
Abstract
The objective of this study was to examine the extent to which bitter melon seed (BMS) alleviates the symptoms associated with metabolic syndrome and elucidate the mechanism by which BMS exerts beneficial effects. Three-month-old female Zucker rats were assigned to following groups: lean control (L-Ctrl), obese control (O-Ctrl), and obese + BMS (O-BMS). The control groups were fed AIN-93M purified rodent diet, and the O-BMS group was fed AIN-93M diet modified to contain 3.0% (wt/wt) ground BMS for 100 days. After 100 days of treatment, BMS supplementation in the obese rats lowered the total serum cholesterol by 38% and low-density lipoprotein-cholesterol levels by about 52% and increased the ratio of serum high-density lipoprotein-cholesterol to total cholesterol compared to the O-Ctrl group. The percentage of total liver lipids was about 32% lower and serum triglyceride levels were 71% higher in the O-BMS group compared to the O-Ctrl group. Serum glucose levels were significantly lowered partly because of the increase in the serum insulin levels in the BMS-based diet groups. BMS supplementation increased the expression of peroxisome proliferator-activated receptor-γ (PPAR-γ) in the white adipose tissue of the obese rats significantly (P < .05) and down-regulated the expression of PPAR-γ, nuclear factor-κB (NF-κB), and interferon-γ mRNA in heart tissue of the obese rats. The findings of this study suggest that BMS improves the serum and liver lipid profiles and serum glucose levels by modulating PPAR-γ gene expression. To our knowledge, this study for the first time shows that BMS exerts cardioprotective effects by down-regulating the NF-κB inflammatory pathway.
Introduction
In addition to increasing physical activity, one of the aspects of lifestyle modification for preventing metabolic syndrome is changes in diet. Foods rich in phytochemicals that possess health benefits such as hypoglycemic and hypolipidemic effects have become a central focus for research to prevent risk factors associated with metabolic syndrome. Momordica charantia, commonly known as bitter gourd or bitter melon, a commonly consumed vegetable in Asia, has been recognized for its medicinal value. The most noteworthy health benefit of the juice or whole powder of bitter gourd fruit is the hypoglycemic potential both in diabetic rat models and in human subjects with type 2 diabetes. 3 –6 The hypoglycemic activity of M. charantia was attributed to a few isolated phytochemicals such as charantin, polypeptide-p, momordin Ic, oleanolic acid 3-O-monodesmoside, and oleanolic acid 3-O-glucuronide. 7 –9 In addition, M. charantia has been shown to significantly reduce serum cholesterol and triglycerides in normal rats as well as diabetic rats. 10 –13 Dietary bitter melon was shown to suppress inflammatory responses by enhancing the humoral immunity. 14 However, there is a relative paucity of data regarding the molecular mechanisms through which bitter melon exerts its hypoglycemic and hypolipidemic activity.
In recent years numerous efforts have been directed toward understanding the molecular mechanisms that govern adipogenesis. Peroxisome proliferator-activated receptor (PPAR)-γ, a nuclear receptor, is highly expressed in adipose tissue and now recognized to be a master regulator of adipogenesis. 15 Exposure of cultured primary human preadipocytes to PPAR-γ agonists such as thiazolidinediones induces their differentiation. 16 PPAR-γ agonists improve insulin sensitivity by promoting adipogenesis and postprandial fatty acid/triglyceride storage within adipocytes. 16 In addition to PPAR-γ, the nuclear factor-κB (NF-κB) signaling pathway has received much attention because of its role in several pathological processes in the myocardium, including cardiomyocyte pro-inflammatory cytokine release, ischemia/reperfusion injury, hypertrophy, and apoptosis. 17
Because bitter melon is known for its anti-inflammatory and anti-hyperglycemic activity, we hypothesized that bitter melon seed (BMS) might interfere with PPAR-γ and NF-κB signaling pathway to lower cardiovascular disease risk factors. In the present study, we have investigated the extent to which BMS alleviated the symptoms associated with metabolic syndrome and modulated the associated molecular mechanisms in the obese (ff a/fa) Zucker rat, a model of metabolic syndrome.
Materials and Methods
Study design
Thirty 3-month-old female Zucker rats (Harlan Sprague-Dawley, Indianapolis, IN, USA) were housed in an environmentally controlled animal care facility for the duration of the experiment. University of Arkansas (Fayetteville, AR, USA) Institutional Animal Care and Use Committee guidelines for the treatment and care of the animals were followed throughout the study duration. After 1 week of acclimatization, the animals were divided into the following treatment groups: lean + control (L-Ctrl), obese + control (O-Ctrl), and obese + BMS (O-BMS). The rats in the lean and obese control groups received AIN-93M rodent diet, and the rats in treatment group received AIN-93M diet modified to contain freeze-dried, powdered BMS at 3% (wt/wt) concentration. M. charantia L. (Indian green variety) used in the study was grown at the University of Arkansas farm. The composition of the diet is given in Table 1. The moisture, protein, and lipid content of the BMS has been published elsewhere. 18 The BMS diet had similar levels of carbohydrate, fiber, fat, and protein following modification of the AIN-93M formulation based on proximate analysis of BMS powder. Rats were pair-fed to the mean food intake of the lean group and had free access to deionized water. The weight of the animals was measured once weekly, and their food intake was monitored three times a week.
BMS, bitter melon seed.
Body composition
Dual-energy X-ray absorptiometry (GE Lunar DXA, Waukesha, WI, USA) was used to analyze the body composition of the rats before and after the study. The animals were anesthetized and placed stomach down on the scan bed of the dual-energy X-ray absorptiometry instrument. The absorbance of two X-ray beams is measured, and percentage of lean tissue and percentage of fat tissue are calculated by the software appropriate for the body composition assessment of small animals.
Animal necropsy and sample processing
After 100 days of treatment, the rats were fasted for 12 hours before being sacrificed by exsanguination via cardiac puncture. Blood was centrifuged at 3,000 g for 20 minutes at 4°C to separate serum from whole blood. Aliquots of serum were stored at −80°C until time for analysis. Organs of interest were removed, placed in cryogenic storage containers, flash-frozen in liquid nitrogen, and stored at −80°C.
Serum glucose and lipid profiles
Serum triglycerides, total cholesterol, high-density lipoprotein-cholesterol (HDL-C), low-density lipoprotein-cholesterol (LDL-C), and glucose concentrations were determined using commercially available kits from Alfa Wassermann Diagnostic Technologies (West Caldwell, NJ, USA). An ACE Alera® clinical chemistry system (Alfa Wassermann Diagnostic Technologies) was used according to the manufacturer's instructions to perform these tests.
Liver total lipids and cholesterol
One gram of liver was homogenized in a 20-fold volume of 2:1 (vol/vol) chloroform-methanol mixture. Following homogenization, 0.58% NaCl solution was added to achieve separation of the phases and centrifuged for 20 minutes at 500 g. Supernatant was discarded, and the organic phase was filtered and washed with chloroform through fat-free filter paper (Whatman International Ltd., Maidstone, United Kingdom). The filtered organic phase containing the tissue lipids was then transferred to a preweighed scintillation vial. Liver lipids were determined using the gravimetric method of Folch et al. 19 Liver total cholesterol was determined using the method described by Searcy and Bergquist. 20
Insulin
Serum insulin was quantified using a commercially available enzyme-linked immunosorbent assay (Alpco Immunoassays, Salem, NH, USA). Serum was thawed on ice before analysis. A 96-well plate in the kit was prepared following the manufacturer's instructions to include duplicate standards, appropriate controls, and sample duplicates. Absorbance was measured using a BioTek (Winooski, VT, USA) ELx808 microplate reader attached to a PC running BioTek Gen5 data analysis software.
RNA extraction and cDNA synthesis
RNA was extracted from approximately 100 mg of white adipose tissue by the TRIzol® (Invitrogen, Carlsbad, CA, USA) reagent method using the RNeasy Lipid Tissue Mini Kit (Qiagen, Valencia, CA, USA) by following the supplier's instructions. Total RNA from rat heart was extracted using the TRIzol method as per the instructions. In brief, rat heart tissues were homogenized in a ground glass homogenizer using TRIzol (1 mL/100 mg of tissue) and were transferred to a 1.5-mL microcentrifuge tube. The rest of the RNA extraction steps were followed as per the manufacturer's instructions, and the final washed pellets of RNA were dissolved in 40 μL of RNase-free water. The total amount of RNA present in each sample was quantitated using a Nanodrop (Thermo Scientific, Wilmington, DE, USA), and a 1-μg sample of total RNA was used for cDNA synthesis using the Bio-Rad (Hercules, CA, USA) iSCRIPT™ cDNS synthesis kit.
Quantitative real-time polymerase chain reactions
Following cDNA synthesis, real-time quantitative polymerase chain reactions (PCRs) were performed using the Bio-Rad iQ™ SYBR® Green PCR system. Concentrations of reagents were used per the manufacturer's instructions. Real-time PCR primers were designed using Primer3 primer design software, and all primer sets were synthesized by Invitrogen. Primer sequences used for this study are summarized in Table 2. The real-time PCR was performed on the Bio-Rad iCYCLER real-time PCR instrument. The following experimental conditions were used for all target gene expression, including generation of standard curves. The initial denaturation cycle was performed at 95°C for 5 minutes. All subsequent denaturation and annealing cycles were repeated 45 times at 95°C for 15 seconds and 60°C (55°C annealing for reference gene β-actin) for 45 seconds, respectively. The changes in gene expression ratio by real-time quantitative PCR were calculated using REST software.
IFN-γ, interferon-γ; NF-κB, nuclear factor-κB; PPAR-γ, peroxisome proliferator-activated receptor-γ.
Statistical analysis
The data analyses involved estimation of means and SEM using JMP 8 (2009) (SAS Institute Inc., Cary, NC, USA). Analysis of variance was performed to determine the statistically significant (P < .05) differences among the groups. When analysis of variance indicated any significant differences among the means, Fisher's Least Significant Difference follow-up multiple comparison test was used to determine which means were significantly different (P < .05).
Results
Food consumption and body weight and composition
The results for the average food consumption, body weight, and percentage body fat are presented in Table 3. There was no difference in the food intake between the obese groups as the animals were pair-fed to the mean food intake of the lean group. There was a significant difference in the final body weight among the lean and the obese groups (P < .05). BMS supplementation did not alter the body weight of the obese rats. There was no significant difference between the final body weights of the O-Ctrl and O-BMS groups. The percentage body fat was significantly lower in the L-Ctrl group compared to the obese groups. No significant difference in percentage body fat was observed between the O-Ctrl and O-BMS groups.
Data are mean ± SEM values.
Values in a row without common superscripts are significantly different (P < .05).
Serum and liver lipids
The results for the serum and liver lipid profiles are presented in Table 4. There was a significant difference in the serum cholesterol levels among the three groups (P < .05). BMS supplementation reduced the serum cholesterol by 38% compared to the obese control. Simultaneously, the ratio of serum HDL-C to total cholesterol increased in the O-BMS group compared to the O-Ctrl group (Table 4). There was a significant difference in the serum LDL-C levels among the three groups (P < .05). BMS supplementation lowered LDL-C levels by about 52% compared to the O-Ctrl group. Serum triglyceride levels in the O-BMS group were significantly higher compared to the O-Ctrl group (P < .05). There was a significant difference in the percentage of total liver lipids among the three groups (P < .05). Compared to the O-Ctrl group, the percentage of total liver lipids was about 32% lower in the O-BMS group. However, there was no difference in the percentage of total liver cholesterol levels between the O-Ctrl and O-BMS groups.
Data are mean ± SEM values.
Values in a row without common superscripts are significantly different (P < .05).
HDL-C, high-density lipoprotein-cholesterol; LDL-C, low-density lipoprotein-cholesterol.
Serum glucose and insulin
Effects of BMS supplementation on circulating glucose and insulin concentrations are shown in Table 5. There was a significant difference in the serum glucose levels among the three groups (P < .05). BMS diet lowered the serum glucose levels in obese Zucker rats by 38% in comparison to the O-Ctrl group. This, in part, may be due to increases in the serum insulin level in the O-BMS group (97% higher compared to the O-Ctrl group).
Data are mean ± SEM values.
Values in a row without common superscripts are significantly different (P < .05).
Expression of PPAR-γ in white adipose tissue in Zucker rats
We analyzed the PPAR-γ expression in the white adipose tissue of obese Zucker rats treated with BMS by real-time PCR (Fig. 1). The results show that BMS supplementation increased the expression of PPAR-γ in the white adipose tissue of obese Zucker rats significantly compared to the lean control (P < .05).
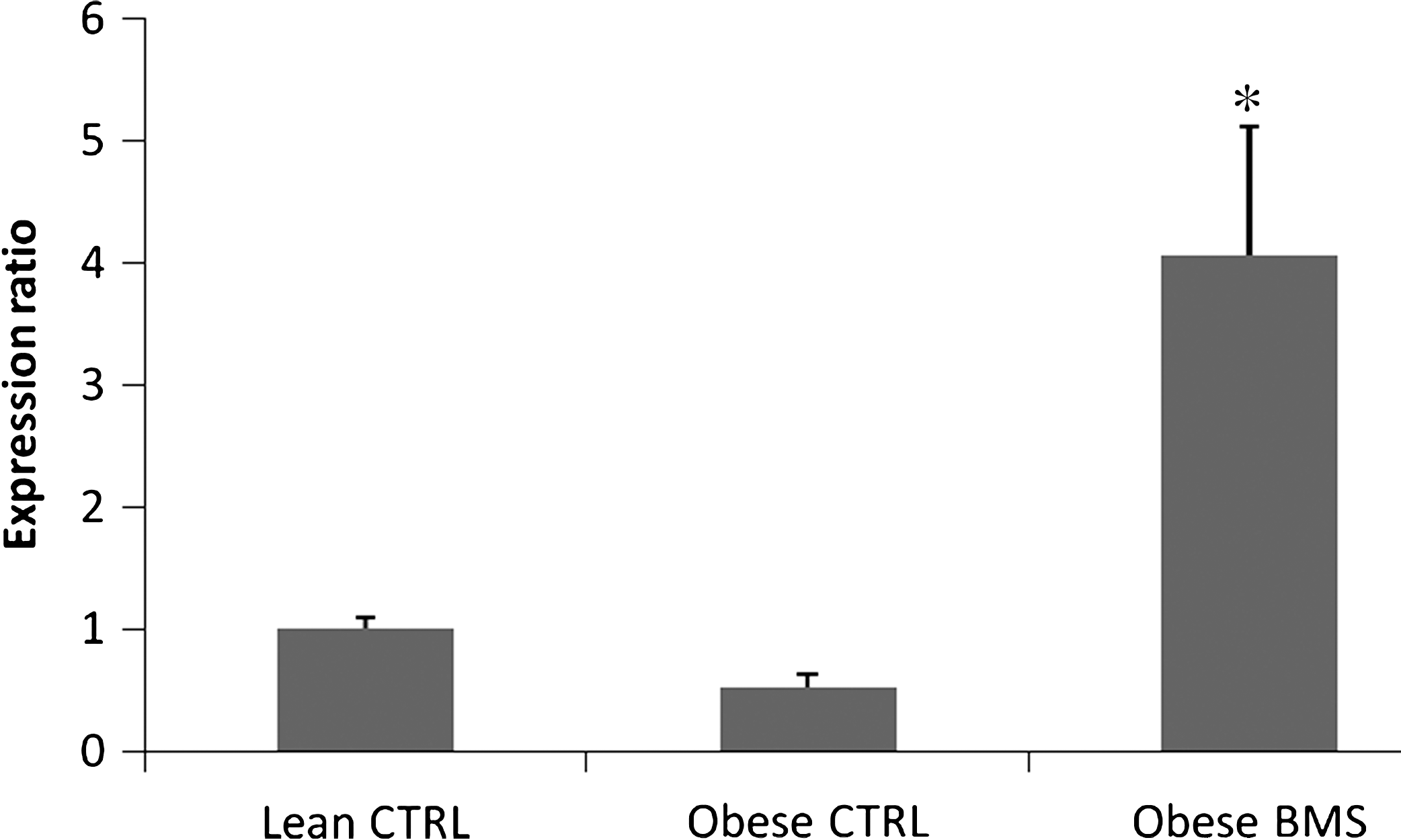
Expression of PPAR-γ in white adipose tissue from Zucker rats treated with BMS compared to the lean control (CTRL) (n = 4 in triplicates) by real-time polymerase chain reaction. *Represents significant difference (P < .05).
Expression of PPAR-γ, NF-κB, type I collagen, and interferon-γ heart mRNA in Zucker rats
To explore the potential molecular mechanism involved in lowering cardiovascular disease risk factors by BMS, expression of genes for PPAR-γ and related proteins was determined by real-time PCR (Fig. 2). PPAR-γ isozymes are a class of nuclear receptors associated with cardiovascular disease such as coronary heart disease. We studied the gene expression of PPAR-γ, NF-κB, and interferon-γ in the heart (Fig. 2). The results showed that BMS diet down-regulated the expression of PPAR-γ, NF-κB, and interferon-γ mRNA in heart tissue of the Zucker rats with the down-regulation of NF-κB being significantly lower among all the genes studied (P < .05).
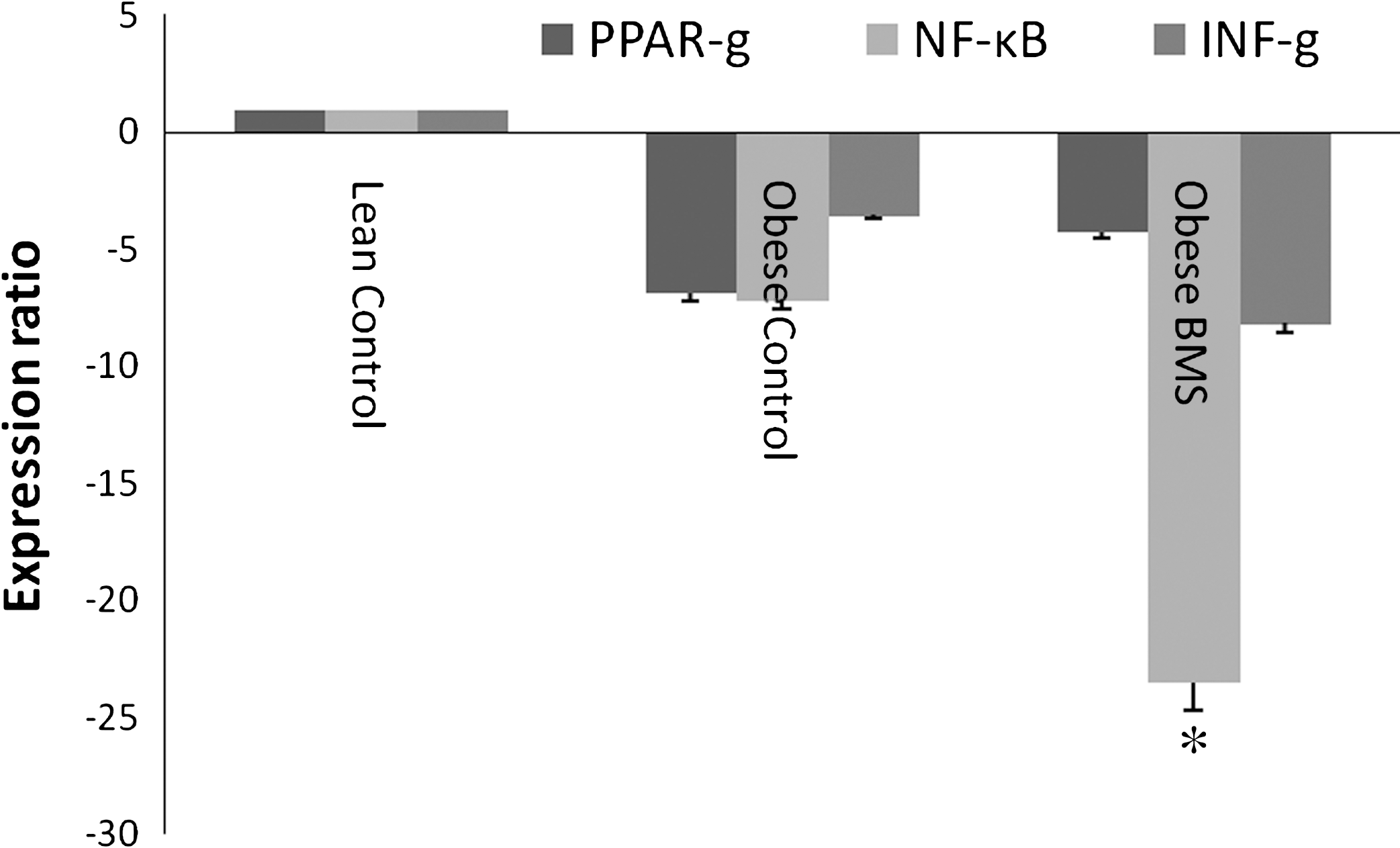
Expression of PPAR-γ, NF-κB, and IFN-γ in hearts from Zucker rats treated with BMS compared to the lean control (n = 4 in triplicates) by real-time polymerase chain reaction. *Represents significant difference (P < .05).
Discussion
An increasing number of studies indicate that dietary patterns incorporating unprocessed plant foods rich in phytochemicals play an important role in alleviating metabolic syndrome-associated abnormalities. 21 –23 In the present study the effect of BMS in reversing the hyperlipidemia, hypercholesterolemia, and hyperglycemia associated with excess body weight and the molecular mechanisms by which it exerts these positive effects were studied in obese Zucker rats. The Zucker rat is an established and widely used model of obesity. These animals are hyperlipidemic, hypercholesterolemic, hyperinsulinemic, and hyperphagic. 24 –26
BMS supplementation in obese animals did not prevent weight gain or lower the percentage body fat. Other studies using freeze-dried bitter melon fruit extract in a high fat diet showed a decrease in the mean body weight and percentage body fat. 4 The differences in the response can in part be explained due to the difference in the diet and supplementation with bitter melon fruit extract. High fat diet has been shown to increase the energy efficiency in obese Zucker rats. 27 Chen et al. 4 demonstrated that bitter melon is effective in decelerating the body weight in Zucker rats by lowering the energy efficiency. In contradiction to the study by Chen et al. 4 with bitter melon extract, BMS in this study was not effective in lowering the body weight in obese rats.
The present study demonstrated that BMS lowered serum cholesterol levels significantly. Interestingly, in this study, we have also observed that BMS supplementation increased serum HDL-C levels and lowered serum LDL-C levels significantly in the obese rats, providing evidence for the possible anti-atherogenic activity of BMS. Because the risk for coronary heart disease increases with the increase in total cholesterol and LDL-C levels and the decrease in the HDL-C levels, a reduction in the serum cholesterol and LDL-C fraction and an increase in the HDL-C fraction due to BMS supplementation should contribute to a long-term positive prognosis in obesity. In addition, a pronounced reduction in the liver lipids was also noted in the obese rats fed the BMS diet. However, the total cholesterol level in the liver was unaltered in these rats. Our findings are consistent with the results of other researchers who have demonstrated that bitter melon exerts a beneficial cholesterol-lowering effect and improves the serum lipid profile. 11,28 The hypolipidemic activity of BMS in the liver can be explained because of the increased insulin output in the obese rats. Insulin is known to activate the enzyme lipoprotein lipase, which hydrolyzes lipoprotein-bound triglycerides. 29 Furthermore, insulin reduces lipolysis and mobilization of peripheral depots by inhibiting adipose tissue hormone-sensitive lipase. 28 Bitter melon juice was shown to improve lipid metabolism in the liver by increasing phosphorylation of insulin signaling molecules or through activating AMP-activated protein kinase. 30 Other mechanisms such as increased activity of the key lipid oxidation enzyme acyl-coenzyme A dehydrogenase have also been demonstrated. 31 It is therefore reasonable to summarize that the mechanisms of action of bitter melon in improving lipid profiles are multifold.
The reasons for the BMS-associated increase in the serum triglyceride levels in our study are not apparent and might be related to the lipid content of BMS. The 50% lipid content of BMS is in the form of a conjugated linolenic acid (CLN). 32 CLN in bitter melon oil has been shown to increase the serum concentration of triglycerides and cholesterol in rats. 33 A CLN-associated increase in serum triglycerides was also reported by Koba et al. 34 This study also showed a significant decrease in serum cholesterol levels, which is in agreement with our studies. The elevated levels of triglycerides in the CLN-fed rats was hypothesized to be due to inability to clear very-low-density lipoprotein, possibly because of an inhibition of lipoprotein lipase. 34 However, this mechanism is unlikely in our study because we also observed BMS-associated induction of expression of the PPAR-γ gene in the adipose tissue (discussed below). PPAR-γ agonists are known to decrease plasma triglyceride and free fatty acids levels by enhancing their catabolism via the induction of lipoprotein lipase. 35
Among the well-documented biological activities of bitter melon, the hypoglycemic effect has received the most attention. The majority of studies conducted used extracts of bitter melon fruit pulp for hypoglycemic properties in normal and diabetic rat models. 36 In this study, we have shown that BMS powder lowers the blood glucose levels of obese rats to levels similar to that of their lean counterparts. More than one mechanism for hypoglycemic effects of bitter melon has been proposed. For instance, active insulinomimetic components from the BMS such as galactose binding lectin and p-insulin or v-insulin have been reported in earlier in vitro studies. 9,37 Other phytochemicals such as a mixture of steroidal saponins known as charantin as well as alkaloids in bitter melon have been reported to be responsible for the hypoglycemic actions. 38 A direct hypoglycemic effect of bitter melon fruit by a mechanism similar to insulin has been demonstrated by Akhtar et al. 6 Other studies have shown that bitter melon extracts stimulated insulin release from isolated pancreatic islet cells. 39 The present data suggest that BMS increases serum insulin levels and reverses obesity-associated glucose intolerance.
To explore potential molecular mechanisms in lipid metabolism and lowering cardiovascular disease risk factors of BMS, expressions of PPAR-γ and related genes were investigated. PPARs are lipid-activated transcription factors that control energy homeostasis through genomic action. 40 PPAR-γ is highly expressed in adipose tissue, where it triggers cellular differentiation, promotes lipid storage, and modulates the action of insulin. 32 Excitingly, we observed that BMS activated the PPAR-γ in white adipose tissue. Similar findings were reported by Chao and Huang, 32 who showed that an ethyl acetate extract of bitter melon activated PPAR-γ and that the activation was comparable to that of thiazolidinedione antidiabetic drugs. PPAR-γ has been implicated as the primary receptor modulating the insulin-sensitizing activity of the thiazolidinediones, a class of antidiabetic drugs and fibrate hypolipidemic drugs. 41,42 The active compound in BMS responsible for activating PPARs is 9c,11t,13t-CLN. 43 Hence, the data reported in this study show that BMS improves hyperglycemic and hyperlipidemic conditions by acting as a ligand activator of PPAR-γ, which can induce the expression of genes that participate in lipid catabolism and or utilization of glucose. 44
A growing body of evidence suggests a possible role for PPAR-γ as a protective regulator in the cardiovascular system, particularly by decreasing hypertrophy, remodeling, and infarct size. 45 –47 Although the difference was not significant, we found that BMS down-regulated expression of the PPAR-γ gene in obese rat hearts. In addition, we studied the effect of BMS on the expression of atherogenic cytokine expression such as interferon-γ and the transcription factor NF-κB, which regulates expression of a variety of genes involved in immune responses, inflammation, proliferation, and programmed cell death. 48 We found that BMS had a particularly significant attenuating effect on NF-κB expression. This is the first study to show that BMS significantly down-regulates expression of NF-κB in obese rat hearts. Activation of this transcription factor plays an important role in the hypertrophic growth of the myocardium, which can lead to heart failure. 48 –50 This study provides evidence that BMS may work in improving cardiovascular health through down-regulating pro-inflammatory transcription factors such as NF-κB. We also speculate that the possible mechanism of BMS in regulating NF-κB is through inhibition or decreasing IκB kinase β complex, which is responsible for activation of NF-κB through IκB phosphorylation. It has been shown that anti-inflammatory drugs such as salicyclates inhibit NF-κB and its upstream activator, IκB kinase β, 51 and reduction in the expression of IκB kinase β by salicyclates significantly improved insulin sensitivity. 52 IκB kinase β activation of NF-κB in certain cells would enhance the production of tumor necrosis factor α, leading to insulin resistance. Hence, taken together, our data suggest that BMS improves insulin resistance and reduces cardiovascular disease risk factors by modulating the NF-κB pathway.
Conclusions
In this study, we not only demonstrated that BMS exerts beneficial effects in glucose metabolism but also improved serum and hepatic lipid profiles. We also demonstrated that BMS improves hyperglycemic and hyperlipidemic conditions by acting as a ligand activator of PPAR-γ and that the potential cardioprotective effects may be due to its ability to modulate NF-κB inflammatory pathways. In conclusion, BMS may offer a novel dietary strategy to prevent complications associated with metabolic syndrome. Further studies are needed to understand the dose-dependent effects of BMS.
Footnotes
Acknowledgment
This research was funded by the Institute of Food Science and Engineering, Division of Agriculture at the University of Arkansas.
Author Disclosure Statement
The authors have no competing financial interests or associations that might create a conflict of interest.