
Abstract
The effect of daily ingestion of collagen hydrolysate (CH) on skin extracellular matrix proteins was investigated. Four-week-old male Wistar rats were fed a modified AIN-93 diet containing 12% casein as the reference group or CH as the treatment group. A control group was established in which animals were fed a non–protein-modified AIN-93 diet. The diets were administered continuously for 4 weeks when six fresh skin samples from each group were assembled and subjected to extraction of protein. Type I and IV collagens were studied by immunoblot, and activities of matrix metalloproteinase (MMP) 2 and 9 were assessed by zymography. The relative amount of type I and IV collagens was significantly (P < .05) increased after CH intake compared with the reference diet group (casein). Moreover, CH uptake significantly decreased both proenzyme and active forms of MMP2 compared with casein and control groups (P < .05). In contrast, CH ingestion did not influence on MMP9 activity. These results suggest that CH may reduce aging-related changes of the extracellular matrix by stimulating anabolic processes in skin tissue.
Introduction
I
Skin characteristics are known to be affected by endogenous and environmental factors, including chronological aging, ultraviolet radiation, hormones, and nutrition. These factors impair skin metabolism, up-regulating synthesis of matrix metalloproteinases (MMPs) that degrade skin collagenous extracellular matrix. Collagen atrophy is considered the major characteristic of skin aging. Type I collagen is the predominant collagen found in dermis, being responsible for skin tensile strength. At the same time, type IV collagen forms a network, highly cross-linked, maintaining the mechanical stability of the basement membrane, and seems to be decreased at the bottom of the wrinkles. 1 MMPs are a family of structurally related molecules, including interstitial collagenases (MMP1 in humans and MMP13 in rodents) and gelatinases (MMPs 2 and 9), that are capable of degrading all components of the extracellular matrix such as collagen, elastin, fibronectin, proteoglycans, and laminin. 2 In particular, MMPs 1 and 13 initiate the degradation of type I collagen, and MMP9 further degrades the collagen fragments produced. In contrast, MMP2 degrades type IV collagen, which leads to collagen deficiency and wrinkling. MMPs 2 and 9 are known to play an important role in the processes of skin metabolism and aging. 3
Dietary supplements have demonstrated beneficial effects on skin health. Experimental and clinical trials investigating the effects of oral supplementation with vitamins, polyphenols, micronutrients, and proteins have indicated that dietary compounds can modulate skin function. 4 –8 Moreover, the photoprotective potential of antioxidant intake has been the subject of a considerable number of studies. 9,10 The classical route of administration of active compounds is by topical application, and manufacturers have substantial experience of formulating ingredients in this field. In recent years, protein hydrolysates have been studied as potential dietary ingredients and in development of functional foods. The newest relationship between food and skin has drawn considerable attention due to the physiological effects of some dietary compounds on the skin-aging process. 11
Collagen hydrolysate (CH) and gelatin are currently used in diverse fields, including food, cosmetic, and biomedical industries. Gelatin is a high-molecular-weight polypeptide derived from collagen, the primary protein component of connective tissues. Industrial preparation of gelatin involves partial hydrolysis of the native structure of collagen to obtain the water-soluble form. Gelatin has chemical and physical properties that are interesting from the technological perspective. A further enzymatic degradation of gelatin results in a product called CH, which contains peptides with molecular mass ranging from 3 to 6 kDa. The most important sources for gelatin and CH production are bovine hide and bone and pigskin. 12,13
There is an agreement that biological effects promoted by food-derived collagen peptides are related to the ingestion of collagen in its hydrolyzed form. 14 –22 CHs have been reported to have beneficial biological functions that might justify their use in food supplements and pharmaceutical preparations. Clinical investigations suggest that ingestion of CH reduces pain in patients suffering from osteoarthritis. 14,15 CH is also involved in cartilage matrix synthesis. 16 Some collagen peptides exhibit antihypertensive activity, inhibiting the effect of angiotensin I converting enzyme (ACE) 17,18 and promoting inhibition of cardiovascular damage to the endothelial cells via its ACE inhibitory activity and regulation of nitric oxide and intercellular adhesion molecule-1. 19 Based on in vitro studies, bioactive collagen peptides derived from fish, bovine, and porcine skin have also shown potent antioxidative activities in different oxidative systems. 20 –22
Before speculating about the mechanism of skin effectiveness of CH, it is important to emphasize that CH is able to cross the intestinal barrier, reaching blood circulation and becoming available for metabolic process and storage in skin. 23 –25 To date, preclinical and clinical studies have demonstrated that CH may improve skin conditions and protect skin from ultraviolet damage. 26 –30 Elucidating, at least partially, the action mechanism of CH in skin anabolic process after its oral administration may be a major contribution to understanding its feasibility for use in dietary products with the purpose of skin improvement.
We investigated the effects of CH intake on type I and IV collagen biosynthesis and MMP activity in order to determine the potential action mechanisms of CH in skin, which could support the value-added utilization of CH and food supplements.
Materials and Methods
A bovine hide CH (GELITA® CPB1000) was kindly supplied by Gelita do Brasil Ltda. (Cotia, SP, Brazil). According to the manufacturer, this product was obtained by a sequential enzymatic hydrolysis, using a cocktail of enzymes, including alcalase. Commercial casein (CS) as well as the other components of the diet were purchased from MCassab Ltda. (São Paulo, SP, Brazil).
Animals and diets
All animal experiments were performed in accordance with the guidelines of the Committee for Animal Research of the State University of Campinas, São Paulo.
Four-week-old male Wistar rats were housed in individual cages at 20 ± 2°C on a 12-hour light/12-hour dark cycle, with free (ad libitum) access to water and diets. After the acclimation period, the animals were divided into three groups (six animals per group) such the body weight did not differ significantly among groups. The animals were fed a modified AIN-93 diet 31 (isocaloric and isoproteic) containing 12% protein of CS—as the reference group—or a special CH enriched in essential amino acids, according to Food and Agricultural Organization/World Health Organization guidelines 32 —as the treatment group. A negative control group was established in which the animals were fed a non–protein-modified AIN-93 diet. The diets were administered continuously for 4 weeks. Body weight change and food intake were recorded three times a week.
The composition of the modified AIN-93 diet is shown in Table 1, and the centesimal composition of the two protein sources used in this study is described in Table 2.
Source: Reeves et al. 31
Collagen hydrolysate or casein, calculated with the respective nitrogen-to-protein conversion factors (5.55 and 6.38).
According to the AIN-93 diet proposed by Reeves et al. 31
Nitrogen factor for commercial casein (CS) and collage hydrolysate (CH) of 6.38 and 5.55, respectively.
Determined by the Association of Official Analytical Chemists' method. 33
Determined by the methodology proposed by Bligh and Dyer. 34
Calculated by difference: carbohydrate = 100 – (protein + fat + ash + moisture).
Molecular weight and amino acid compositions
The average molecular weight of CH was determined by gel permeation chromatography on a TSK gel 2000 SWXL column (Tosoh, Tokyo, Japan), using a high-performance liquid chromatography system (Waters, Milford, MA, USA). The mobile phase used was 400 mM sodium dihydrogen phosphate buffer, pH 5.3. The samples were eluted at a flow rate of 0.6 mL/minute and monitored at 214 nm at 25°C. A molecular size calibration curve was prepared from the average retention times of BrCN peptides (human collagen-type I) of 38 kDa, 25 kDa, 18 kDa, 13.5 kDa, and 3.5 kDa (Sigma Chemical Co., St. Louis, MO, USA). Then the amino acid compositions of CH were analyzed by the Waters Pico-Tag system (Waters) described by White et al. 35
Skin protein measurement
After the 4-week experimental period, animals were sacrificed by cervical dislocation, and six fresh skin samples from each group were assembled and subjected to extraction of protein. Skin proteins were extracted with a 50 mM Tris-HCl buffer (pH 7.4) containing 1% sodium deoxycholic acid, 150 mM NaCl, 1% Nonidet P-40, and protease inhibitor cocktail (catalog number P8340, Sigma) containing 4-(2-aminoethyl)benzenesulfonyl fluoride hydrochloride, pepstatin A, E-64, bestatin, leupeptin, and aprotinin after being frozen-crushed in liquid nitrogen. 36 The extract was separated by centrifugation at 14,000 g for 7 minutes at 4°C. The supernatant was collected, and the amount of protein was estimated by BCA assay (Pierce Inc., Rockford, IL, USA). The water-soluble complex formed exhibits a strong absorbance at 562 nm that is nearly linear with increasing protein concentrations. 37
Analysis of type I and IV collagens by immunoblotting
After the protein concentration was measured, samples were loaded in sample buffer containing 0.3 M Tris-HCl, 5% sodium dodecyl sulfate, 50% glycerol, and 100 mM dithiothreitol (Pierce). Equivalent protein samples (40 μg) were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis on 10% (wt/vol) polyacrylamide gel. Proteins were transferred to a polyvinylidene difluoride membrane for 2 hours at 100 V. The membrane was soaked in blocking buffer (Tris-buffered saline [25 mM Tris, 150 mM NaCl, and 2 mM KCl, pH 7.4] containing 0.1% Tween-20 and 5% skimmed milk) for 1 hour. The primary mouse monoclonal antibody against type I collagen (Sigma) and rabbit monoclonal antibody against type IV collagen (Rockland Immunochemicals, Inc., Gilbertsville, PA, USA) were used at a dilution of 1:500 in blocking buffer. Secondary anti-mouse and anti-rabbit monoclonal antibodies conjugated with horseradish peroxidase (Amersham Inc., Arlington Heights, IL, USA) were diluted to 1:1,000 in blocking buffer. The resulting bands were detected by the horseradish peroxidase reaction with an enhanced chemiluminescence kit (Amersham).
Immunoblot band intensities were quantified by Image J software (National Institute of Mental Health, Rockville, MD, USA). To confirm equal loading conditions membranes were stripped and reprobed with β-actin antibody (Sigma).
Activities of MMPs 2 and 9 by zymography
The same skin extracts as described above were used for the zymographic study. After the protein concentration was measured, samples were resuspended in sodium dodecyl sulfate–polyacrylamide gel electrophoresis sample buffer (without β-mercaptoethanol) (Pierce). A total of 10 μg of each sample was loaded in polyacrylamide gels containing 0.2% gelatin type A from porcine skin (Sigma). After electrophoresis, the gels were washed in 2.5% Triton X-100 for 30 minutes, equilibrated in 10 mM Tris (pH 8.0), and incubated at 37°C in a developer buffer containing 50 mM Tris (pH 8.0), 5 mM CaCl2, and 0.02% NaN3 for 16 hours. The gels were stained with 0.2% Coomassie Brilliant Blue and destained with a solution of acetic acid and methanol. Gelatinolytic bands were observed as clear zones against the blue background, and the intensity of bands was estimated using Image J software.
Statistical analysis
Data were analyzed using Minitab software, version 15.1.1.0 (Minitab Inc., State College, PA, USA). Differences among groups were assessed by analysis of variance, followed by the Tukey–Kramer multiple comparison post-test. Values of P < .05 were considered statistically significant.
Results
Molecular weight and amino acid compositions
The gel permeation chromatography technique was applied to analyze the molecular size distribution of CH, as shown in Figure 1. The results revealed an average molecular mass of 3,551 Da and a main fraction of peptides with molecular sizes ranging from 1,000 to 3,000 Da (66.06% of total protein). The molecular mass of the main peak of CH was 1,547 Da, and its relative proportion was more than 90%.
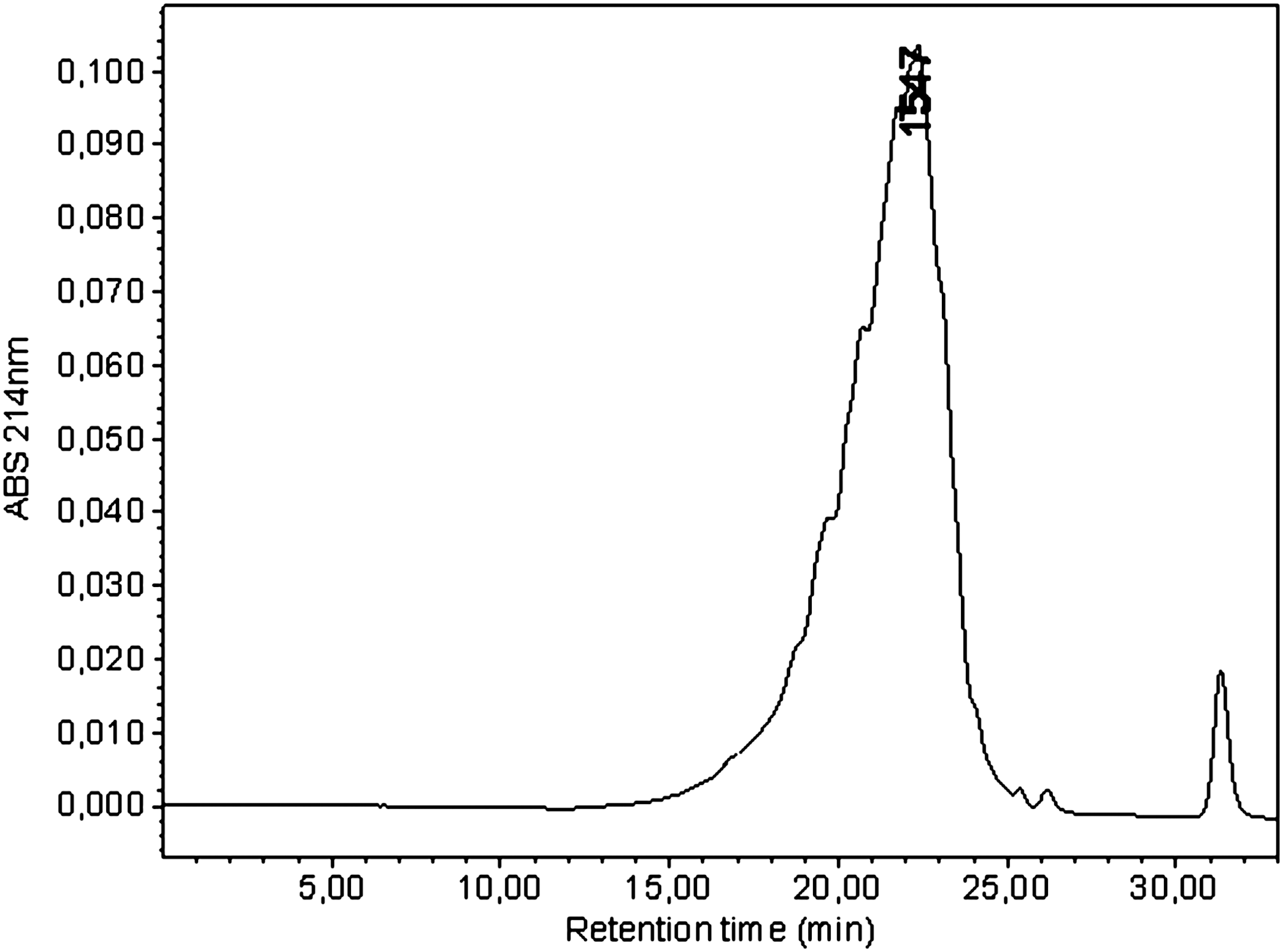
Molecular weight of CH analyzed by the gel permeation chromatography technique. ABS, absorbance.
In order to know whether the effect of CH ingestion is collagen-specific or is due to the ingestion of protein itself, animals were fed a diet containing the same amount of protein, 12% of CS or CH, changing the protein source. CS is considered by the Food and Agriculture Organization/World Health Organization 32 as a reference standard because of its essential amino acid profile. In this study, CS contained all amino acids, whereas CH showed deficiency in all essential amino acids and the complete absence of tryptophan (Table 3). CH had high contents of glycine (24.5%), glutamic acid (10.1%), arginine (8.1%), proline (13.8%), and hydroxyproline (7.4%) residues and small amounts of tyrosine, cysteine, histidine, and methionine residues.
ND, not determined.
The amount of protein intake throughout the study differed between the CS and CH groups (57.2 ± 6.2 g and 46.5 ± 6.0 g, respectively) (data not shown). Consequently, animals grew quite differently, and the final body weight in the CS group was significantly higher (268.9 ± 31.6 g) compared with the CH group (197.4 ± 22.2 g). Although CH is deficient in essential amino acids, it has an excellent digestibility 38 and is often used to supplement other proteins to give a higher protein value. Moreover, it is interesting to note that from the nutritional perspective CH is characterized by a considerably high safety profile. 12
CH ingestion increased skin type I and IV collagens
In immunoblots, antibodies recognized type I collagen protein in both the CH and CS groups. However, CH resulted in a statistically significant (P < .05) increase of type I collagen synthesis, after 4 weeks of daily ingestion (Fig. 2). The relative amount of type I collagen increased up to fourfold after CH intake (3.4 ± 1.7) compared with the reference diet CS group (0.8 ± 0.5). Similar results were found relating to type IV collagen expression: the collagen IV amount increased up to threefold after CH intake (6.7 ± 1.1) compared with the reference diet CS group (2.3 ± 1.2) (P < .05) as observed in Figure 2.
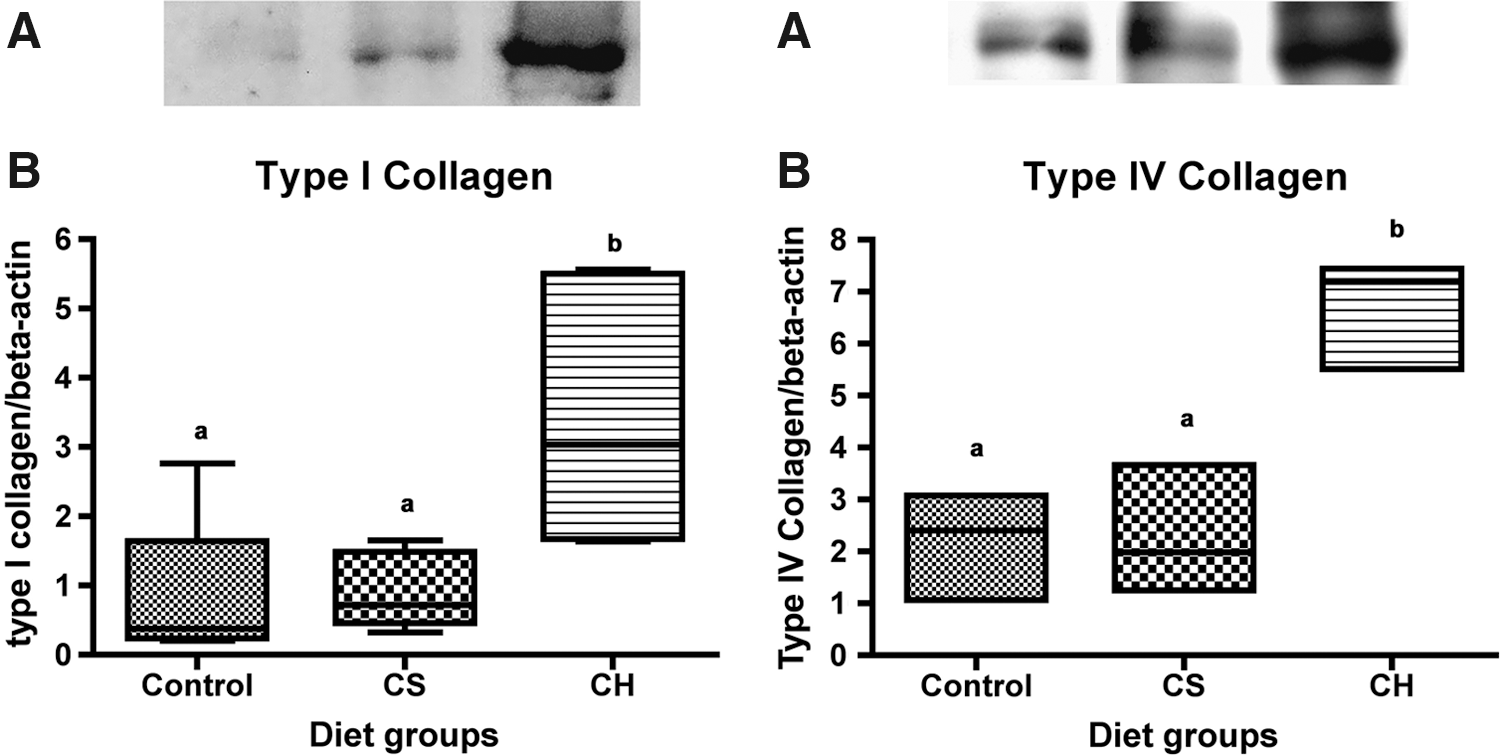
(Left panel) Type I and (right panel) type IV collagen expression in the skin of rats determined by immunoblotting. Control animals were fed a non–protein-modified AIN-93 diet. CS animals were fed a modified AIN-93 diet containing 12% CS, and CH animals were fed one with 12% CH. (
CH decreased skin MMP2 activity
Zymography showed gelatinolytic bands at molecular weights of those corresponding to MMPs 2 and −9 (Fig. 3). Gelatin is a general substrate for proteolytic activity, suitable for the detection of different proteases. However, gelatin zymography is particularly suitable for MMPs 2 and 9. The oral administration of CH was associated with a statistically significant decrease (P < .05) of MMP2 activity shown by zymography (Fig. 4) in relation to the reference group (CS). In contrast, MMP9 activity was not influenced by administration of CH in the diet as shown in Figure 4.

Zymograms for the determination of activities of matrix metalloproteinases (MMPs) 2 and 9 in the skin of rats fed diets containing non-protein (control), CS, and CH. Gelatinolytic activities of MMPs 2 and 9 in skin lysates were detected by electrophoresis of soluble protein on a gelatin-containing 10% polyacrylamide gel. Representative zymograms were obtained from three independent experiments.
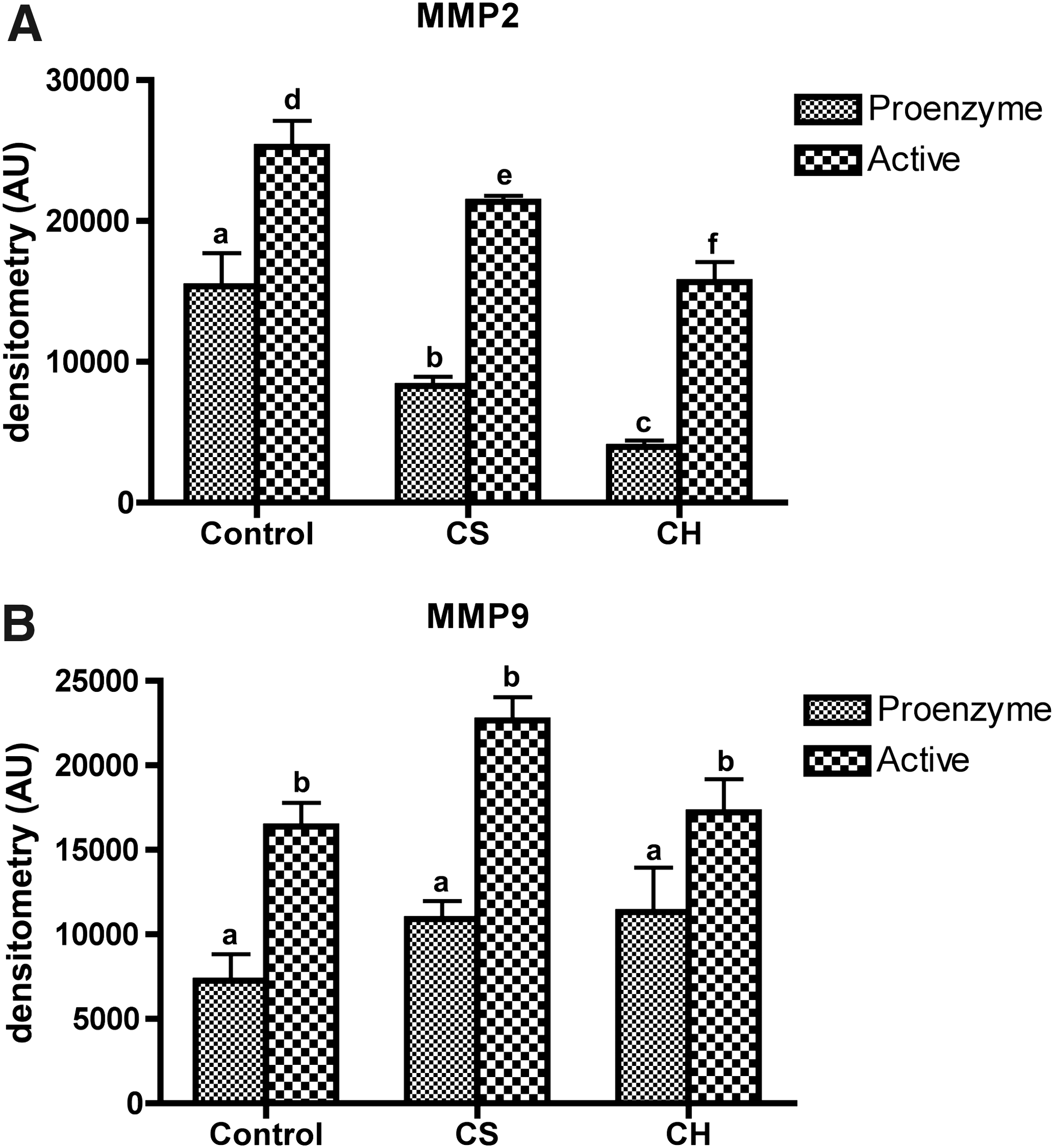
Densitometry of both latent and active forms of MMPs (
Discussion
CHs have long been used in pharmaceuticals and food supplements for improving skin and cartilage tissues. It is digested and absorbed in the digestive tract, appears in the human blood partly in a peptide form, 23,24 and is accumulated in skin for up to 96 hours as shown by Oesser et al. 25 Because of its biochemical similarity to genuine collagen in connective tissue and especially to type I collagen in dermis, it has been suggested that oral ingestion of CH might have beneficial effects and can promote both collagen type I biosynthesis and the repair process in a dermal wound. Thus CH ingestion would have a potential positive impact, protecting skin from the aging process.
Studies have reported that food-derived collagen peptides in human blood exert a chemotactic role on skin fibroblasts 39 and increase the migration and growth of mouse skin fibroblasts. 40 However, the effects of CH on expression of skin collagen protein and activity of MMPs have not yet been examined to date. Our present study provides additional evidence for an effect of CH on skin.
Despite of its reduced nutritional profile, CH promoted stimulatory effects on skin tissue, increasing expression levels of the most abundant extracellular matrix protein in skin, type I collagen (Fig. 2). The CH effect appears collagen-specific because CS (reference diet group) failed to stimulate collagen expression. Our results agree with those of Matsuda et al. 41 They investigated effects of CH ingestion on fibroblast and collagen densities of pig skin; CH was administered orally to pigs at 0.2 g/kg of body weight/day for 62 days, and its effects were compared with those of lactalbumin and water controls. Fibroblast density and diameter and density of collagen fibrils were significantly larger in the collagen peptide group than in the control groups. This implies that the effect of CH was protein-specific and did not depend merely on an increase of amino acid intake.
Although our results are related to expression levels of type I collagen in normal rat skin, they are also in accordance with those of Tanaka et al. 42 These authors demonstrated that CH ingestion inhibited ultraviolet B-induced decrease of type I collagen, thus improving skin conditions in mice.
Taking into account that a gradual decrease in type I collagen level is known to occur during the course of a lifetime and induces skin aging, 3 an increase of type I collagen after consumption of CH is desired for anti-aging benefits.
Our results complement those reported by Nishimoto et al., 43 investigating the effects of oral administration of CH on the hydroxyproline content of mouse skin. CH was administered at 50 and 100 mg/kg for 21 days and increased the hydroxyproline content in unshaved and shaved skin, which may be related to skin collagen synthesis.
Based on in vitro studies, Rodrigues 44 showed that CH at concentrations ranging from 0.1 mg/mL up to 6 mg/mL stimulated fibroblast to secrete 1.8-fold more collagen than the untreated cells. Furthermore, fibroblast cell cultures treated with CH showed a higher percentage of cells in S phase (25.8 ± 4.0%) than control cells (18.9 ± 2.0%) and decreased percentage of cells in apoptosis (8.2 ± 4.0%) compared with controls (22.4 ± 3.0%).
In our study, it was also found that daily CH ingestion for 4 weeks significantly suppressed MMP2 activity compared with the control and reference groups. In contrast, MMP9 activity was not influenced by CH ingestion (Fig. 4). MMPs can degrade skin collagen and contribute to the connective tissue breakdown, resulting in collagen deficiency. This deficiency may be followed by skin wrinkling. 3 MMPs 2 and 9 are known to play an important roles in the processes of skin aging and metabolism. 3 Inhibition of MMPs can take place at different levels, from their gene expression to enzyme activity. Because MMPs 2 and 9 belong to the group of MMPs that use gelatin as their substrate, gelatin zymography was carried out to screen their activity. 45
MMP2 degrades type IV collagen, the major structural component of basement membrane in skin, being partly responsible for skin wrinkles and furrow formation. 1 We demonstrated that CH uptake suppressed MMP2 and increased the expression level of type IV collagen (Fig. 2). These effects may contribute to explain the effects of CH on photodamaged skin previously reported. 29,30,42
It is commonly known that MMPs injure the dermis by two related mechanisms: (1) direct degradation of collagen and (2) indirect inhibition of collagen synthesis by MMP-generated collagen fragments. Direct MMP13-induced breakdown of type I collagen generates high molecular-weight fragments. 46 These fragments serve as negative regulators of type I collagen synthesis. On the other hand, further breakdown of MMP13-cleaved collagen by MMP9 can diminish this inhibition. 3,47 It is remarkable that MMP9 activity remained unchanged after CH intake in our study and may contribute to the CH anti-aging benefit.
Taken together, CH uptake increased type I collagen expression levels and did not suppress MMP9 activity. Based on these results we may hypothesize that the presence of small, food-derived collagen peptides in skin after CH ingestion may simulate those in damaged skin. These peptides would inhibit further collagen degradation by MMPs and mitigate indirect inhibition of collagen synthesis by MMP-generated collagen degradation.
We may also speculate on signaling pathways underlying CH-mediated enhancement of collagen expression. Collagen peptides might bind cell surface receptors, stimulating biosynthesis of collagen and other extracellular matrix molecules. Alternatively, collagen peptides might positively affect interaction between extracellular matrix and fibroblasts. The MMP2 inhibitory effect shown by CH may be mediated by the inhibition of the intracellular signal transduction response. However, the effects of CH on the cell signal pathways need to be further studied.
Conclusions
Although CH does not present high nutritional quality profile, our findings indicated a stimulatory effect of CH on rat skin metabolism. CH ingestion reduced MMP2 activity and increased expression of types I and IV collagens. Thus, CH can be used as a potential candidate for reducing aging-related changes of extracellular matrix by stimulating anabolic processes in skin tissue. Further studies are necessary to elucidate its anti-aging mechanisms, particularly those related to the signaling pathways, in parallel with in vivo tests. At least fair scientific evidence suggests that clinical effects on skin may be achieved by CH ingestion. However, preclinical results are too preliminary to make general clinical recommendations.
Footnotes
Acknowledgment
This work was funded by GELITA of Brazil Ltd.
Author Disclosure Statement
V.Z. and G.A.deC. are employees of GELITA of Brazil Ltd. No competing financial interests exist for V.deF., M.daC.R., R.G.J., and G.M.M.-S.