
Abstract
In this study, hydrolysates obtained from the freshwater rotifer Brachionus calyciflonus were investigated for angiotensin I converting enzyme (ACE) inhibitory peptides. Freshwater rotifer protein was hydrolyzed using six separate enzymes in a batch reactor. The peptic hydrolysate had the highest ACE inhibitory activity compared to the other hydrolysates. The highest ACE inhibitory peptide was separated using Sephadex G-25 column chromatography and high-performance liquid chromatography on a C18 column. The 50% inhibitory concentration (IC50) value of purified ACE inhibitory peptide was 40.01 μg/mL. ACE inhibitory peptide was identified as being seven amino acid residues of Ala-Gln-Gly-Glu-Arg-His-Arg by N-terminal amino acid sequence analysis. The IC50 value of purified ACE inhibitory peptide was 47.1 μM, and Lineweaver-Burk plots suggested that the peptide purified from rotifer protein acts as a competitive inhibitor against ACE. The results of this study suggest that peptides derived from freshwater rotifers may be beneficial as antihypertension compounds in functional foods or as pharmaceuticals.
Introduction
A
Bioactive peptides are often inactive within the sequence of the parent protein and can be released by enzymatic hydrolysis. Powerful ACE inhibitory hydrolysates were obtained from whey protein after hydrolysis with trypsin. 6,7 However, it has also been suggested that hydrolysates with high levels of ACE inhibition can be obtained from caseins hydrolyzed by thermolysin, proteinase K, and pepsin, and thermolysin, proteinase K, and pepsin are more efficient than trypsin in releasing ACE inhibitory peptides from casein substrates. 8 More recently, the use of thermolysin in the study of ACE inhibitors from enzymatic hydrolysates has been reported. 9 Choosing the proper enzyme to hydrolyze different proteins is an important factor in obtaining peptides exhibiting high levels of ACE inhibition. Since the discovery of ACE inhibitors in snake venom, 10 ACE inhibitory peptides have been isolated from enzymatic hydrolysates of various marine organisms and fish waste, such as sea bream scales, 11 yellowfin sole frame protein, 12 freshwater clam, 13 oyster proteins, 14 shark meat, 15 and algae waste. 16
Freshwater rotifers are generally very small in size (100–230 μm) and exhibit rapid growth, which makes them good candidates for high-density culture. As such, freshwater rotifers are the most commonly used freshwater live feed for fish larvae cultures. 17 To date, studies on bioactive materials from freshwater rotifers remain limited. In particular, the antihypertensive effects of freshwater rotifers are yet to be investigated. The aim of this study was to purify and characterize an ACE inhibitory peptide derived from enzymatic hydrolysates of freshwater rotifer proteins.
Materials and Methods
Materials
The freshwater rotifer Brachionus calyciflonus was cultured under controlled conditions of water temperature, feed frequency, pH, and dissolved oxygen according to the procedure of Park et al. 18 Fully grown freshwater rotifers were harvested and lyophilized at −80°C using a freeze-dryer. Lyophilized freshwater rotifer powder was stored at −80°C until use. Alcalase® and Neutrase® were purchased from Novozyme Co. (Bagsvaerd, Denmark), whereas α-chymotrypsin, trypsin, papain, ACE (lung acetone powder from rabbit), substrate (Hip-His-Leu), and Sephadex G-25 were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Pepsin was purchased from Junsei (Tokyo, Japan). All other reagents were of the highest grade commercially available.
Proximate analysis
Proximate compositions of freshwater rotifers were analyzed in duplicate for moisture, crude protein, lipid, and ash according by methods of the Association of Official Analytical Chemists. 19 Moisture was determined by the oven-drying method at 105 ± 1°C for 24 hours. Crude protein was determined by the Kjeldahl method (B-324/435/412 Auto Kjeldahl System, Buchi, Flawil, Switzerland) with diethyl ether solvent. Total nitrogen content was analyzed following the Kjeldahl procedure (Kjeltec™ 1035, Foss, Höganäs, Sweden). Crude protein content was calculated using a conversion factor of 6.25. Ash was determined using a muffle furnace (model F6000, Barnstead-Thermolyne Co., Dubuque, IA, USA) at 550°C for 4 hours. Amino acid composition of the experimental diet was analyzed using an automatic analyzer (Biochrom 20, Li+-type high-performance ultra pack, Pharmacia, Milton Keynes, UK).
Enzymatic hydrolysis of freshwater rotifer protein
Harvested freshwater rotifers were hydrolyzed using various enzymes (Alcalase, α-chymotrypsin, Neutrase, papain, pepsin, and trypsin) at their optimal conditions. Before hydrolysis, specimens were subjected to ultrasonication in order to break down freshwater rotifer tissue. Subsequently, 1 g of substrate (on the basis of protein weight) and 50 mg of enzyme were mixed in a 100-mL flask under controlled temperature and pH conditions. The mixture was incubated for 12 hours at each optimal temperature with shaking and then heated in a boiling water bath for 5 minutes to inactivate the enzyme. The degree of hydrolysis was determined following the methodology of Kim et al., 20 while all freshwater rotifer hydrolysates were lyophilized at −80°C in a freeze-dryer.
Measurement of ACE inhibitory activity
ACE inhibitory activity was measured according to the methods of Cushman and Cheung, 21 with slight modification. A 50-μL freshwater rotifer hydrolysate solution with 50 μL of ACE solution (25 mU/mL) was preincubated at 37°C for 10 minutes, and then the mixture was subsequently incubated with 100 μL of substrate (8.3 mM Hip-His-Leu in 50 mM sodium borate buffer, pH 8.3) for 60 minutes at the same temperature. The reaction was terminated with addition of 250 μL of 1 M HCl. The resulting hippuric acid was extracted with 0.5 mL of ethyl acetate. After centrifugation (1,200 g, 10 minutes), 0.2 mL of the upper layer was transferred into a test tube and dried at 80°C for 1 hour. The hippuric acid was dissolved in 0.5 mL of distilled water, and absorbance was measured at 228 nm using a UV-spectrophotometer (JASCO, Tokyo). ACE inhibitory activity was calculated as follows: ACE inhibitory activity (%) = ([E c – E s])/[E c – E b]) × 100, where E c is the absorbance of the control, E s is the absorbance of the sample, and E b is the absorbance of the blank. The 50% inhibitory concentration (IC50) was defined as the concentration required to inhibit 50% of ACE inhibitory activity.
Purification of ACE inhibitory peptide
Hydrolysates with ACE inhibitory activity were dissolved in distilled water and loaded onto a Sephadex G-25 gel filtration column (2.5 × 70 cm) that had previously been equilibrated with distilled water. The column was then eluted with distilled water at a flow rate of 1.5 mL/minute. Fractions showing ACE inhibitory activity were lyophilized, and their ACE inhibitory activities were measured. The active fraction was dissolved in distilled water and separated by reversed-phase high-performance liquid chromatography (HPLC) on a Grom-sil 120 ODS-5 ST column (particle size, 5 μm; 10 × 250 mm) using a linear gradient of acetonitrile (0–50% [vol/vol], 60 minutes) containing 0.1% trifluoroacetic acid at a flow rate of 2.0 mL/minute. Elution peaks were monitored at 280 nm and purified by rechromatography by HPLC on a C18 column (particle size, 5 μm; 4.6 × 10.0 mm) using isocratic elution with 20% acetonitrile at a flow rate of 0.5 mL/minute for 30 minutes. The purified fraction with ACE inhibitory activity was collected and lyophilized, and its amino acid sequence was identified.
Determination of amino acid sequence
Amino acid analysis was carried out with purified active peptide, which was used for N-terminal amino acid sequencing at the Korea Basic Science Institute (Daejeon, Republic of Korea). The purified activity peptide was excised from the two-dimensional gels and subjected to in-gel digestion. 22 The trypsin- or pepsin-digested peptide mixtures were analyzed with a model 4700 Proteomics Discovery System (PerSeptive Biosystems, Framingham, MA, USA) and with Ultraflex MALDI TOF/TOF mass spectrometer (Bruker Daltonics GmbH, Bremen, Germany). Peptide mass finger printings were acquired by summing 750 laser shots. Monoisotopic peptide masses were selected in the range of 800–3,000 Da, and the proteins were identified using the Matrix Science Mascot and National Center for Biotechnology Information protein sequences database. 23
Determination of ACE inhibition pattern
Various concentrations of purified ACE inhibitory peptide were measured for inhibition pattern according to the method of Kim et al., 20 with some modifications. Enzyme activity was measured with different concentrations of substrate. The ACE inhibitory pattern was determined in the presence of the inhibitor with Lineweaver-Burk plots.
Statistical analysis
All results were expressed as mean ± SEM values (n = 3).
Results
Proximate compositions
The results of proximate composition analysis for crude protein, lipid, carbohydrate, and ash content were 63.72%, 13.13%, 7.09%, and 3.68%, respectively (data not shown). The crude protein content was higher than the other compositions. The amino acid composition of the freshwater rotifer is shown in Table 1. The major amino acids were Gln (11.82%), Asp (10.53%), Leu (10.28%), and Gly (8.98%). Gln, Asp, Leu, and Gly constituted the majority of the total amino acid composition.
Preparation of hydrolysates and their ACE inhibitory activity
Various ACE inhibitory peptides have been purified from enzymatic hydrolysates of marine organisms. However, freshwater zooplankton remained unused, until now. In this study, freshwater rotifer hydrolysates were prepared by means of hydrolysis for the extraction of ACE inhibitory peptides using commercial proteases, including Alcalase, α-chymotrypsin, Neutrase, papain, pepsin, and trypsin. The degree of hydrolysis was observed to be 70.05%, 65.41%, 63.52%, and 56.31% for trypsin, pepsin, α-chymotrypsin, and papain, respectively (data not shown). In ACE inhibitory activation (Table 2), the highest IC50 value was exhibited by the peptic hydrolysate at 0.57 mg/mL.
Purification of ACE inhibitory peptide
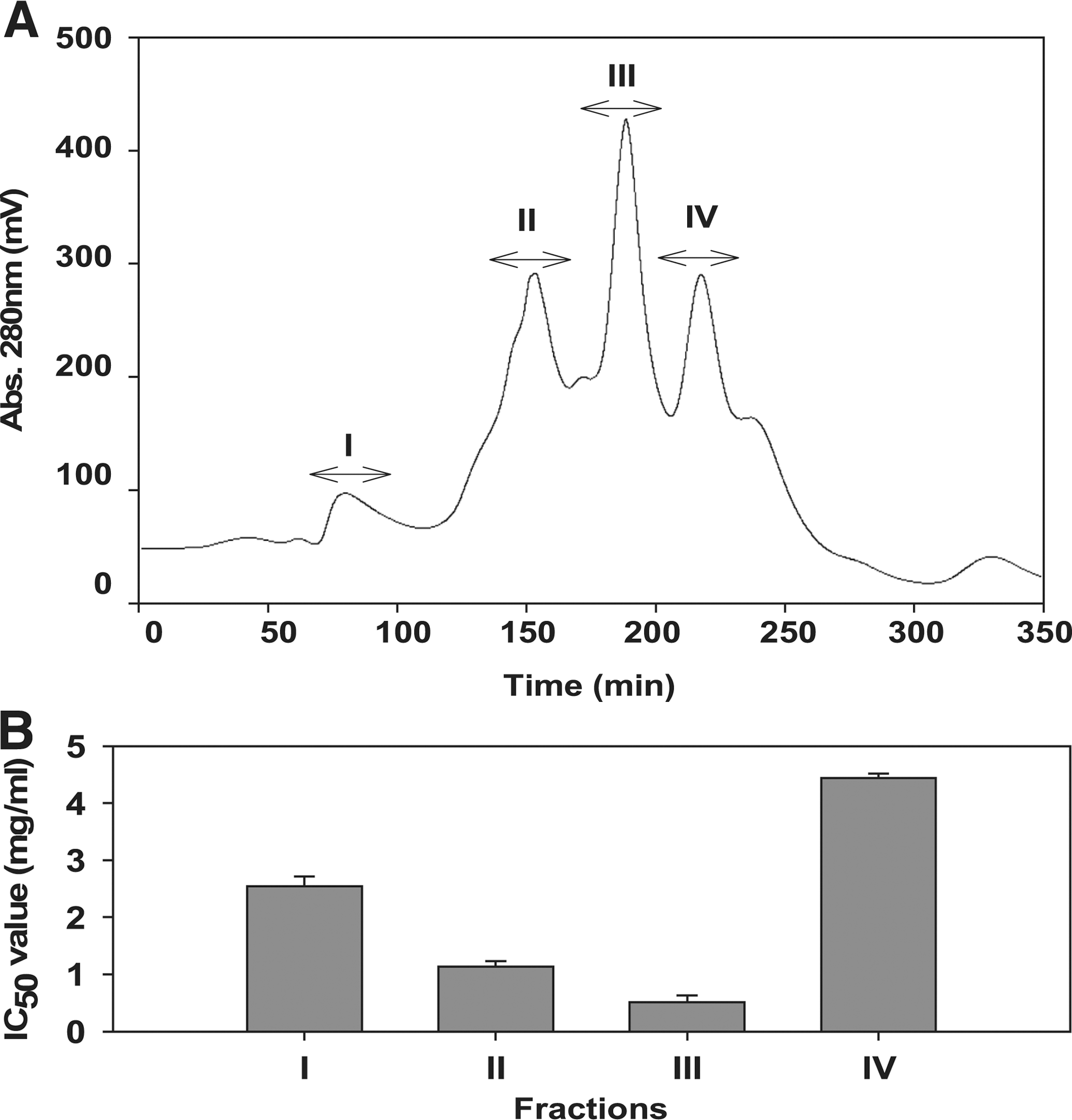
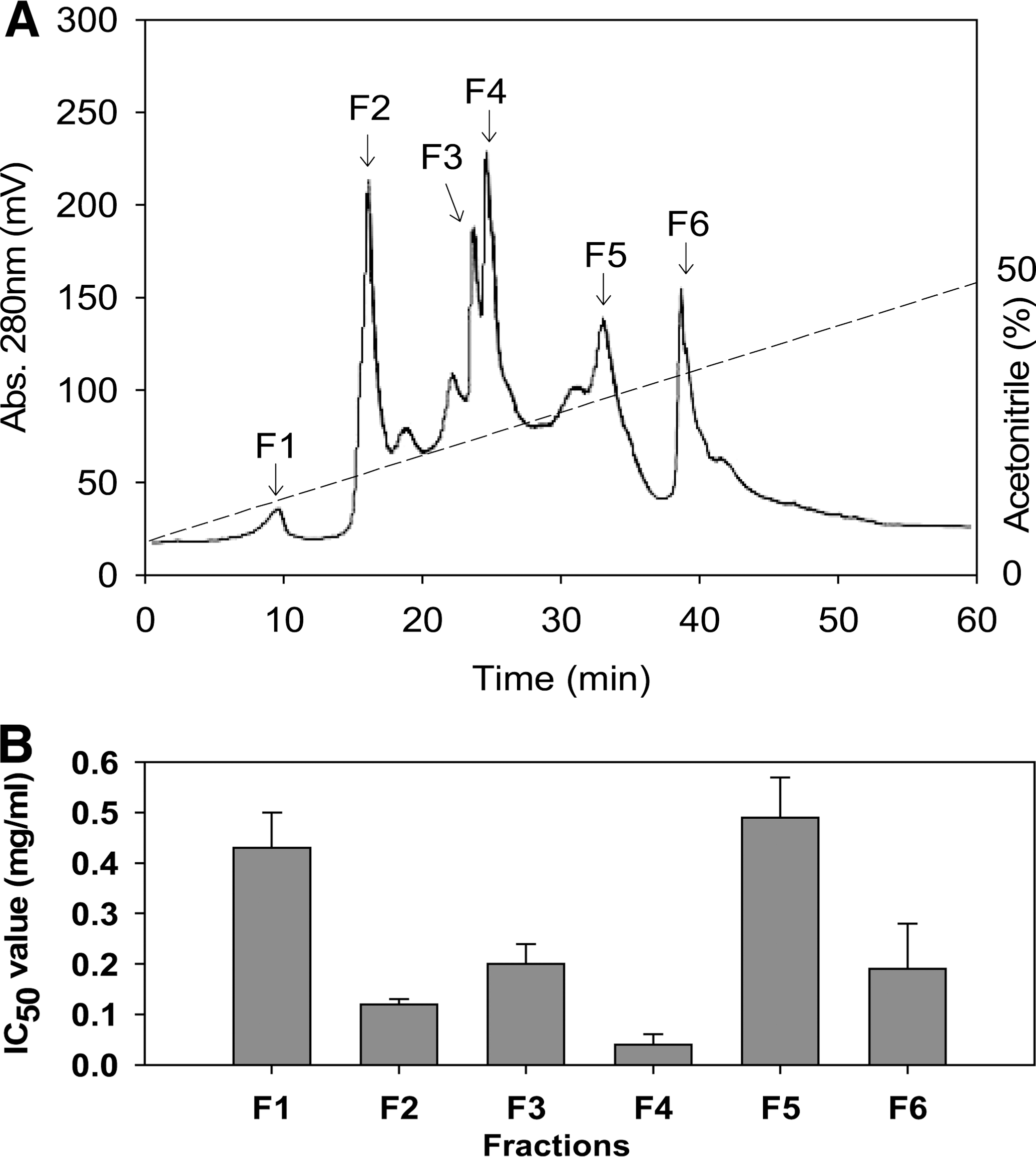
To assess the ACE inhibitory activity of the freshwater rotifer hydrolysates, purification of peptides from hydrolysates was undertaken using Sephadex G-25 column chromatography. Fractions of 7.5 mL were collected and pooled in order to obtain fractions I–IV as shown in Figure 1. Fraction III had the highest ACE inhibitory activity with an IC50 value of 0.51 mg/mL. Based on activity, fractions III and IV appear to contain the most potentially useful peptides. Fraction III was still a complex mixture of peptides, which was separated according to the hydrophobicity of the peptides. Fraction III was applied to a reversed-phase HPLC system using a Grom-sil 120 ODS-5 ST column with gradient elution by acetonitrile (0–50%). As can be seen from the chromatographic evidence (Fig. 2A), a total of six fractions were collected, and ACE inhibitory activity was determined. The peptide concentration of each fraction was adjusted to a predetermined IC50 value of ACE inhibitory activity (Fig. 2B), so that the fraction with the lowest peptide content had the highest activity. F4 showed the most potent ACE inhibitory activity with an IC50 value of 40.01 μg/mL. Finally, purified fraction F4 had the highest ACE inhibitory activity. The active fraction F4 was subjected to rechromatography on the HPLC column using a linear isocratic elution of 20% acetonitrile for 30 minutes at a flow rate of 0.5 mL/minute. The ACE inhibitory peptide was purified at 14.2-fold from hydrolysate (0.57 mg/mL) using a triplicate purification procedure.
(
(
Amino acid sequences of purified peptide
Amino acid sequences of purified ACE inhibitory peptides were identified using N-terminal amino acid sequencing and shown to be Ala-Gln-Gly-Glu-Arg-His-Arg for F4 with ACE inhibitory IC50 values of 47.1 μM and a molecular mass of 848 Da (Fig. 3). The amino acids composing peptides with ACE inhibitory activity are critical factors in whether a peptide possesses ACE inhibitory activity.
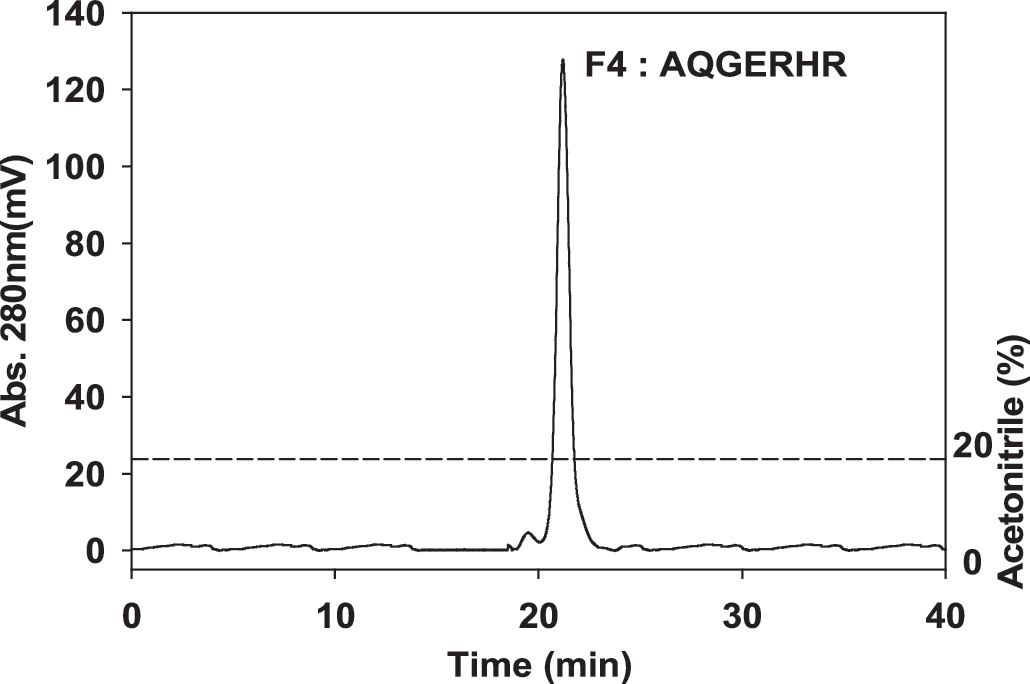
The reverse-phase HPLC pattern of active fraction F4 on a C18 column (particle size, 5 μm; 4.6 × 250 mm) eluted at a flow rate of 0.5 mL/minute.
ACE inhibition pattern of purified peptide
ACE inhibition patterns for the reaction at different concentrations of the purified peptide were determined according to Lineweaver-Burk plots. The inhibition pattern was determined to be competitive (Fig. 4). This indicates that the peptide could bind competitively with the substrate at the active site of ACE.
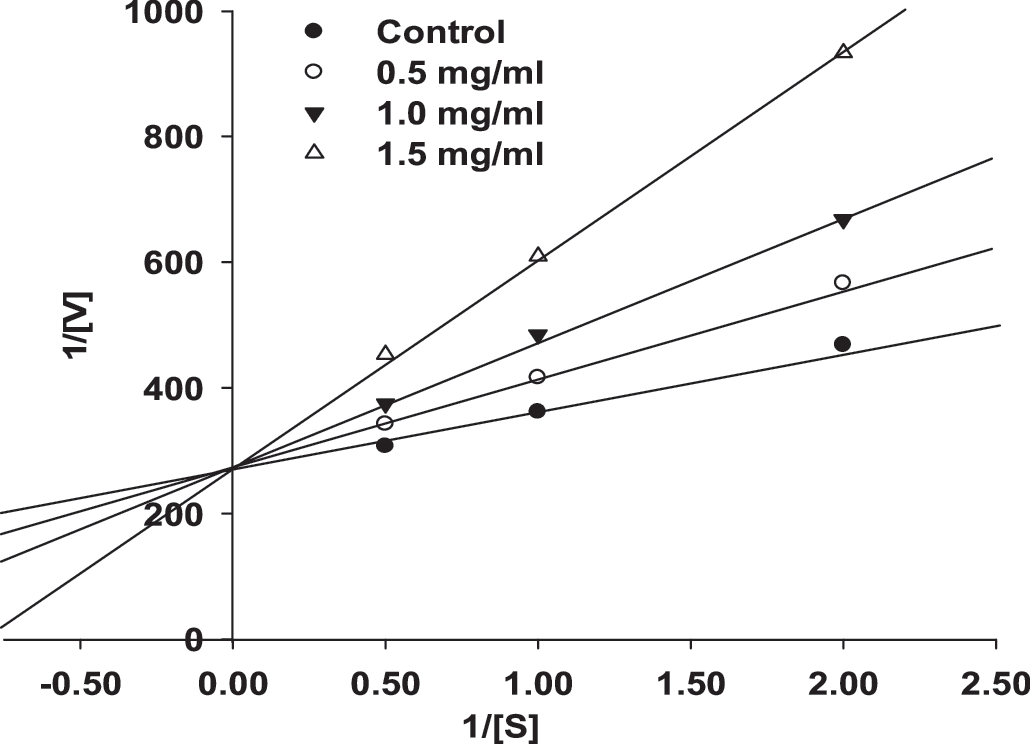
Lineweaver-Burk plots of ACE inhibition by the purified peptide from freshwater rotifer protein.
Discussion
ACE inhibitory peptide was produced from freshwater rotifer protein using enzymatic hydrolysis. Enzymatic hydrolysis is one of the approaches for the effective release of bioactive peptides from protein sources. It is widely applied to improve and upgrade the functional and nutritional properties of proteins. In the present study, various hydrolysates were produced by different proteases, and their ACE inhibitory activities were evaluated. The freshwater rotifer had the highest protein content of any marine organisms tested. In general, the protein content of freshwater rotifers is between 28% to 67% of dry weight. 24 Larval fish generally have a high demand for dietary protein because of their rapid growth rates and extensive catabolism of amino acids for the production of metabolic energy. 25 Table 1 shows the amino acid compositions of harvested freshwater rotifer in our laboratory. Among the amino acids, Gln, Asp, Leu, and Gly were present in the highest concentrations. Amino acid compositions of rotifers cultured with Chlorella were determined to be 9.74%, 7.79%, 2.56%, and 5.29% for Gln, Asp, Leu, and Gly, respectively; these values for Leu and Gly appeared to be somewhat different from other reports. 26
The degree of hydrolysis after proteolytic digestions was observed to be 70.05%, 65.41%, and 60.51% for trypsin, α-chymotrypsin, and pepsin, respectively. In algae waste hydrolysates, the degree of peptic hydrolysate has been reported to be 30%, 16 whereas pepsin specificity hydrolyzes the front of hydrophobic amino acids such as Phe, Met, Leu, and Trp. 8
Peptides from six hydrolysates were evaluated for their ACE inhibitory activity; the highest IC50 value (Table 2) was exhibited by the peptic hydrolysate at 0.57 mg/mL, whereas that of bullfrog peptic hydrolysate was 0.33 mg/mL, 27 and peptic hydrolysate showed the highest ACE inhibitory activity from algae waste hydrolysate. 16 Considering the ACE inhibitory activity of peptides from enzymatic hydrolysates, the unique composition and sequence of amino acid residues may play a vital role in ACE inhibitory activity of the purified peptides. Furthermore, ACE inhibitory activity is also limited by the structure of the polypeptide, and the amino acid sequence of the peptide is also important. 28
Purifying the ACE inhibitory peptide from peptic hydrolysate of freshwater rotifer required the use of different chromatographic techniques. The peptic hydrolysate was separated into five fractions on a Sephadex G-25 column (Fig. 1). Sephadex G-25 column chromatography separates according to molecular size, where FI and FII represented large-molecular-size peptides, and fractions FIII–FV contained small-molecular-size peptides. According to the report of Pihlanto-Leppala, 29 most bioactive peptides contain two to 20 amino acids and exhibit a small molecular size. As such, fractions III and IV appeared to have the most potential to contain bioactive peptides. Fraction III was still a complex mixture of peptides, which was separated according to the hydrophobicity of the peptides.
Following different chromatographic methods, we purified the peptide with the most potent ACE inhibitory activity, which had the sequence Ala-Gln-Gly-Glu-Arg-His-Arg (molecular mass, 848 Da) (Fig. 3). We reported that Gln, Asn, Pro, Gly, and Ala are common amino acids among ACE inhibitory peptides: for example, peptic hydrolysate of the microalgae Spirulina platensis, with peptides having IC50 values for Ile-Ala-Glu, Ile-Ala-Pro-Gly, and Val-Ala-Phe of 34.7, 11.4, and 35.8 μM, respectively 30 ; freshwater clam (Corbicula fluminea, Muller) with IC50 values for Val-Lys-Pro and Val-Lys-Lys of 3.7 and 1,045 μM, respectively 13 ; algae waste protein with an IC50 value for Val-Glu-Cys-Tyr-Gly-Pro-Asn-Arg-Pro-Gln-Phe of 29.6 μM 16 ; sea bream scales hydrolysate with an IC50 value for Val-Ile-Tyr of 7.5 μM 12 ; and pearl oyster with an IC50 value for Leu-Val-Glu of 14.2 μM. 14 With regard to the relationship between peptide structure and ACE inhibition, Cheung et al. 31 reported that those peptides that contained Pro, Phe, or Tyr at the C-terminal and those that contained the amino acids Val and Ile at the N-terminal had the highest ACE inhibitory activity. It has been confirmed that functional peptides are dependent on amino acid sequence and structure. 28 Thus, the sequencing and structure of peptides might be related to ACE inhibitory activity. Similarly, other structure–activity correlation studies have indicated that ACE binding is strongly affected by the C-terminal tripeptide sequence of the substrate and that the tripeptide could interact with subsites S1, S'1, and S'2 of ACE. 32 The amino sequencing strongly affects potential ACE inhibition because of the inclusion of hydrophobic amino acid residues (aromatic or branched side chains) at the C-terminal. 33,34 Hydrophobic amino acid residues in the ACE inhibitor sequence are a critical factor in inhibitory activity. 34 Therefore, we concluded that the purified peptide exhibited low ACE inhibition activity because of nondistribution of hydrophobic amino acids at the C-terminal.
Enzymes catalyze reactions in physiological systems. In equilibrium, an enzyme binds a substrate to form an enzyme-substrate complex. Enzyme inhibition is a common goal for the pharmaceutical industry. All inhibitors cause the substrate to react at a lower rate than without the inhibitor. Reversible enzyme inhibitors fall into three categories: competitive, noncompetitive, and uncompetitive. Furthermore, noncompetitive inhibitors can be divided into two additional categories: pure and mixed. The Lineweaver-Burk equation can be used to categorize different types of inhibitors. Understanding the type of inhibitor will give clues on how its structure might be modified to increase its potency. This indicates that the peptide could bind competitively with the substrate at the active site of ACE. Purified peptides have been reported to exhibit competitive inhibition of ACE (Fig. 4), as shown by captopril. 13 Our result was similar to those with the peptide extracted from freshwater clam (C. fluminea, Muller), 13 oyster sauce (Crassostrea gigas), 35 and the soy fermented product natto. 36 Although most reported peptide inhibitors of ACE acted as competitive inhibitors, a few have exhibited noncompetitive-type ACE inhibition. 13 However, the inhibition site of the competitive inhibitor of ACE has not been identified, and the exact inhibition mechanism of ACE inhibitory peptide is yet to be clearly revealed. 37
Freshwater rotifers are the most commonly used live feed in larval aquaculture. In this study, the ACE inhibitory activity of peptide purified from freshwater rotifer peptic hydrolysate was assessed. Based on the results of this study, it appears that this peptide may be beneficial as a bioactive material. However, further investigation is needed in order to confirm its in vivo physiological effects in blood pressure regulation.
In conclusion, antihypertensive effects of various enzymatic hydrolysates prepared from the freshwater rotifer B. calyciflonus were evaluated by ACE inhibitory activity measurement. Among the six hydrolysates, peptic hydrolysate showed the highest ACE inhibitory activity. These results indicate that purified ACE inhibitory peptide from peptic hydrolysate possesses a potent competitive inhibition pattern. Based on the results of this study, it appears that this peptide may be beneficial as a bioactive material and functional food resource.
Footnotes
Acknowledgments
This research was supported by a grant from Marine Bioprocess Research Center of the Marine Biotechnology funded by the Ministry of Land, Transport and Maritime, Republic of Korea.
Author Disclosure Statement
M.-S.L. is an employee of Nonione Co. J.K.L., H.G.P., S.-K.K., and H.-G.B. declare no competing financial interests exist.