
Abstract
In this study, the protective effects of 17 Korean native plants against amyloid β peptide (Aβ)-induced oxidative stress were screened using the 2′,7′-dichlorofluorescin diacetate assay and the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. Ipomoea batatas exerted the highest protective effects against oxidative stress and was selected for further investigation. To confirm the protective activity of this extract, the I. batatas extract was fed to ICR mice that had been injected with Aβ to induce neuronal deficits. In these experiments, the extract of I. batatas significantly reversed Aβ-induced neurotoxicity as assessed by the passive avoidance test, a behavioral experiment. Moreover, I. batatas administration reduced the level of lipid peroxidation and increased catalase activities in biochemical studies using the brain tissue of mice. These results indicate that I. batatas might be beneficial against Alzheimer's disease, especially by limiting oxidative stress in the brain.
Introduction
P
The central nervous system is readily damaged by oxidative stress because of high oxygen consumption, which corresponds to its volume. The central nervous system constitutes only 2% of body weight but consumes 20% of total oxygen, which provides more opportunities to generate ROS. Moreover, it has relatively low levels of antioxidant compounds compared to those of other tissues. 4 In particular, the unsaturated carbon side chains of membrane lipids have been shown to be easily damaged by ROS attack, and their oxidation could result in membrane dysfunction and cell lysis. 5 Several studies have also shown increased oxidative damage to DNA, which is a feature of neuron cell death that has been observed in AD brains. 6,7 Thus, inhibiting and/or reversing the Aβ-induced deleterious effects by dietary antioxidant agents or rich foods is a rational approach to prevent AD.
Ipomoea batatas is a creeping dicotyledonous plant that belongs to the Convolvulaceae family. The edible tuberous root of sweet potato is long and tapered, and its colors range from red to purple brown and white. The sweet potato has been an important food source in human diets. Its nutritional composition consists of protein, carbohydrates, minerals, carotenoids, dietary fiber, and vitamins with small amounts of fat and sodium. Also, the sweet potato has been considered a promising alternative to artificial colorants due to its high concentrations of anthocyanin and β-carotene with good stability. 8,9
The aim of this work was to screen and measure the protective effects of various Korean edible plant extracts against Aβ-induced neurotoxicity. Rat pheochromocytoma cells (PC12) and Aβ-injected ICR mice were used as neuronal models, respectively. Biochemical markers of oxidative stress in the brain tissue of the mice were also analyzed after feeding I. batatas-supplemented diet.
Materials and Methods
Chemicals
2′,7′-Dichlorofluorescin diacetate (DCF-DA), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), dimethyl sulfoxide, hydrogen peroxide (H2O2), Aβ1–42 peptide, and
Preparation of plant extracts
Seventeen different Korean edible plants were obtained from the Gyeong-dong market or a local grocery store in Seoul, Republic of Korea (Table 1). Dried samples were pulverized, and five times the sample weight of ethanol was added. Samples were shaken for 24 hours at the rate of 125 rpm (1.57 g) and filtered through Whatman ashless filter paper No. 42 (Whatman International Ltd., Maidstone, UK). The sample residue was collected, and the above extraction process was repeated five times. The ethanol extract was evaporated under reduced pressure at 39°C. The dried extract was prepared to the concentration of 1 mg/mL with deionized water containing 5% (vol/vol) dimethyl sulfoxide and used for screening.
The results from the present study are given in bold.
Protective effects against reactive oxygen species (ROS) (%) =100 – {[(2′,7′-dichlorofluorescein formation of sample group with oxidative stress – 2′,7′-dichlorofluorescein formation of control group without oxidative stress) × 100]/(2′,7′-dichlorofluorescein formation of negative group with oxidative stress – 2′,7′-dichlorofluorescein formation of control group without oxidative stress)}.
Cell viability (%) = 100 – {[(formazan formation of control group without oxidative stress – formazan formation of sample group with oxidative stress) × 100]/(formazan formation of control group without oxidative stress – formazan formation of negative group with oxidative stress)}.
NA, not applicable.
Cell culture
The PC12 cell line (ATCC CRL-1721) was obtained from the American Type Culture Collection (Rockville, MD, USA) and maintained as previously described. 10
Measurement of intracellular oxidative stress and cell viability
Levels of intracellular ROS formation (DCF-DA) and cell viability (MTT) after inducing oxidative stress were evaluated as previously described. 11,12 Both assays were not blinded but were confirmed twice by further repeated experiments.
Animals and in vivo experimental design
ICR mice were purchased from Daehan Biolink (Chungnam, Republic of Korea) to assess in a behavioral study whether Aβ-induced neurotoxicity could be mitigated by supplementing the diet with the plant extract. The mice were housed in a room maintained at 23 ± 2°C with a 12-hour light/-dark cycle and fed for 4 weeks ad libitum a commercial diet (Purina Korea, Seoul) supplemented with the plant extract. The plant extract was mixed with the commercial diet at concentrations of 400, 800, and 1,200 mg/kg of body weight/day (0.25%, 0.50%, and 0.75%, respectively). The concentrations of plant supplementation were based on our previous experience with other plant extracts. There were nine mice per dietary group. Aβ was injected intracerebroventricularly to induce learning and memory impairment. 10 The control group mice were injected with reverse Aβ42–1, which is known to be nontoxic. All animal experimental procedures were approved by the Animal Care and Use Committee of Korea University, Seoul.
Passive avoidance test
After the diets were administered for 4 weeks, a behavioral study was carried out to assess the passive avoidance response of ICR mice in a nonblinded manner. 10
Biochemical experiments
Mice were sacrificed after performing the passive avoidance test. Then, brain tissues were collected and stored at −80°C until further analyzed using biochemical assays. Brain tissues were homogenized in 5 volumes of cold phosphate-buffered saline. The homogenates were immediately centrifuged twice at 10,000 rpm (33,600 g) for 10 seconds. The resulting supernatants were used for measuring the level of lipid peroxidation and catalase (CAT) activity.
Measurement of lipid peroxidation
The level of lipid peroxidation in brain tissues was analyzed by measuring thiobarbituric acid-reactive substance (TBARS) formation as described previously. 13
Measurement of CAT activity
CAT activity was measured using a slightly modified method. 14 In brief, 650 μL of phosphate buffer (50 mM, pH 7.0) and 50 μL of brain homogenate were mixed in a quartz cuvette. Then, the reaction was initiated by adding 300 μL of H2O2. The decomposition of H2O2 was measured spectrophotometrically at 240 nm (model UV-1601 spectrometer, Shimadzu, Kyoto, Japan). CAT activity was expressed as U/mg of protein.
Statistical analysis
Each result was expressed as the mean ± SD. The statistical significance of differences among groups was calculated by a one-way analysis of variance. Data were analyzed by Duncan's multiple-range test using the Statistical Analysis System (SAS) software package, version 9.0 (SAS Institute, Cary, NC). P values < .05 were considered statistically significant.
Results
Screening of Korean edible plant extracts
The results of screening 17 Korean edible plant extracts are presented in Table 1. All of the plants are commonly consumed as foods or teas in Korea. The levels of intracellular oxidative stress and cell viability were evaluated after a 2-hour H2O2 treatment on PC12 cells using DCF-DA and MTT. Of all the plant extracts investigated, the I. batatas extract exhibited the most potent inhibitory activity against 2′,7′-dichlorofluorescein (DCF) formation, which represents the level of oxidative stress in PC12 cells (Table 1).
Comparison of I. batatas peel versus flesh extract
A 2-hour H2O2 treatment resulted in a 142.03 ± 8.51% increase of intracellular oxidative stress and 66.56 ± 4.23% decrease in cell viability, respectively, compared with control group (Fig. 1). A 48-hour pretreatment with skin and flesh extracts resulted in protective activity against H2O2. Interestingly, however, the peel extract demonstrated more potent activity in both cell viability and inhibition of DCF formation in neuronal cells (cell viability [peel vs. flesh], 48.81 ± 0.38% vs. 40.15 ± 1.24%; DCF formation, 120.79 ± 4.30% vs. 189.69 ± 8.48%).
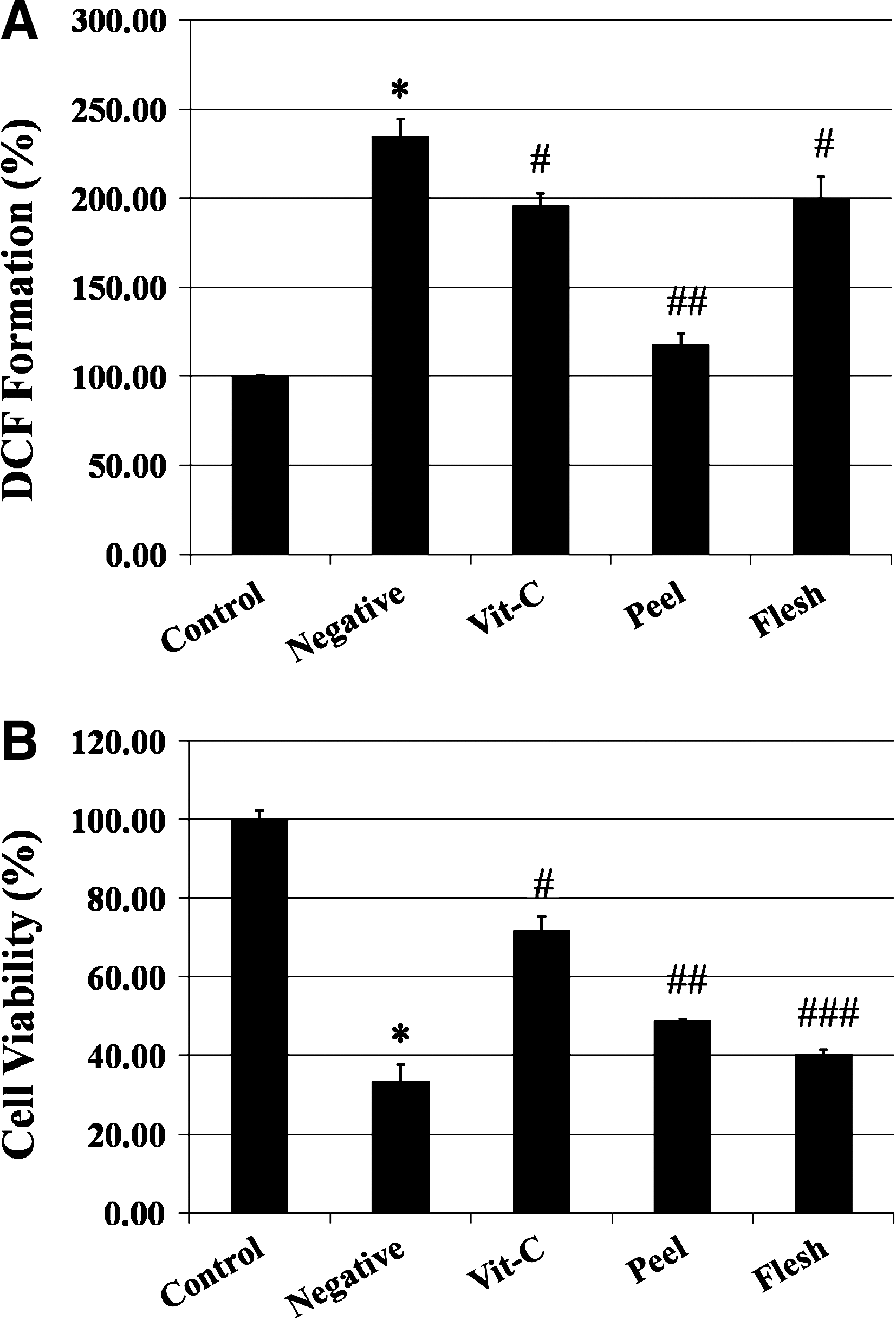
Comparison of the protective activities of I. batatas peel and flesh against oxidative damage. The control group was not treated. The negative group was treated with 100 μM H2O2 for 2 hours. The vitamin C (Vit.-C) group was preincubated with 100 μM vitamin C for 48 hours before treatment with 100 μM H2O2. Sample groups (peel and flesh) were pretreated with 1 mg/mL sample extract for 48 hours before treatment with 100 μM H2O2. (
Passive avoidance test
To confirm the protective activity of I. batatas, ICR mice were treated with one of three concentrations of the I. batatas peel extract for 4 weeks ad libitum, and Aβ was delivered via intracerebroventricular injection. As shown in Table 2, mice with Aβ-only treatment exhibited a significantly shorter step-through latency than the control group (Table 2, P < .01). In contrast, administration of the I. batatas peel extract for 4 weeks effectively reversed this Aβ-induced reduction in step-through latency by 96.7% (400 mg/kg of body weight), 104.9% (800 mg/kg of body weight), and 101.9% (1,200 mg/kg of body weight). There were no statistical differences in the step-through latency among the three groups receiving different doses (P < .01).
Data are mean ± SD values (n = 9). Duncan's multiple range test indicates a significant difference.
The mean latency in the passive avoidance test (*P < .01 vs. control group, # P < .01 vs. Aβ group).
Lipid peroxidation levels evaluated by measuring thiobarbituric acid-reactive substances (TBARS) in the brain tissues (*P < .05 vs. control group, # P < .05 vs. Aβ group).
Antioxidant enzyme (catalase [CAT]) activity (*P < .01 vs. control group, # P < .05 vs. Aβ group).
Control was injected with 410 pmol of nontoxic reverse Aβ42–1 per mouse.
Each mouse was injected with 410 pmol of Aβ1–42. Peel extract groups were injected with the same concentration of Aβ1–42 after feeding I. batatas peel extract for 4 weeks (400 mg/kg, 800 mg/kg, and 1,200 mg/kg per day, respectively).
Aβ, amyloid β peptide.
Measurement of lipid peroxidation
The effects of administration of I. batatas peel extract on lipid peroxidation was measured using the TBARS level in the brain tissues of ICR mice. Mice receiving the Aβ-only treatment had an increased level of TBARS compared with those of the control group (control group, 100.0 ± 1.31%; Aβ group, 127.7 ± 23.8%; P < .05), whereas all peel extract groups had lower levels of TBARS in the brain tissues compared with control (Table 2, P < .05). There were no statistically significant differences in TBARS level among the peel extract groups.
Measurement of CAT activity
The CAT activity in the brains of mice was measured. As shown in Table 2, Aβ injection decreased the CAT activity by 65.1 ± 9.6%. In contrast, compared to the Aβ-only mice, all peel extract groups had statistically significant higher CAT activity (P < .01).
Discussion
The initial approach for AD therapy was to inhibit cholinesterase, which is responsible for degrading acetylcholine in the synaptic clefts, based on evidence of the loss of basal forebrain cholinergic neurons in AD patients. 15 This “cholinergic hypothesis” has led to the development of cholinesterase inhibitors such as tacrine, denepezil, and ravastigmine for AD treatment. Their efficacies, however, are modest and provide only palliative effects on the progress of the disease. Moreover, the use of some of the above cholinesterase inhibitors (e.g., tacrine) was limited as they resulted in considerable side effects, including hepatotoxicity. 16 The newer approach for preventing and/or treating AD is blocking Aβ-induced neurotoxicity, which is known to cause cognitive decline through various mechanisms (e.g., inflammation, synaptic damage, and mitochondrial dysfunction). 17,18 Although the mechanisms of pre-/postsymptomatic damage are not fully elucidated, it seems clear that intracellular oxidative stress and production of ROS from Aβ play major roles in neurodegeneration. 7,19 Given the variety of antioxidants in the diet, the screening of commonly consumed foods for effective and specific compounds against Aβ-induced oxidative stress that may have less adverse effects than pharmacologic agents is warranted.
Based upon the screening of results, even though several sample groups displayed higher cell viability (Table 1), the Aβ-specific protective activity of I. batatas was investigated further as it showed the highest potency against intracellular oxidative stress. To compare the antioxidative activity of its peel and flesh, I. batatas was divided into skin and body parts. Consequently, the peel extract showed higher protective effects against oxidative stress in both MTT assay and DCF formation assay (Fig. 1). These results are in agreement with a previous study that compared the total phenolic and anthocyanin values between the peel and the flesh part of sweet potato. 20 It is well known that Aβ generates oxidative stress, including H2O2. H2O2 itself and hydroxyl radical (•OH), which can be produced by either the Fenton reaction or the Haber-Weiss reaction from H2O2, may damage most cellular macromolecules such as proteins, lipids, mitochondria, and DNA, eventually leading to cell death. 4 –6,17 In this respect, these results suggest that the antioxidative capacity of I. batatas, particularly the peel, may provide protection against oxidative damage caused by Aβ.
In order to evaluate the effects of the dietary intervention against Aβ neurotoxicity, it is important to use the proper methods to induce cognitive deficits and measuring memory ability. Mice given an intracerebroventricular injection of Aβ have been commonly used for AD research as it has been previously shown that intracerebroventricular injection of Aβ effectively causes memory deficits. 10,21 During the experimental period of this study, all mice receiving the peel extract diets gained body weight normally (control group, 40.3 ± 1.46 g; Aβ group, 38.4 ± 2.17 g; peel extract groups, 41.4 ± 2.75 g), and no significant differences in aspartate aminotransferase and alanine aminotransferase were observed between the peel extract groups and control group (data not shown). The passive avoidance test was performed 3 days after injection of Aβ to test the effects of the I. batatas peel extract. The step-through latency from the light chamber to the dark chamber was used to measure the learning and memory ability of mice. As described in Results, our data indicated that administration of the I. batatas peel extract effectively reversed Aβ-induced cognitive deficits in the passive avoidance test without any acute toxicity.
There is a significant amount of evidence showing that oxidative damage is closely related with neurodegeneration and the pathogenesis of AD. In addition, many studies have found increased levels of lipid peroxidation and decreased levels of polyunsaturated fatty acids, which are the major constituents of neuronal cell membranes, in the brain tissue of AD patients. 22,23 Besides the direct damage to lipids by ROS, by-products of peroxidation such as 4-hydroxynonenal and acrolein can also exert neurotoxic effects on neurons. Several studies have indicated that these by-products not only impair enzymes such as Na+,K+-ATPase, 24 glucose transport, glutamate uptake, 25 and calcium homeostasis, 26 but also damage DNA bases. 27 Therefore, it may be possible to delay or prevent the progress of AD through the oral administrations of potential antioxidants that suppress the level of lipid peroxidation in brain tissues. As shown in Table 2, all groups given sample extracts showed significantly decreased level of lipid peroxidation in the brain tissues, and this may explain the beneficial effects of I. batatas against cognitive deficits that were induced via intracerebroventricular injection of Aβ.
On the other hand, oxidative damage in brain could be accelerated by a failure of antioxidant defense systems. Hence, antioxidant enzymes are important in preventing the accumulation of ROS. CAT, one of these antioxidant enzymes, catalyzes the decomposition of H2O2 to water and oxygen. 14 As shown in Table 2, CAT activity negatively corresponded to the level of TBARS; thus, it is possible that the protective effects of I. batatas are due to the increased activity of antioxidative enzymes, which would decrease the level of lipid peroxidation.
Among the several pathological features of AD, deposition of Aβ has been demonstrated to play a pivotal role in neurodegeneration. Even though the exact mechanism of neurotoxicity induced by this peptide still remains unclear, growing evidence supports the notion that Aβ may increase intracellular oxidative damage and inflammatory response in AD. 18,19 These burdens are closely related with learning and memory deficits. 2 The brain tissue is particularly susceptible to oxidative stress because of its characteristics, including a high metabolic rate and polyunsaturated fatty acid content and low levels of antioxidant defense enzymes, yet it is rich in enzymatically active transition metals, which can accelerate radical generation. 4 Therefore, it is possible that dietary antioxidants may either lower the level of oxidative stress or contribute to maintaining ROS homeostasis in the brain. Several studies have been done in terms of examining the protective effects of consuming antioxidant containing foods and supplements (e.g., green tea, Gingko biloba, and red wine). 5,28 However, this is the first report where the I. batatas peel extract has been shown to possess anti-Aβ activities by lowering oxidative stress in the brain.
One potential limitation of this study is that the performance of the behavioral test and cell viability assay (i.e., MTT reduction assay) could not be blinded in order to reduce any possible bias. However, all of the in vitro experiments were done by only one of the authors (J.K.K.) and then confirmed by further repeated experiments as well. Another limitation is the concentrations (i.e., 400, 800, and 1,200 mg/kg of body weight/day) that were given to mice; although these concentrations did not exhibit any hepatotoxicity in mice, they are difficult to achieve through direct consumption of sweet potato peels based upon the extraction yield (19.4%). To achieve these concentrations in a 70-kg adult, he or she would have to eat more than 0.5 kg of sweet potato peel/day; thus it will be more reasonable to have as a supplement. And, future studies are also warranted investigating the efficacy of dietary concentrations lower than those tested here. Lastly, we obtained the samples from a local grocery store in order to use what is normally consumed; however, this approach may also be limited because of biochemical and nutritional variations of I. batatas depending on genetic and environmental factors. 8
In summary, we showed that the I. batatas peel extract effectively reversed the deleterious effects of oxidative damage in both in vitro and in vivo models of AD. Moreover, biochemical experiments using brain tissues clearly showed lowered oxidative stress levels, and these biochemical changes could explain, at least in part, the improved learning and memory ability of the mice in the behavioral test. Therefore, it is suggested that consumption of I. batatas, especially with the peel, might be beneficial to mitigate the effects of AD by possibly decreasing oxidative stress.
Footnotes
Acknowledgment
This work was supported by a National Research Foundation of Korea grant (KRF-2008-313-F00165) funded by the Korean Government.
Author Disclosure Statement
The authors have declared no conflict of interest.