
Abstract
We tested the correlation between the tightness of tight junctions (TJs) and the anti-invasive activity of the ethyl alcohol extract of Hizikia fusiforme (EHF) in Hep3B human hepatocarcinoma cells. EHF inhibited cell proliferation, motility, and invasiveness, which were associated with increased TJ tightness, as demonstrated by an increase in transepithelial electrical resistance. EHF dose-dependently decreased the secretion of matrix metalloprotease-2 and -9, which correlated with a decrease in mRNA and protein expression, but increased tissue inhibitor of metalloproteinase-1 and -2 mRNA levels. Additionally, immunoblotting results indicated that EHF suppressed the major components of TJ, claudins (-1, -3, and -4), which play a key role in the control and selectivity of paracellular transport. These data indicate that EHF may inhibit cancer cell invasion through the tightening of TJs, which may counteract the up-regulation of claudins. Furthermore, EHF treatment decreased the expression of insulin-like growth factor-1 receptor proteins, while concurrently increasing that of thrombospondin-1 and E-cadherin. In conclusion, these results suggest that EHF treatment may inhibit tumor metastasis and invasion and therefore act as a dietary resource for decreasing the risk of developing cancer.
Introduction
T
Cancer cells are characterized by a high rate of proliferation, turnover, and polarization, with a structurally distinct apical side. Two parameters—the movement and invasiveness of cancer cells—are important for their ability to spread or metastasize to other sites. 9 During metastasis, cancer cells undergo disruption of the cell–cell junction and also dysregulation of junctional complex proteins. 10,11 Tight junctions (TJs) are localized at the apical end of the lateral membrane surfaces of epithelial cells and function as the primary barrier to the paracellular transport of solutes across the cell membrane. 12,13 Claudins, novel integral membrane proteins, are major cell adhesion molecules at the TJs. Claudins contain four transmembrane domains, two extracellular loops, and an intracellular carboxyl tail. 14,15 Distinct patterns of claudin expression are found at the TJs in different organs and tissues. Altered claudin expression occurs in a wide variety of human malignancies, including colonic, 16 pancreatic, 17 breast, 18 liver, 19 and prostate 20 cancer.
The inverted expression of matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs) suggests that MMPs function in cancer invasion, metastasis, and progression. 21,22 For example, claudin-10 overexpression in Hep3B cells resulted in up-regulation of MMP and other members of the claudin family. 23 Claudin-1 also modulates MMP activity, 24 E-cadherin, an adherent junction protein, regulates TJ formation, 25 and high thrombospondin-1 (TSP-1) expression is also inversely correlated with invasiveness and lymph node metastasis. 26,27 Moreover, insulin-like growth factor-1 receptor (IGF-1R) signaling conveys invasive and metastatic capability in human tumors. 17,18
In the course of our screening program of bioactive marine natural products from seaweeds, we found the ethyl alcohol extract of H. fusiforme (EHF) has strong antiproliferative effects in human cancer cells by inducing apoptosis. Recently, we also reported that treatment with tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) in combination with subtoxic concentrations of EHF sensitizes TRAIL-resistant AGS human gastric carcinoma cells to TRAIL-mediated apoptosis. The results suggest that the combined treatment of AGS cells with EHF plus TRAIL may provide an effective and selective therapeutic strategy. 28 In the present study, we examined the effects of EHF on invasive parameters, including Matrigel™ (BD Biosciences, Bedford, MA, USA) membrane invasion, zymography for MMP activities, and gene expression for the TJ- and metastasis-associated proteins, using a Hep3B human hepatocellular carcinoma cell line.
Materials and Methods
Preparation of EHFs, cell culture, and cell viability
H. fusiforme, harvested along the southern coast of Korea, was soaked in tap water for 24 hours to remove impurities and then dried in the shade for 5 days. Dried seaweeds were extracted in 80% ethyl alcohol at a ratio of 10 mL/g for 24 hours in a refrigerator at 4°C. The EHFs were vacuum-filtered through a Whatman (Maidstone, UK) no. 2 filter and lyophilized, yielding a dried residue, and then kept at −80°C. The powder was dissolved in dimethyl sulfoxide as a stock solution at 100 μg/mL and stored in aliquots at −20°C. Human hepatocelluar carcinoma Hep3B cells were obtained from the American Type Culture Collection (Rockville, MD, USA) and cultured in Dulbecco's Modified Eagle's Medium (Invitrogen Corp., Carlsbad, CA, USA) supplemented with 10% heat-inactivated fetal bovine serum (GIBCO-BRL, Gaithersburg, MD, USA), 2 mM glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin (GIBCO-BRL) at 37°C in a 5% CO2 incubator. For the cell viability study, Hep3B cells were grown to 70% confluence and treated with EHF. Control cells were supplemented with complete medium containing 0.1% dimethyl sulfoxide (vehicle control). Following treatment, cell viability was determined by 3-(4,5-dimethyl-2 thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) (Sigma, St. Louis, MO, USA) assays, which is based on the conversion of MTT to MTT-formazan by mitochondria. 29
Wound healing migration assay
Cells were grown to confluence on 30-mm-diameter cell culture dishes coated with rat tail collagen (20 μg/mL) (BD). A scratch was made in the cell layer with a pipette tip. After washing with phosphate-buffered saline, serum-free medium (to prevent cell proliferation) containing either vehicle (dimethyl sulfoxide) or 300 μg/mL EHF was added. Photographs of the wounded area were taken immediately after the scratch was made and 48 hours later to monitor cell movement into the wounded area.
Measurement of transepithelial electrical resistance (TER)
TER was measured with an EVOM epithelial tissue volt-ohm meter (World Precision Instruments, Sarasota, FL, USA), equipped with a pair of STX-2 chopstick electrodes. In brief, Hep3B cells were seeded into the 8.0 μm pore size insert (upper chamber) of a Transwell® (Corning Costar Corp., Corning, NY, USA) and allowed to reach full confluence, after which fresh medium was replaced for further experiments. Inserts without cells, inserts with cells in medium, and inserts with cells with EHF were treated for 48 hours. Electrodes were placed at the upper and lower chambers, and resistance was measured with the volt-ohm meter.
Matrigel invasion assay
Matrigel invasion assays were used to assess the ability of the Hep3B cells to penetrate the extracellular matrix in the presence or absence of EHF as previously described. 30 In brief, the cells were exposed for 6 hours to EHF, and the treated cells (50,000) were then plated onto the apical side of the Matrigel-coated filters in serum-free medium containing either dimethyl sulfoxide or EHF. Medium containing 20% fetal bovine serum was placed in the basolateral chamber to function as a chemoattractant. After 48 hours, the cells on the apical side were wiped off with a cotton-tipped swab. Cells on the bottom of the filter were stained with hematoxylin (Sigma) and counted (three fields of each triplicate filter) with an inverted microscope.
RNA extraction and reverse transcription-polymerase chain reaction
Total RNA was prepared using an RNeasy kit (Qiagen, La Jolla, CA, USA) and primed with random hexamers to synthesize cDNA using AMV reverse transcriptase (Amersham Corp., Arlington Heights, IL, USA) according to the manufacturer's instructions. Polymerase chain reaction was performed in a Mastercycler® (Eppendorf, Hamburg, Germany) with the primers indicated in Table 1. Conditions for PCR reactions were once at (94°C for 3 minutes), 35 times at (94°C for 45 seconds, 58°C for 45 seconds, and 72°C for 1 minute), and once at (72°C for 10 minutes). Amplification products obtained by polymerase chain reaction were separated on a 1% agarose gel and visualized by ethidium bromide staining.
GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Protein extraction and western blot analysis
Total cell lysates were prepared in an extraction buffer (25 mM Tris-Cl [pH 7.5], 250 mM NaCl, 5 mM EDTA, 1% Nonidet P-40, 0.1 mM sodium orthovanadate, 2 μg/mL leupeptin, and 100 μg/mL phenylmethylsulfonyl fluoride). Protein concentration was determined with a Bio-Rad Laboratories (Hercules, CA) protein assay kit. For western blot analysis, proteins (30–50 μg) were separated by 8–13% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred to a nitrocellulose membrane (Schleicher & Schuell, Keene, NH, USA). The membranes were blocked with 5% skim milk for 1 hour, and the membranes were subjected to immunoblot analysis with the desired antibodies. Proteins were then visualized by the enhanced chemiluminescence method according to the recommended procedure (Amersham). Primary antibodies were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA) and Calbiochem (Cambridge, MA, USA). Peroxidase-labeled donkey anti-rabbit immunoglobulin and peroxidase-labeled sheep anti-mouse immunoglobulin were purchased from Amersham.
Gelatin zymographic analysis of secreted MMPs
After incubation with EHF for 48 hours, cell culture supernatants were collected and centrifuged at 400 g for 5 minutes. The cell-free supernatant was mixed with 2 × sample buffer (Invitrogen), and zymography was performed using precast gels (10% polyacrylamide and 0.1% gelatin). After electrophoresis, the gels were washed twice at room temperature for 30 minutes in 2.5% Triton X-100, subsequently washed in buffer containing 50 mM Tris-HCl, 150 mM NaCl, 5 mM CaCl2, 1 μM ZnCl2, and 0.02% NaN3 at pH 7.5, and incubated in this buffer at 37°C for 24 hours. Thereafter, the gels were stained with 0.5% (wt/vol) Coomassie Brilliant Blue G-250 (Bio-Rad) for 1 hour and then lightly destained in methanol:acetic acid:water (3:1:6 by volume). Clear bands appeared on the Coomassie-stained blue background in the areas of gelatinolytic activity. Gels were scanned, and images were processed extracting the blue channel signal, converting it to black and white and inverting it to quantify the gelatinolytic activities from the integrated optical density.
Statistical analysis
Data are expressed as mean ± SD values. Significant differences among the groups were determined using the unpaired Student's t test. P < .05 was accepted as an indication of statistical significance. All means were obtained from at least three independent experiments.
Results
EHF inhibits the growth and motility of Hep3B cells
To investigate the effects of EHF on cell viability, Hep3B cells were treated with EHF and subjected to MTT assays. Treatment with 300 μg/mL and 500 μg/mL HEF caused approximately 12% and 68% inhibition of cell growth, respectively, compared to controls (Fig. 1A). Furthermore, EHF (300 μg/mL) treatment, which was not cytotoxic, reduced cell motility compared with controls in the wound healing assay (Fig. 1B).
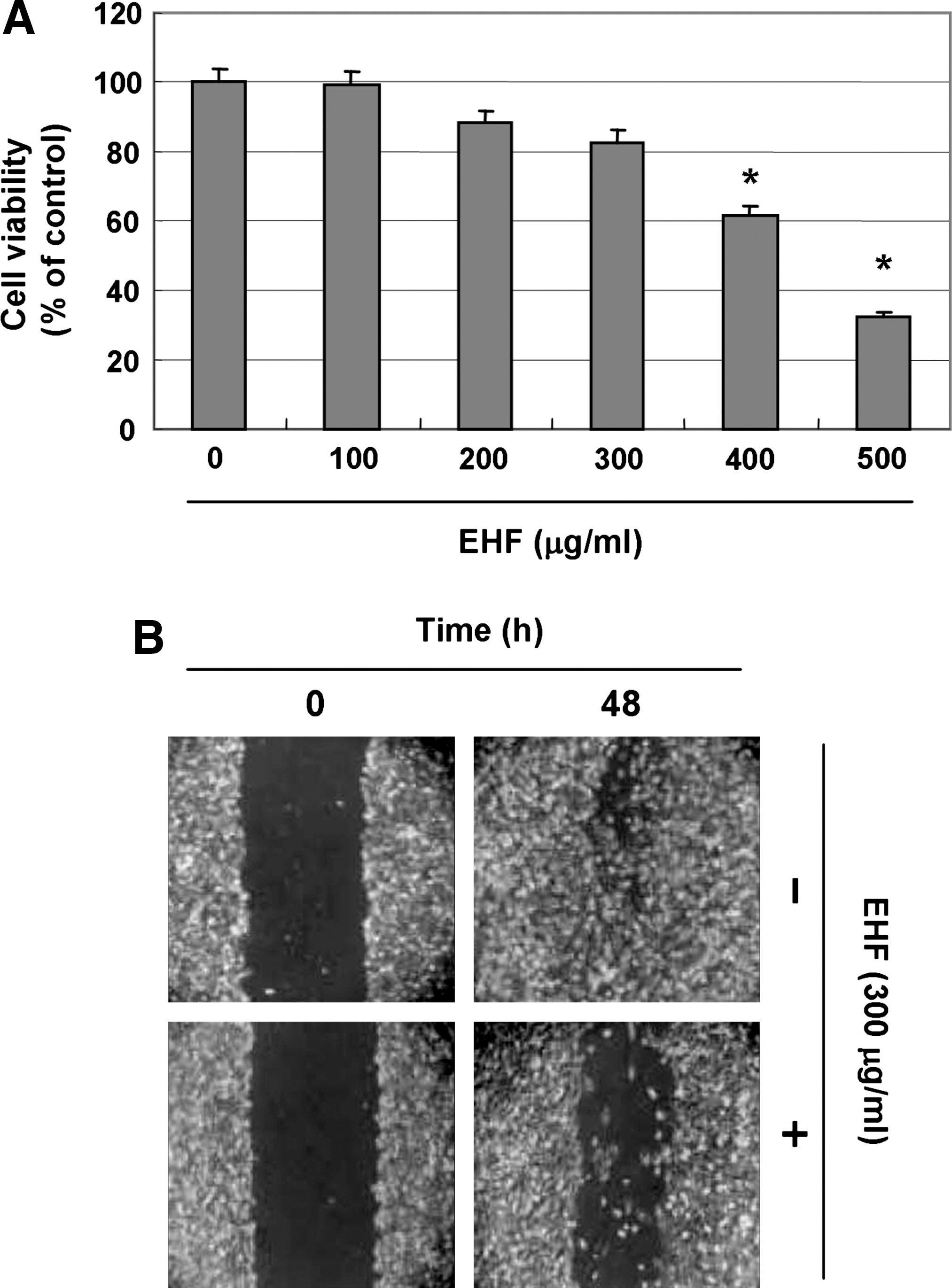
Inhibition of cell growth and motility of human hepatocarcinoma Hep3B cells by EHFs. (
EHF increases TER values and decreases cell invasion
To examine the relationship between TJ tightening and invasive activity, we measured TER (a measure of TJ formation). EHF treatment dose-dependently increased TER levels (Fig. 2A), and removal of EHF allowed TER to return to baseline levels (Fig. 2B). These results suggest that the TER increase induced by EHF involves a stable, reversible change. We next examined whether EHF decreases cell invasion using a Boyden chamber invasion assay. EHF treatment reduced cell invasion through the Matrigel chamber in a concentration-dependent manner (Fig. 2C and D), suggesting that increases in TER correlated with the inhibition of invasion.

Effect of EHF on TER values and invasion in Hep3B cells. (
EHF inhibits the activities and expression of MMP-2 and -9
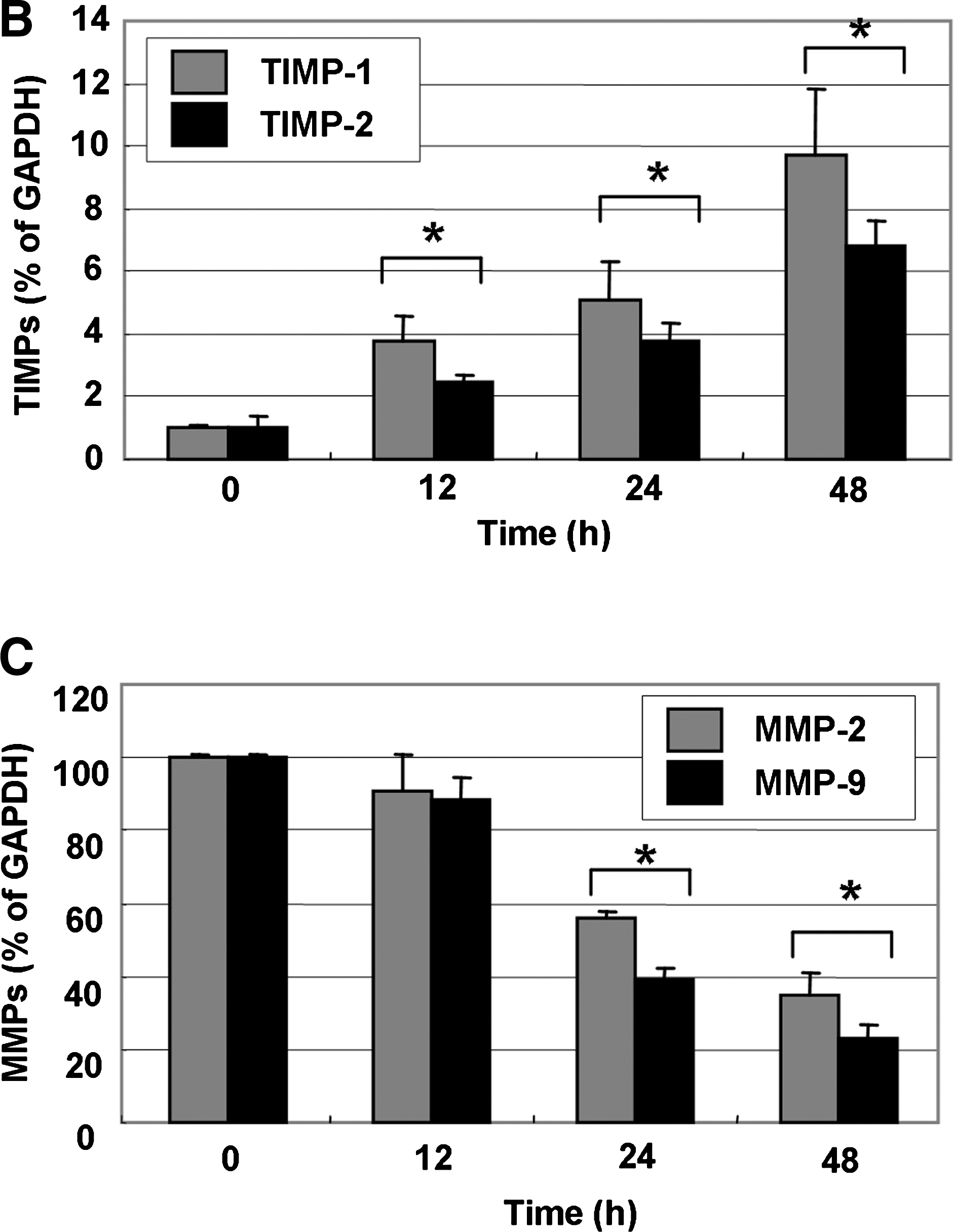
Because migration influences metastasis and invasion of the basement membrane is primarily mediated by the gelatinase MMPs, we tested the effects of EHF on TIMP and MMP mRNA levels by reverse transcription-polymerase chain reaction. EHF time-dependently increased TIMP-1 and -2 mRNA levels but decreased MMP-2 and -9 levels (Fig. 3). We next investigated the effects of EHF on the protein levels and activities of MMPs using western blot and gelatin zymography. HEF dose-dependently decreased MMP-2 and -9 activities, which was consistent with the concurrent down-regulation of their protein levels (Fig. 4). These results suggest that the anti-invasive effect of EHF is associated with increased TIMP-1 and -2 levels as well as inhibition of MMP-2 and -9 mRNA, protein expression, and activity in Hep3B cells.

Effect of EHF on TIMP and MMP mRNA expression in HEP3B cells. (

Effect of HEF on MMP-2 and -9 protein expression and activities in Hep3B cells. (
EHF modulates the expression levels of TJ and metastasis proteins
To elucidate the mechanism by which EHF enhances TJ activity, we determined the levels of TJ components and TJ regulators, claudins, E-cadherin, IGF-1R, and TSP-1, using western blot analyses. EHF treatment time-dependently decreased levels of claudin proteins (claudin-1, -3 and -4), the most important components of the TJ, 11,12 suggesting that this modulation contributed to TJ tightening. EHF treatment also decreased protein levels of IGF-1R, a promoter of tumor invasion and metastasis in various human cancers, 17,18 but increased levels of E-cadherin, an adherent junction protein that regulates TJ formation. 25 EHF treatment also time-dependently increased the expression of TSP-1, which limits tumor growth 26,27 (Fig. 5).

Effect of EHF on expression of claudins, E-cadherin, IGF-1R, and TSP-1 proteins in Hep3B cells. (
Discussion
Because cancer cell invasion and migration are critical steps in metastasis, the inhibition of tumor cell invasion and metastasis are important mechanisms for anticancer drugs. The aim of this study was to investigate whether H. fusiforme has potent anti-invasion and antimetastasis activities in human hepatocarcinoma Hep3B cells. We found that EHF markedly inhibited invasive activity by tightening TJs, decreasing MMP activity, and modulating TJ and metastasis proteins.
Alterations in cell permeability and loss of cell polarity are hallmarks of epithelial cell tumorigenesis. TJs, the critical structures for maintaining these functions in epithelial cells, are modulated in a number of epithelial cancers, including hepatocellular carcinoma. 12,13 Thus, TJ disruption and the dysregulation of its composite proteins play critical roles in cancer progression, invasion, and metastasis, especially epithelial cancers. 14,15 Soler et al. 31 first demonstrated that the TER of colon carcinoma tissue was significantly lower than that of normal colon tissues but showed higher transepithelial paracellular permeability, which confirmed the loss of the TJs. Butyrate, prostasin, and tumor necrosis factor-α can inhibit motility and invasiveness by enhancing transepithelial paracellular permeability. 32 –34 Here, treatment with EHF dose-dependently increased the TER of Hep3B cells, which was associated with lower cell motility and invasiveness (Figs. 1 and 2). Elevated TER levels gradually returned to baseline when EHF was removed from the culture medium, here and in another hepatocarcinoma cell line, HepG2 (data not shown). These results indicate that HEF may prevent or reverse TJ leakiness. Because TJ leakiness is associated with cancer progression, TJ tightening may have anticancer activity. 9 Thus, the anti-invasive activity of HEF may be due, in part, to its ability to enhance TJ activity.
MMPs are important proteolytic enzymes in organ development and tissue regeneration, but they also play important roles in cancer invasion and metastasis. In particular, MMP-2 and -9 play important roles in tumor invasion and angiogenesis, and tumor metastasis can be inhibited by blocking MMP synthesis and activity. 21,22 For example, Hazgui et al. 35 reported that the antimetastatic activity of a polyphenolic constituent of green tea, epigallocatechin-3-gallate, was associated with a reduction in MMP-2 activity. Takada et al. 36 also showed that indole-3-carbinol, a phytochemical agent occurring naturally in cruciferous vegetables, inhibited cell proliferation and invasion of leukemic cells through inhibiting MMP-9 activity. These results may explain why the expression of MMP-2 and -9 is elevated in parametrial invasion. MMP activity in vivo is tightly controlled by transcriptional activation, by a complex proteolytic activation cascade, and by an endogenous system of TIMPs. TIMPs inhibit the MMPs by forming 1:1 stoichiometric complexes to regulate matrix turnover. 37,38 Here, EHF induced a marked inhibition of MMP-2 and -9 mRNA, as well as elevated the protein levels and activities of TIMP1- and -2 mRNA (Figs. 3 and 4). These data indicate that the anti-invasive activity of HEF in Hep3B cells was associated with inhibition of MMP-2 and -9 activities through the elevation of TIMP expression and suggest that the increased TIMPs/MMPs ratio is a key factor in regulating the antimetastatic process.
TJs include 24 members of the claudin family. 14,15 The claudins are transmembrane proteins with extracellular domains that interact with other claudin proteins on adjacent cells to regulate paracellular permeability. 20 TJ disruption and dysregulation of TJ protein are early events in cancer cell invasion and metastasis. For example, claudin-1 and claudin-7 are down-regulated in invasive ductal carcinomas of the breast, as well as in most established breast cancer cell lines. 39 Conversely, claudin-3 and -4 are overexpressed in breast cancers 10,39,40 and other cancers, including gastric, 41 ovarian, 42 and pancreatic 43 cancer. The nature of the dysregulation is highly cancer type specific. Recently, Miyamori et al. 24 reported that claudin promotes activation of pro-MMP-2 mediated by membrane-type MMPs. Ip et al. 23 also showed that down-regulation of claudin-10 reduced MMP activity in Hep3B cells. Additionally, Agarwal et al. 16 reported that the overexpression of claudin-3 and -4 proteins is associated with increased MMP-2 activity. These reports imply a close relationship among MMP activity, overexpression of claudin, and metastasis of cancer cells. Our results showed that EHF treatment markedly down-regulated claudin levels (-1, -3 and -4) (Fig. 5), which is associated with the tightening of TJs and decreased MMP activity, indicating that the down-regulation of claudin expression by EHF relates to increased MMP activity.
We also measured the effects of EHF on the metastasis and TJ proteins, E-cadherin, IGF-1R, and TSP-1. E-cadherin is a cell–cell adhesion molecule as well as a TJ regulator that is expressed on the membranes of epithelial cells, and low expression is associated with cancer invasion and metastasis. 25 IGF-1R is highly expressed in many types of human cancer and promotes tumor invasion and metastasis in vivo. 17,18 The expression of TSP-1 is decreased in tumor cells, which suppresses tumor growth, and high TSP-1 expression is inversely correlated with invasiveness and lymph node metastasis. 26,27 Although the regulation of these genes is closely related to metastatic and invasive activity, less is known about their impact on TJ complexes. In this study, EHF time-dependently increased levels of E-cadherin and TSP-1 but decreased IGF-1R (Fig. 5), indicating a potential mechanism for changing TER values and TJ permeability.
In summary, the present results revealed that EHF inhibits cell invasion and motility in Hep3B cells through influencing MMP activity and the expression of several important tumor invasiveness and metastasis-related genes. Our results also indicate that H. fusiforme extracts may be promising as a new dietary intervention for decreasing the risk of developing liver cancer.
Footnotes
Acknowledgments
This research was supported by grant M2007-03) from the Marine Bioprocess Research Center of the Marine Bio 21 Center funded by the Ministry of Land, Transport and Maritime and Blue-Bio Industry RIC at Dong-Eui University as an RIC (08-06-07) program of KIAT under the Ministry of Knowledge Economy, Republic of Korea.
Author Disclosure Statement
No competing interests exist.