
Abstract
Tea polyphenolics such as catechins are known to have the potential to inhibit many bacterial pathogens. Helicobacter pylori has been identified as an etiologic agent in the development of gastric ulcer, peptic ulcer, gastritis, and many other stomach-related diseases. In this study, we investigated the effect of 9 tea extracts—3 different brands representing 4 different processed types (white, green, oolong, and black)—on the inhibition of H. pylori. Extraction times of 2 and 5 minutes were compared. Most 5-minute extracts showed H. pylori inhibition, whereas 2-minute extracts only of Choice darjeeling black and Tazo white showed inhibition. No recovery was observed after the addition of 0.5 and 5 mM proline, indicating that tea polyphenols do not inhibit H. pylori by inhibition of proline oxidation via proline dehydrogenase. Extracts that showed inhibition were further evaluated for their effect on beneficial lactic acid bacteria. None of the samples showed inhibition, suggesting that tea might be able to inhibit H. pylori without affecting the beneficial lactic acid bacteria. High-performance liquid chromatography indicated the presence of gallic acid, quercetin, caffeine, and tea catechins (including catechin, epicatechin, and epigallocatechin) in all the tea samples. Our study indicates that tea can be potentially used as a low-cost dietary support to combat H. pylori–linked gastric diseases without affecting the beneficial intestinal bacteria.
Introduction
H
Several reasons explain the failure of H. pylori eradication therapy; these include antibiotic resistance by chromosomal mutation, limited efficacy of antibiotics due to low gastric pH, and high bacterial load. 11,12 Ongoing studies are investigating whole-food and healthy dietary support agents in lieu of costly and rigorous antibiotic treatment regimens, which are associated with many side effects.
Phenolic compounds that have been shown inhibit H. pylori include tannins, tea catechins, quercetins. 13 –15 Tea, which is a commonly consumed household beverage, has also been shown to inhibit H. pylori. 16 –18
Lactic acid bacteria, such as Lactobacillus species and Bifidobacterium species, are a class of probiotics, which are defined as living microorganisms that exert health benefits beyond basic nutrition, when ingested in certain numbers. 19 Lactic acid bacteria confer several benefits to the host, including better absorption of nutrients from food, decrease in lactose intolerance in some individuals, control of diseases originating from intestinal infections, potential to control some types of cancer, and stimulation of host immune response. 20 –22 Oregano and rosemary delay the growth and acid production of lactic acid bacteria. 23,24 Other authors have reported that tannins, tannic acid, and related compounds did not affect the growth of lactic acid bacteria or had very slight effects at high concentrations. 25,26 Existing reports are contradictory regarding the effect of tea on some species of lactic acid bacteria. Horiba et al. 27 reported that green tea inhibited the growth of Bifidobacterium bifidum, whereas Ahn et al. 28 found that green tea did not affect Bifidobacterium. In addition, stimulatory effect of water extract of Panax ginseng has been reported on the growth of Bifidobacteria. 29
Antibacterial and bactericidal effects of tea have been shown against many bacteria. 27,28,30 Therefore, we explored the relevance of daily consumption of several types of commonly consumed tea (green, black, oolong, and white) and extraction time for H. pylori inhibition. We also assessed the effects of tea consumption on different strains of lactic acid bacteria with probiotic potential, such as L. acidophilus, B. longum, and L. plantarum. Furthermore, we investigated the mode of action of H. pylori inhibition by tea on the basis of the rationale that simple phenolics may mimic proline analogues and may exhibit antimicrobial activity by inhibition of proline oxidation via proline dehydrogenase. 31 –34
Materials and Methods
Sample extraction
One tea bag was added to 200 mL of boiling water. It was then allowed to steep for 2 or 5 minutes. The extract was then allowed to cool at room temperature and centrifuged at 10,000g for 10 minutes. Samples evaluated were as follows: Choice tea (Seattle, WA, USA), white, Japanese green, oolong, and darjeeling; Tazo tea (Portland, OR, USA), Berry Blossom White, green, and black; and Bigelow's tea (Fairfield, CT, USA), green and Earl Grey. All teas were purchased from Whole Foods Market, Hadley, MA, USA. Absorbance of the extracts were measured at 400 and 600 nm to determine whether the extraction were uniform.
Bacterial strains
Helicobacter pylori (strain American Type Culture Collection 43579, which originated from human gastric samples) was obtained from the American Type Culture Collection (Rockville, MD, USA).
The lactic acid bacteria strains used in this study were B. longum (organisms that were isolated from yogurt in a previous study), 35 L. acidophilus, supplied by Rosell Institute Inc. (Montreal, Canada; lot number XA 0145, sequence number 00014160), and L. plantarum (1) (ATCC 9019).
High-performance liquid chromatography analysis of phenolic profiles
Hot water extracts (2 mL) were filtered through a 0.2-μm filter. A volume of 5 μL of extract was injected by using an Agilent ALS 1100 auto sampler into an Agilent 1100 series high-performance liquid chromatography (HPLC) unit (Agilent Technologies, Palo Alto, CA, USA) equipped with a DAD 1100 diode array detector. The solvents used for gradient elution were 10 mM phosphoric acid (pH, 2.5) and 100% methanol. The methanol concentration was increased to 60% for the first 8 minutes and to 100% over the next 7 minutes, then decreased to 0% for the next 3 minutes. It was maintained for the next 7 minutes (total run time, 25 minutes). The analytical column used was Agilent Zorbax SB-C18, 250×4.6 mm interior diameter, with packing material of 5-μm particle size at a flow rate of 1 mL/min at room temperature. During each run, the absorbance was recorded at 226 nm and 305 nm and the chromatogram was integrated by using the Agilent Chemstation enhanced integrator. Pure standards of gallic acid, catechin, epicatechin, quercetin, caffeine, and epigallocatechin in 100% methanol were used to calibrate the standard curves and retention times. The peaks were identified with consideration of the retention time and the ultraviolet absorption spectra of the corresponding standards.
Preparation of starter culture of H. pylori
H. pylori was cultured according to Stevenson et al. 36 Standard plating medium (H. pylori agar plates) were prepared by using 10 g of special peptone (Oxoid Ltd., Basingstoke, United Kingdom) per L, 15 g of granulated agar (Difco Laboratories, Becton-Dickinson, Sparks, MD, USA) per L, 5 g of sodium chloride (EM Science, Gibbstown, NJ, USA) per L, 5 g of yeast extract (Difco) per L, and 5 g of beef extract (Difco) per L of water.
Broth media were prepared by using 10 g of special peptone (Oxoid Ltd.) per L, 5 g of sodium chloride (EM Science) per L, 5 g of yeast extract (Difco) per L, and 5 g of beef extract (Difco) per L of water. A volume of 1 mL of frozen stock of H. pylori was added to test tubes containing 10 mL of sterile broth media. This was incubated at 37°C for 48 hours. Ten mL of sterile broth media was inoculated with 1 mL of the 48-hour culture and incubated at 37°C for 24 hours. The active culture was then spread on H. pylori agar plates to make a bacterial lawn for the agar-diffusion assay.
Agar-diffusion assay
The antimicrobial activity of the sample extracts was analyzed by using the agar-diffusion method. The assay was done aseptically using sterile 12.7-mm-diameter paper disks (Schleicher & Schuell, Inc., Keene, NH, USA) to which 100 μL of test extracts was added. Saturated disks were placed onto the surface of seeded agar plates. Controls consisted of disks with distilled water only. Treated plates were incubated at 37°C for 48 hours in BBL GasPak jars (Becton-Dickinson) with BD GasPak Campy container system sachets (Becton-Dickinson). The diameter of clear zone (no growth) surrounding each disk under static incubation with plates in GasPak jars was measured; the zone of inhibition was determined and expressed in mm. For the dose-dependency studies, 50 μL and 75 μL of the sample were used.
Proline growth response assay
A model for mode of action of phenolic phytochemicals was developed on the basis of the rationale that small phenolics could behave as proline analogues or proline analogue mimics and probably inhibit proline oxidation via proline dehydrogenase. 31 Furthermore, the likely inhibitory effects of phenolic phytochemicals should be overcome by proline if the site of action is proline dehydrogenase.
Bacterial lawns of H. pylori were prepared as described previously. Plating media were prepared by using standard plating medium as described for the agar-diffusion assay, with some modifications. Proline (Sigma, St. Louis, MO, USA) was added to the medium to a final concentration of 0.5 and 5 mM. Then, a protocol similar to that described for the agar-diffusion assay was followed.
Lactic acid bacterial proliferation assay
Initially, 100 μL of frozen stock from the lactic acid bacterial strains were inoculated into 10 mL de Man, Rogosa, Sharpe (MRS) broth (Difco) and incubated for 24 hours at 37°C. Then, 100 μL of the 24-hour strain was re-inoculated into 10 mL MRS broth for 24 hours at 37°C. Sample volumes of 1 mL were filter-sterilized by using Millex GP 0.22-μm filters (Millipore Corp., Bedford, MA, USA). Filter-sterilized sample extracts (1 mL) and 100 μL of the 24-hour strain with approximately 108 colony-forming units/mL (diluted 100 times with sterile distilled water) were added to 9 mL of MRS broth tubes and incubated at 37°C for 24 hours. A control with 1 mL of sterile distilled water instead of sample extract was also included. One hundred microliters of the serially diluted samples were plated in triplicates every 0, 6, 9, 12, and 24 hours on MRS agar (Difco) plates and incubated in anaerobic BBL GasPak jars (Becton-Dickinson) with BD GasPak EZ anaerobe container system sachets (Becton-Dickinson) at 37°C for 48 hours to determine the colony-forming units/mL.
Statistical analysis
All experiments were performed in duplicate, and the analysis at every time point from each experiment was carried out in triplicate (n=6). The HPLC experiments were performed at least in triplicate. Results are expressed as means±standard error. Data were subjected to the Student t-test using Microsoft Excel 2003 (Redmond, WA, USA) with P value at P<.05.
Results and Discussion
Extract absorbance analysis
The absorbance of the 2- and 5-minute extracts was measured at 400 and 600 nm. Table 1 presents results of the analysis. In general, black teas, as expected, had the highest absorbance, followed by partially fermented oolong teas; green tea and white tea had the lowest absorbance. The differences in absorbance did not indicate a correlation to phenolic profiles based on HPLC analysis (see discussion below) and total phenolics or antioxidant activity reported elsewhere.* In general, the differences reflect the state of oxidation—black teas (obtained from mature leaves) are considered the most oxidized teas, and white tea (obtained from leaf buds) the least oxidized.
HPLC analysis of phenolic profiles
HPLC analysis of the tea samples was performed to determine specific phenolic compounds present in the samples. Table 2 shows the amount of each compound detected by HPLC analysis in each sample with a 2- and 5-minute extraction. Gallic acid, quercetin, caffeine, and tea catechins (including catechin, epicatechin, and epigallocatechin) were found in all the tea samples. Epigallocatechin gallate and epicatechin gallate were not found in any of the samples evaluated. A strong correlation (Pearson correlation coefficient) was found between catechin (R=0.91), epicatechin (R=0.88), epigallocatechin (R=0.85), and the zone of inhibition in the samples that showed inhibition. However, even some samples that did not show inhibition showed the presence of these phenolic compounds. This finding suggests that these compounds may work in combination with other compounds that were not detected in the HPLC analysis in inhibiting H. pylori. Zhu et al. 37 have reported that the structures of tea catechins were not affected for 18 hours at a pH less than 4.0, which is similar to the pH that phenolic structures would encounter in the stomach. This indicates that phenolic structures involved in H. pylori inhibition will not be affected by stomach pH.
Data are expressed as means±standard error.
H. pylori inhibition and proline growth response assay
The potential of several commercial tea varieties and the influence of extraction time on H. pylori inhibition were evaluated. Of all the tea varieties tested with 5-minute extraction time, Choice darjeeling, Choice oolong, Choice white, Tazo white, and Tazo black showed inhibition. In contrast, with the 2-minute extraction time, only Choice darjeeling and Tazo white showed inhibition (Fig. 1a). Bigelow brand did not show any inhibition. The highest inhibition zone was observed for Choice darjeeling at 5 minutes (16.95 mm), whereas the lowest was observed for Tazo black at 5 minutes (14.68 mm). Significant difference (P<.05) was observed in H. pylori inhibition of the 2- and 5-minute sample for both Choice darjeeling and Tazo white, with 5-minute sample being superior.
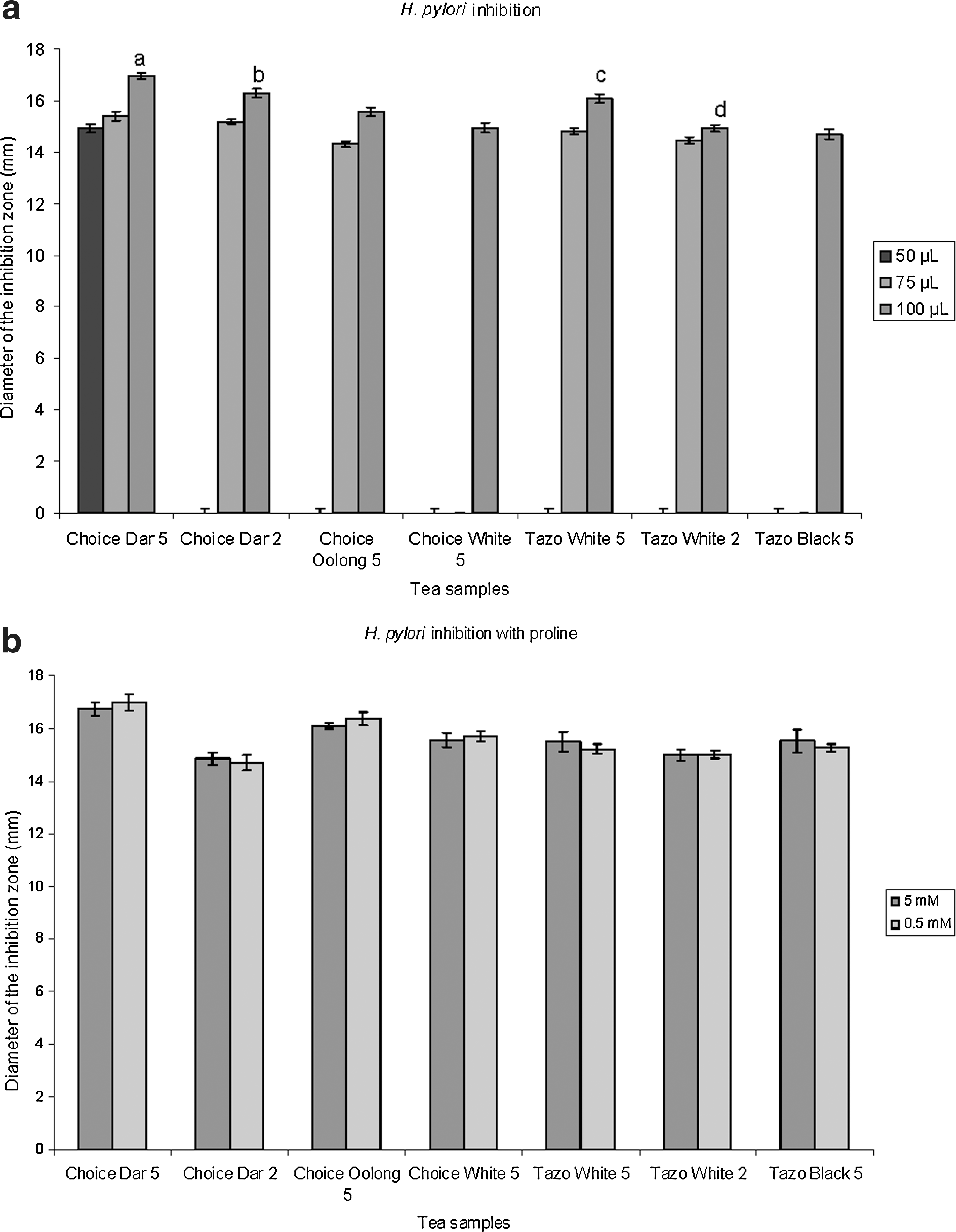
(
The proline growth response assay is based on the rationale that small phenolics in tea could behave as proline mimics and can inhibit proline oxidation via proline dehydrogenase at the plasma membrane level in prokaryotic cell, disrupting the oxidative phosphorylation-linked proton motive force and resulting in inhibition of the bacterium. 4 If so, addition of proline could overcome inhibition of the bacterium by proline's mimicking of phenolics present in the tea extract. The results obtained in our study (Fig. 1b) indicate that microaerophillic H. pylori inhibition by water extracts of tea is not associated with the proline dehydrogenase-based oxidative phosphorylation and that other mechanisms associated with membrane disruption or acidification could be involved.
Tea polyphenolic antioxidants might be able to quench electrons from electron transport chain along the bacterial membrane and act as antimicrobials by disrupting oxidative phosphorylation or inhibit dehydrogenase-linked proton efflux by interfering with the flow of electrons at the level of cytochromes. 4 However, simple soluble phenolics that are not very hydrophobic may not be effective in disrupting oxidative phosphorylation because of the external lipopolysaccharide layer of the gram-negative H. pylori, which creates a hydrophobic microenvironment along the bacterial surface. It is also thought that soluble phenolics can disrupt the H+-adenosine triphosphatase required for adenosine triphosphate synthesis by causing hyperacidification via proton donation at the plasma membrane and by intracellular cytosolic acidification. 31,33 Another explanation is that partial hydrophobicity of polyphenols in more fermented teas can attach to the cell wall, thereby allowing them to possibly stack themselves. The stacking, in turn, would cause destabilization of the membrane and lead to membrane disruption or transport inhibition. These mechanisms may work synergistically: i.e., a disrupting and destabilizing effect of the polyphenols may make it easier for the simple soluble phenolics to exert their hyperacidification.
Ikigai et al. 38 have reported that the bactericidal effect of catechins is by way of damage to bacterial membranes. Funatogawa et al. 13 showed this effect of tea catechins in H. pylori. Damage to DNA of gram-negative H. pylori could be an underlying mechanism involved in inhibition. Puupponen-Pimia et al., 39 with their studies on gram-negative Escherichia coli, reported that the reaction of phenolic compounds with DNA was the main cause of inhibition.
Polyphenols may show anti–H. pylori properties by inhibiting ureases. Matsubara et al. 18 reported inhibition of H. pylori ureases by green tea, oolong tea, jasmine tea, and black tea. In that study, both tea catechins and green tea extracts suppressed H. pylori–induced gastric lesions in Mongolian gerbils. Lin et al. 32 reported urease inhibition by phenolic phytochemicals in vitro. In contrast, Shin et al. 15 examined the effect of some flavonoids, including tea catechins, quercetin, and naringin, and reported no inhibitory effect on urease. In addition, catechins may restrict the supply of ions Fe3+ and Ni2+ essential for bacterial growth by forming complexes with these metal ions. 3,40 Overall, it is not clear from our studies whether the effects of tea extracts are bacteriostatic or bacteriocidal and what the precise mode of action is. Further studies based on these initial studies will clarify these mechanisms proposed on disruption of membrane functions and will form the foundation of yet further studies.
Probiotic lactic acid bacteria proliferation studies
Tea samples that showed inhibition against H. pylori were further investigated for their effects on lactic acid bacteria to determine relevance for positive health benefits. The concentrations of extracts used in this study were similar to those used in H. pylori inhibition studies. No inhibitory or stimulatory effects of the extracts were observed on probiotic strains of B. longum (Fig. 2a and Fig. 3a), L. plantarum (Fig. 2b and Fig. 3b), and L. acidophilus (Fig. 2c and Fig. 3c) as compared to control. This finding indicates that tea extracts can inhibit H. pylori without affecting the intestinal bacteria when extracted in a way typical of daily consumption.

(
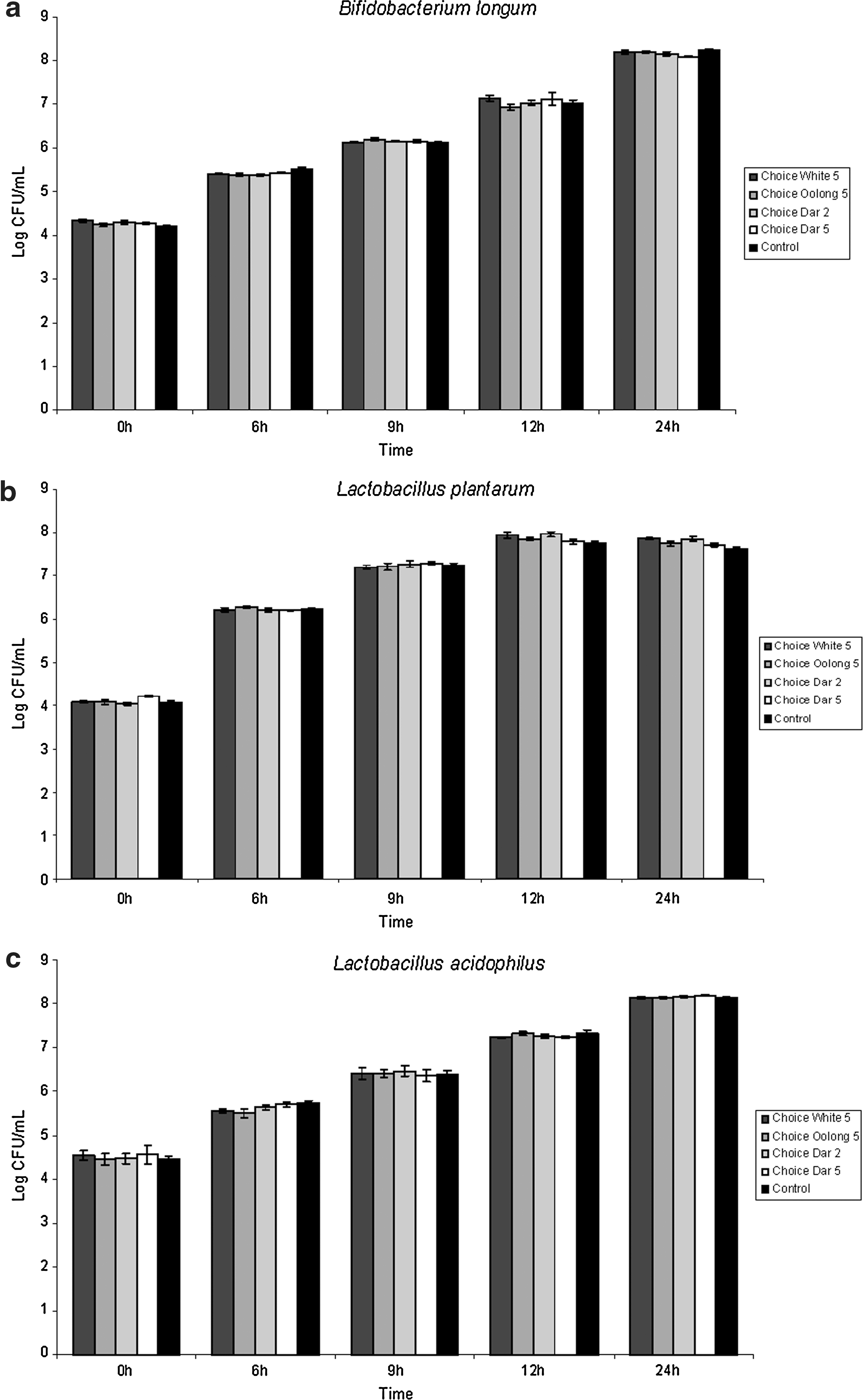
(
Funatogawa et al. 13 studied the effect of hydrolyzable tannins and reported that although these compounds could inhibit H. pylori, they did not affect E. coli, a normal inhabitant of the gut microflora. Results reported elsewhere suggest that tannins derived from Galla rhois inhibited pathogens but had a very slight inhibitory effect or no effect on intestinal flora. 25 Chung et al. 26 found no inhibition of lactic acid bacteria when tannic acid was used. They further indicated that this might be because of the absence of heme enzymes and the replacement of the crucial iron-containing ribonucleotide reductase with adenosylcobalamine in some lactic acid bacteria. Thus, although metal-chelating compounds would inhibit the growth of other bacteria by sequestering essential metal ions, it would have no effect on lactic acid bacteria. The growth of lactic acid bacteria has also been shown to be stimulated by extracts from Panax ginseng and green tea. 28,29 The results might have varied because the growth was evaluated by using methanol extracts as against water extracts in this study. By contrast, Horiba et al. 27 reported a 10-fold decrease in the number of L. acidophilus organisms in broth and a significant zone of inhibition for B. bifidum using agar diffusion assay with 20% methanol extract of green tea. Puupponen-Pimia et al. 39 discussed the structure–activity relationship and reported that lipophilic phenolics such as quercetin and kaempferol could not inhibit lactic acid bacteria, whereas hydrophilic phenolics such as myricetin were completely inhibitory. This might partly explain our findings of why H. pylori was inhibited but lactic acid bacteria were not. The difference in dependency on partial oxidative phosphorylation in microaerophilic bacteria such as H. pylori may be another reason for inhibition and lack of inhibition in lactic acid bacteria that use substrate level phosphorylation for energy generation.
Conclusion
The antimicrobial effects of tea extracts and the influence of extraction time were evaluated on the inhibition of H. pylori. Five-minute extraction was found to be superior for the in vitro inhibition of H. pylori linked to gastric diseases. Major phenolic compounds detected by HPLC analysis were gallic acid, quercetin, and tea catechins (including catechin, epicatechin, and epigallocatechin). Preliminary insights into the mechanism of action revealed that inhibition is not linked to proline dehydrogenase-based oxidative phosphorylation. Other mechanisms involving membrane disruption, DNA damage, or urease inhibition could be involved and will be explored in further studies. These tea extracts did not inhibit species of the beneficial lactic acid bacteria, including L. acidophilus, L. plantarum, and B. longum. This finding leads us to conclude that tea can be potentially used as a dietary support to manage H. pylori–linked infections without affecting the intestinal probiotic bacteria.
Author Disclosure Statement
None of the authors have any competing interests.
Footnotes
*
Ankolekar C, Terry T, Johnson KC, et al.: Anti-hyperglycemia properties of tea (Camellia sinensis): phenolic bioactives using in vitro assay models and influence of extraction time. Unpublished paper. University of Massachusetts, Amberst, MA, USA, 2011.