
Abstract
The potential of soy isoflavones (SIs) to reduce colon cancer has been investigated in animal models. These studies have found that outcomes are variable and depend on SI dose. The present study investigated dose–response effects of SIs on colon carcinogenesis in a chemically induced rat cancer model. Sprague-Dawley male rats were injected with 1,2-dimethylhydrazine (DMH) and were provided experimental diets that contained 0, 10, 50, 150, or 500 mg of SI aglycones/kg of diet for 12 weeks. Plasma concentrations of genistein, daidzein, and equol were determined using time-resolved fluoroimmunoassay. Plasma concentrations of these SIs tended to increase in a dose-dependent manner in DMH-treated rats. The numbers of aberrant crypt foci (ACF) and the expression of cyclooxygenase-2 (COX-2) proteins of colons were significantly decreased in the SI-fed groups compared with the control group; however, suppression was not dose-dependent. Furthermore, there were no significant correlations between plasma SI concentrations and ACF or COX-2 expression. Increased SI intake and increased plasma levels of SIs and metabolites were not associated with tissue levels of lipid peroxidation. We conclude that dietary supplementation of SIs suppresses DMH-induced ACF formation and COX-2 expression in a dose-independent manner.
Introduction
G
SIs are potential preventive agents against some hormone-dependent cancers such as breast and prostate cancer. 5 –8 Isoflavones bind to estrogen receptors (ERs), preferentially ER-β, with a much lower affinity than estradiol, and act either as an estrogen agonist, which leads to cell proliferation, or as an estrogen antagonist, which may lead to growth inhibition in hormone-dependent cancer cells. 9 Genistein is believed to be the primary bioactive isomer in soybeans and inhibits the proliferation of various cancer cells and normal or transformed intestinal epithelial cells. 10 Genistein and daidzein both bind to ERs; however, their binding affinities differ. Genistein has a much higher affinity for ER-α and ER-β compared with daidzein. 11 Furthermore, daidzein is converted to equol, which exhibits stronger estrogenic activity than the precursor molecule. 12 Isoflavones and their metabolites show antioxidant activity at concentrations within the range found in human plasma as measured by in vitro assay. 13 In one study, SI supplementation reduced oxidative stress, as measured by levels of oxidative DNA damage in human lymphocytes. 14 Additionally, genistein has been found to stimulate antioxidant protein gene expression in Caco-2 cells. 15
In moderation, isoflavones may exert beneficial effects and are probably not dangerous; however, for some cancers, large doses may increase risk. 16,17 In a previous study we observed an additive effect of high SI dose (1,000 mg/kg of diet) to that of fructooligosaccharide on the formation of aberrant crypt (AC) foci (ACF) and cyclooxygenase-2 (COX-2) expression in rats treated with 1,2-dimethylhydrazine (DMH). 18 The dose, 1,000 mg per kg, was far above normal physiological levels. Similar high doses frequently have been used in many studies of the anticancer efficacy of SIs. 19,20
Therefore, in the present study we investigated the effects of low to high SI doses on biomarkers of colon cancer such as ACF, colonic mucosa COX-2 expression, plasma concentrations of SIs, and tissue levels of lipid peroxidation in a chemically induced rat colon cancer model.
Materials and Methods
Animals and diets
Four-week-old male Sprague-Dawley rats (n = 60) were obtained from Daehan Biolink Co. (Chungbuk, Republic of Korea). All protocols were approved by the Animal Care and Use Committee of Daegu University (Gyeongsan, Republic of Korea). Two rats were housed per each wire-mesh bottom cage at a temperature of 20°C, 55% humidity, and a 12-hour light/12-hour dark cycle. Rats were given free access to standard rodent chow (Samyang Feed Co., Gangwon, Republic of Korea) and water and acclimated to the facility for 1 week. The experiment was designed as indicated in Figure 1. Fifty rats were injected subcutaneously twice per week for 4 weeks with DMH (20 mg/kg of body weight) in EDTA (1 mM, pH 6.7) to induce colon cancer. Ten normal rats were injected with the equivalent volume of EDTA solution without DMH on the same injection schedule as for DMH injection. The DMH-injected rats were divided into five groups of 10 rats each, and rats of each group were fed one of five experimental diets (Table 1) for a period of 12 weeks. The experimental diets were based on AIN93 diet 21 and contained 0, 10, 50, 150, or 500 mg of SI/kg of chow (Table 1). Normal rats that served as the negative control were fed the same SI-0 diet as the positive control (SI-0) group.
Experimental design and protocol. Fifty rats were injected with DMH (20 mg/kg of body weight) in EDTA (1 mM, pH 6.7) subcutaneously twice per week for 4 weeks. Ten normal rats were injected with the equivalent volume of EDTA solution on the same injection schedule as for DMH injection. After a 1-week rest, rats were fed experimental diets containing 0, 10, 50, 150 or 500 mg of SIs/kg for 12 weeks.
The amount of SI product (20.23% content based on aglycones) added to each diet to provide 10 mg, 50 mg, 150 mg, or 500 mg/kg of SI-10, SI-50, SI-150, or SI-500 diet, respectively.
Ingredients for the AIN93 diets were purchased from Harlan Teklad (Madison, WI, USA). The SI diet (Shindongbang Co., Seoul, Republic of Korea) contained 20.23% aglycones (8.18% daidzein, 9.85% glycitein, and 2.20% genistein).
Blood samples were taken from the abdominal aorta with the animal under diethyl ether anesthesia. Colons were removed, flushed with ice-cold saline, and slit open from the anus to the cecum on a cold plate. The distal 5-cm portion was excised and fixed in 10% buffered formalin solution. The mucosa of the remaining portion was scraped with a glass slide, frozen in liquid nitrogen, and stored at −70°C.
Quantification of ACF
The distal colon was fixed in 10% buffered formalin solution for 24 hours, stained in 0.2% methylene blue solution for 10–15 minutes, placed mucosal side up on a microscopic slide, and viewed using a microscope at × 40 magnification. The numbers of ACF and ACs observed per focus were recorded. 18,22
Measurement of plasma concentrations of genistein, daidzein, and equol
Glucuronides and sulfates were hydrolyzed, and genistein, daidzein, and equol were extracted and measured by time-resolved fluoroimmunoassay using genistein, daidzein, and equol kits (Labmaster Ltd., Turku, Finland). 23 All reagents were purchased from Wallac Oy (Turku, Finland). All reagents and samples were brought to room temperature (20–25°C) before use. An aliquot of 200 μL of 0.1 M acetate buffer (pH 5.0) containing 0.2 U/mL β-glucuronidase (Boehringer Mannheim, Mannheim, Germany) and 2 U/mL sulfatase (Sigma-Aldrich Co., St. Louis, MO, USA) was added to tubes containing 200 μL of plasma. After mixing, the samples were incubated overnight at 37°C. Hydrolyzed samples were extracted with 1.5 mL of diethyl ether using a vortex-mixer for 3 minutes. Samples were then frozen in a dry ice–ethanol mixture, and the ether phase was collected. After thawing, the water phase was re-extracted with ether in a disposable glass tube. The ether phases were combined and evaporated to dryness in a 45°C water bath. Assay buffer (200 μL) was added to the tubes, and after thorough mixing, 20 μL of the solution corresponding to 20 μL of the original plasma sample was added to each well. Time-resolved fluoroimmunoassay was performed according to the assay protocol (Labmaster Ltd.). We added 100 μL of anti-genistein antibody and 100 μL of genistein-Eu tracer and incubated the mixture at 25°C for 90 minutes with slow shaking. Wells were washed four times with 100 μL of wash buffer, 200 μL of enhancement solution was added, and the wells were incubated at 25°C for 5 minutes. Within 1 hour, genistein concentrations were determined using a Victor 1420 multi-label counter with software version 2.0 for fluorescence measurements (Perkin Elmer Life and Analytical Sciences, Inc., Turku, Finland). Daidzein and equol measurements were performed using the same procedures described above, but with a daidzein or equol kit.
COX-2 protein measurements
COX-2 protein was measured as described previously. 18,24 Frozen mucosa was homogenized in lysis buffer (10 mM Tris [pH 7.4], 5 mM EDTA, 150 mM NaCl, 1% Triton X-100, 10 mg/mL phenylmethylsulfonyl fluoride, 10 mg/mL aprotinin, 10 mg/mL leupeptin, 5 mM phenanthroline, and 28 mM benzamidine-HCl) at 4°C. After 30 minutes, the cell lysate was centrifuged for 30 minutes at 8,000 g at 4°C. The protein concentration of the supernatant was measured using the BCA Protein Assay Reagent (Bio-Rad, Hercules, CA, USA). The proteins were separated using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and blotted electrophoretically onto nitrocellulose membranes (Bio-Rad). The membranes were washed with Tris-buffered saline (10 mM Tris and 150 mM NaCl) containing 0.05% Tween 20. Membranes were blocked overnight at 4°C in Tris-buffered saline (10 mM Tris and 150 mM NaCl) containing 0.05% Tween 20 and 5% nonfat dried milk and were subsequently incubated with rabbit anti-rat COX-2 polyclonal antibody (Abcam Ltd., Cambridge, UK) at room temperature for 2 hours. After washing, the membranes were treated with goat polyclonal antibody to heavy and light chain rabbit immunoglobulin G (Cy5-conjugated; Abcam Ltd.). The COX-2 protein band densities were measured by a fluorescence imaging analysis system (Typhoon 9200, Amersham Biosciences, Piscataway, NJ, USA) and are expressed in densitometer units. Immunoblot analysis using rabbit polyclonal anti-β-actin (Abcam Ltd.) was performed as a control for sample loading.
Thiobarbituric acid-reactive substances (TBARS) measurements
Tissues (liver and kidneys) were frozen in liquid nitrogen, immediately crushed in a prechilled mortar and pestle, and disrupted in a glass homogenizer. TBARS levels in plasma and homogenates of liver and kidneys were determined using TBARS assay kits (ZeptoMetrix Corp., Buffalo, NY, USA). All reagents were allowed to reach room temperature before use. Sodium dodecyl sulfate solution was added to the test tubes before the thiobarbituric acid buffer reagent was added. The mixture was then forcefully applied down the side of each tube, and each tube was covered with a glass marble and incubated at 95°C for 60 minutes. Test tubes were then removed from incubation and cooled to room temperature in an ice bath for 10 minutes. Cooled samples were centrifuged at 500 g for 15 minutes, and TBARS were quantified by reading the absorbance at 532 nm.
Statistical analysis
The results are expressed as mean ± SEM values. The SPSS release 15.0 software package (SPSS Inc., Chicago, IL, USA) was used for all statistical analyses. Differences in means between the groups that were significant at P < .05 using a one-way analysis of variance were tested by Duncan's multiple range tests at P < .05. To test correlations among variables, Pearson's correlation coefficients were calculated.
Results
Body weight gain and food intake
Weight gain, food intake, and food efficiency ratios were independent of SI dose in DMH-treated rats (Table 2). Average intakes of SI based on body weight and amounts consumed per day were calculated from SI content of the experimental diets and from food intake levels. The daily intakes of SIs per kg of body weight taken from diets containing 10, 50, 150, and 500 mg of SIs/kg were 0.58, 2.95, 8.35, and 29.48 mg, respectively. Daily intakes of SIs per animal from diets containing 10, 50, 150, and 500 mg of SIs/kg diet corresponded to 0.23, 1.14, 3.20, and 11.27 mg, respectively.
Data are mean ± SEM values (n = 10) except for food intake and food efficiency ratio (n = 5 [two rats pooled]).
Means without a common superscript letter are significantly different from each other using Duncan's multiple range test at P < .05.
Plasma concentrations of SIs
Plasma concentrations of genistein and daidzein increased rapidly with SI dose up to 500 mg/kg, as shown in Figure 2. Equol concentration significantly increased with 150 mg/kg SIs in the diet. Equol concentrations were the highest among the three SIs. Plasma concentrations tended to increase with increasing SI intake (mg/kg of body weight), as shown in Figure 3. Correlation coefficients of plasma levels of genistein, daidzein, and equol with SI were r = 0.703 (P < .001), r = 0.765 (P < .001), and r = 0.740 (P < .001), respectively.
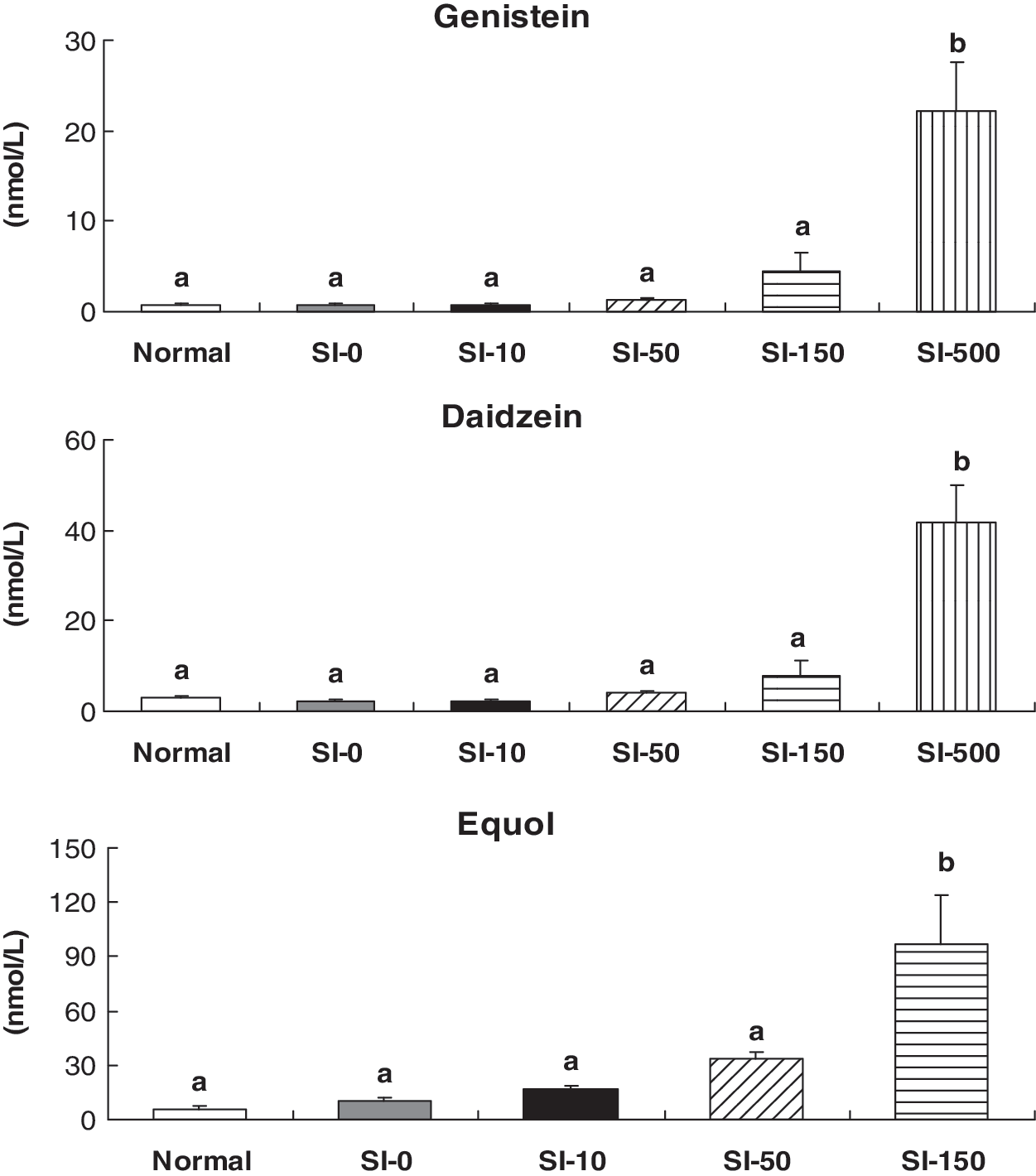
Plasma concentrations of genistein, daidzein, and equol in DMH-treated rats fed different doses of SIs. Data are mean ± SEM values (n = 7–10). Means without common superscript letters are significantly different from each other using Duncan's multiple range test at P < .05.

Relationship between SI intake and plasma concentrations of genistein, daidzein, and equol in DMH-treated rats. Correlation coefficients of genistein, daidzein, and equol are r = 0.703 (P < .001), r = 0.765 (P < .001), and r = 0.740 (P < .001), respectively. B.W., body weight.
ACF and COX-2
Numbers of ACF and total ACs and ACF with four or more ACs per focus are shown in Table 3. Numbers of ACF and total ACs of the positive control (SI-0) group were increased 22- and 35-fold, respectively, compared to the negative control (normal) group. No ACF with four or more ACs per focus were observed in the normal group. This indicates that mucosal ACF were induced specifically by DMH treatment. Numbers of ACF and total ACs were significantly decreased in the SI-fed groups, particularly the SI-10 and SI-50 groups, compared with the control group. Total ACF and total AC numbers of the SI-500 group tended to decrease compared with the control group; however, levels were not significantly different from those of the control group. COX-2 protein expression (Fig. 4) was significantly decreased in the SI-fed groups compared with the control group. No significant correlations, however, were observed between plasma concentrations of SIs and ACF formation or COX-2 expression.

COX-2 protein levels in the colons of DMH-treated rats fed different doses of SIs. Data are mean ± SEM values (n = 10). Means without common superscript letters are significantly different from each other using Duncan's multiple range test at P < .05.
Data are mean ± SEM values (n = 10).
Means without a common superscript letter are significantly different from each other using Duncan's multiple range test at P < .05.
Tissue levels of TBARS
Plasma levels of TBARS were not significantly different among the groups (Table 4). Liver levels tended to be lower in the 10 and 50 mg of SI-fed groups compared with the control (SI-0) group. Kidney levels of the 50 mg of SI-fed group were significantly lower than the control group. Collectively, TBARS levels in the plasma, liver, and kidneys were not dose-dependently responsive to plasma levels of SIs.
Data are mean ± SEM values (n = 10).
Means without a common superscript letter are significantly different from each other using Duncan's multiple range test at P < .05.
Discussion
The major functions of SIs originate from their estrogenic properties. Cell-based studies provide a large body of evidence on the functional effects of isoflavones. Specifically, gene expression is modified, i.e., turned on or off, by purified isoflavones. 25 Previous studies have found an SI dose-dependent inhibition of human colonic tumor cell proliferation in culture. 26 Conflicting results from many studies using animal models of breast cancer have shown that SIs inhibit, as well as promote, the growth of mammary tumors. 8
Studies on the inhibitory effects of SIs on ACF formation and colon tumor formation in an azoxymethane-induced rat colon cancer model also have produced some conflicting results. Some results are positive, 18 –20 whereas others are negative. 27,28 According to a study performed by Thiagarajan et al., 19 soybean or soy flour diets containing 0.049% genistein glycoside derivatives reduce the formation of ACF; however, these diets were less effective in inhibiting ACF compared with diets containing 0.015% (150 mg/kg) genistein as aglycone. In contrast, Daly et al. 17 reported that dietary SI supplementation at a concentration of 2,000 mg/kg exerts adverse effects on older female rats injected with azoxymethane. According to the work of Raju et al., 29 pre- and postnatal exposure to dietary SIs suppresses the growth of colon tumors in male rats. A dose of 40 mg/kg decreased mean tumor size per group, mean tumor size per tumor-bearing rat, and tumor burden compared with control rats that were not fed SIs. However, a 1,000 mg/kg dose did not decrease tumor variables (excluding tumor burden) compared with the control group. Although the expression of ER-β mRNA increased in the tumors of rats receiving either 40 mg/kg or 1000 mg/kg SIs compared with the control, no differences were found between the two doses.
In this study, the inhibitory effects of SIs on suppression of colon cancer biomarkers such as ACF and COX-2 expression were positive. However, rats fed the diet with 50 mg/kg showed significantly fewer numbers of ACF and total ACs compared to rats fed the 500 mg/kg dose. The 50 mg/kg dose is comparable to levels that could be achieved in humans taking SI supplements. COX-2 expression, which is another type of colon cancer biomarker, 30,31 was suppressed in rats fed diets supplemented with 10–500 mg/kg; however, there was no difference in COX-2 expression based on SI dose. The distal 5 cm of colon amounting to about one-third of the whole colon is appropriate to observe ACF because ACs and DMH-induced tumors are located predominantly in the distal colon. 32 The remaining colonic tissue used for COX-2 protein measurement may not be as sensitive as the distal part of colon for DMH-induced COX-2 expression. Also, the biomarkers, ACF and COX-2 expression, may be insufficient to represent mechanistic explanation of colon tumorigenesis.
Despite the increased plasma concentrations of genistein, daidzein, and equol with increasing amounts of SI intake, plasma concentrations of SIs and colon cancer biomarkers were not significantly correlated. These results suggest that plasma SI concentration is not directly related to the suppression of colonic carcinogenesis.
The SIs used in this study consisted of 40.4% daidzein, 48.7% glycitein, and 10.9% genistein. The level of genistein used in this study is 54.4 mg/kg of diet even in the dose for the 500 mg/kg diet. The adverse effects of genistein reported in the literature have been often related to high doses (250 mg/kg and 1,200 mg/kg). 16,17,28 Rao et al. 28 have shown an enhancing effect of 250 mg of genistein/kg of diet on noninvasive and total adenocarcinoma multiplicity. Moreover, Gee et al. 16 emphasized that, when SIs were fed immediately before treatment with DMH, pure genistein or soya protein isolate equivalent to 250 mg of genistein/kg of diet was associated with a three- or twofold increase, respectively, in ACF in the distal colon. However, we have tested pretreatment effect of SI supplementation before injection of DMH and have not found an enhancing effect of SIs up to 500 mg/kg of diet (genistein up to 54.4 mg/kg of diet) on ACF (authors' unpublished data).
Oral administration of isoflavones leads to significant phase II metabolism. Subsequently, isoflavone glucuronides or sulfates become the major circulating forms in the plasma. 33 This metabolism occurs mainly in the liver and epithelial cells of the intestinal wall, which contributes to the first-pass effect. Genistein exists in the systemic circulation after being absorbed as several molecular forms. Its metabolites are then excreted into bile and enterohepatic recirculation. According Kwon et al., 34 the bioavailability of genistein is 38.58%, 24.34%, and 30.75% after oral administration of genistein at doses of 4, 20, and 40 mg/kg of body weight, respectively. Zhou et al. 35 also reported that oral absolute bioavailability was 21.9%, 33.5%, and 19.0% after oral administration of genistein at doses of 6.25, 12.5, and 50 mg/kg of body weight, respectively. Absorption was reduced with the high dose, i.e., 50 mg/kg of body weight, which indicates levels approaching saturation. In the present study, genistein and daidzein intakes at an SI dose of 500 mg/kg corresponded to 3.21 and 12.19 mg/kg of body weight, respectively. Therefore, we hypothesize that the doses in the present study are far below the absorption saturation levels, as indicated in Figure 3.
We observed that increased plasma SI concentrations were not related to tissue levels of lipid peroxidation, which may reflect long-term antioxidative effects of SI. Wei et al. 36 reported that genistein strongly inhibits tumor promoter-induced hydrogen peroxide formation in vitro and in vivo and emphasized the antioxidant properties of genistein. In azoxymethane-treated rats, administration of 250 mg of genistein/kg of diet significantly inhibited levels of 8-isoprostane in colonic mucosa and in turmors, indicating that genistein inhibits the free radical-catalyzed lipid peroxidation. However, the suppression of 8-isoprostane was independent of the colon tumor enhancement. 28 Rufer and Kulling 13 compared the antioxidant capacities of isoflavones and their corresponding metabolites with those of ascorbic acid and quercetin. Oxidative metabolites of genistein and daidzein, as well as of equol, exhibited very high antioxidant activity in an oxygen radical absorbance capacity assay and an in vitro low-density lipoprotein oxidation assay, with the conjugated diene and TBARS formation as end points. Genistein and daidzein were effective only in the oxygen radical absorbance capacity assay; daidzein and genistein were not effective in inhibiting TBARS formation compared with ascorbic acid and quercetin. In this study, TBARS levels in plasma, liver, and kidneys were not significantly inhibited with increasing doses of SIs. These results are consistent with findings suggesting that the consumption of SIs does not promote acute postprandial antioxidant capacity 37 and has little effect on plasma antioxidant capacity and biomarkers of oxidative stress 38 in humans. Therefore, in vitro phenomena, such as antioxidative activity, could be markedly different in in vivo biological activities. 39
Cooke 40 reviewed data from available studies of colon cancer in rodent models and stated that although SIs may afford some protection against the progression of colon cancer, isoflavones do not prevent further development once the cancer has progressed beyond the hormone-dependent stage. In the present study, moderate levels of SIs appeared to be most effective in suppressing ACF formation in a DMH-induced colon cancer rat model.
Therefore, we conclude that the initiation or promotion stage of colon cancer may be inhibited with moderate consumption of SIs. Further, suppression of colon carcinogenesis by SIs is dose-independent. High plasma concentrations of genistein, daidzein, and equol caused by a high SI dose did not affect ACF formation, COX-2 expression, or the inhibition of lipid peroxidation in tissues. We need to have studies of mechanistic aspects of SIs in reducing colon tumor formation more specifically in colonic tissues.
Footnotes
Acknowledgment
This research was supported by a 2008 research grant from Daegu University.
Author Disclosure Statement
No competing financial interests exist.