with these results, it was proposed that eugenol leads to inhibited secretion of the proinflammatory mediators IL-1β and IL-6, 52 inhibitory NO synthase and NO, 53,54 and cyclooxygenase-2. 55 Interestingly, oral administration of the aqueous extract of cardamom is accompanied by a significant reduction in cyclooxygenase-2 and inhibitory NO synthase expression in murine models of colon cancer. 36 Moreover, cardamom was shown to have anti-inflammatory activity against acute carrageenan-induced plantar edema in albino rats. 56 Taken together, the aqueous extract of black pepper exhibits pro-inflammatory activities, whereas the aqueous extract of cardamom exhibits anti-inflammatory activities in macrophages.
Our study also provides experimental evidence suggesting that the aqueous extracts of black pepper and cardamom enhance the cytotoxic activity of NK cells (Fig. 5A and B). Interestingly, the aqueous extracts of black pepper and cardamom cooperate to robustly augment the cytotoxic activity of NK cells to reach about 67% cytotoxicity (Fig. 5C). It is important to mention that neither the aqueous extract of black pepper nor that of cardamom had any direct cytotoxic effect against YAC-1 tumor cells (Fig. 5). These data strongly suggest that black pepper and cardamom possess immunostimulatory effects towards NK cytotoxic activity. Our findings regarding the anticarcinogenic effects of the black pepper and cardamom extracts are in agreement with other in vitro and in vivo studies. In one study, oral administration of black pepper extract significantly improved (by ∼65%) the life span of mice with Ehrlich ascites tumors. 20 In other studies, histopathological analyses have revealed that the rate of inflammatory cell infiltration into the submucosa, the incidence of papillae, and changes in the cytoplasm were decreased when rats with experimentally induced colon carcinogenesis were fed black pepper extract. 21,23 A recent study has shown that piperine has antiproliferative effects on human colon cancer cells. 24 Additionally, piperine was demonstrated to have antitumor activity in vivo. 57 –62 As for the proposed anticarcinogenic effects of cardamom, several studies have demonstrated that eugenol inhibits tumor formation in vivo. 63 –66 Consistent with our findings, eugenol has been shown to significantly enhance the NK cytotoxic activity, suggesting that cardamom exerts immunotoxic effects. 67
Although a large body of research has revealed that various spices and their chemical constituents could potentially play anticarcinogenic roles, the basis for such anticarcinogenic effects has been attributed to enzymatic modulation 26,34 –37 and anti-inflammatory, antiproliferative, and pro-apoptotic activities. 36 Indeed, enzymatic modulation has been deemed a mechanism of action by which black pepper manifests its anticarcinogenic roles. 22,23,25,26 Likewise, enzymatic modulation 26,34 –37 and anti-inflammatory, antiproliferative, and pro-apoptotic activities 36 have been proposed as mechanisms underlying the anti-cancer properties of cardamom. Our findings demonstrate that the anticarcinogenic effects of the black pepper and cardamom extracts may be attributed to the immunostimulatory potential of the spices' constituents to promote NK cytotoxic activity against cancer cells.
In conclusion, our study provides experimental evidence suggesting that black pepper and cardamom have a great potential to serve as immunomodulatory agents. Black pepper seems to play pro-proliferative, pro-inflammatory functions, while cardamom manifests itself as a potent suppressor of inflammation. In addition, our study demonstrates that black pepper and cardamom extracts exert anticarcinogenic effects via promoting the cytotoxic activity of NK cells. Although the exact molecular mechanisms underlying the immunomodulatory effects exerted by the extracts of black pepper and cardamom on splenocytes, macrophages, and NK cells are still unknown, elucidation of the specific signaling pathways involved in this immunomodulation is currently underway. Finally, we anticipate that the active constituents of black pepper and cardamom may serve as potential molecular tools for developing new therapeutic strategies to modulate inflammatory responses and prevent/treat various types of cancer.
Footnotes
Acknowledgments
We are grateful to Dr. Fredrick Palmer (Dalhousie University, Halifax, NS, Canada) for allowing us to use their rotatory evaporators in the preparation of the spice extracts. We thank Hana James, Jillian Tarrant, Wendy Hughes, and Bruce Musgrave for their invaluable technical assistance.
Author Disclosure Statement
No competing financial interests exist.