
Abstract
Flaxseed, rich in the phytoestrogen lignan secoisolariciresinol diglycoside (SDG), provides protection against bone loss at the lumbar vertebrae primarily when combined with low-dose estrogen therapy in the ovariectomized rat model of postmenopausal osteoporosis. Whether SDG metabolites are accessible to skeletal tissue, and thus have the potential to interact with low-dose estrogen therapy to exert direct local action on bone metabolism, is unknown. The objective of this study was to determine whether metabolites of SDG are accessible to the skeleton of ovariectomized rats and to compare the distribution of SDG metabolites in skeletal tissue with that in other tissues. Rats were fed a 10% flaxseed diet and gavaged daily with tritium-labeled SDG (7.4 kBq/g of body weight) in deionized water (500 μL) (n=3) or deionized water alone (n=3) for 7 days, after which tissues were collected for liquid scintillation counting. Radioactivity was detected in similar concentrations in the lumbar vertebrae, femurs, and tibias. Compared with non-skeletal tissues, total radioactivity in the skeleton was significantly lower than in the liver, heart, kidney, thymus, and brain (P<.001). There were no significant differences in levels of radioactivity between skeletal tissue versus the spleen, lung, bladder, uterus, vagina, and mammary gland. In conclusion, SDG metabolites are accessible to skeletal tissue of ovariectomized rats. Thus, it is biologically plausible that SDG metabolites may play a direct role in the protective effects of flaxseed combined with low-dose estrogen therapy against the loss of bone mass and bone strength in the ovariectomized rat model of postmenopausal osteoporosis.
Introduction
P
Dietary estrogens (i.e., phytoestrogens) such as isoflavones from soy and lignans from flaxseed are currently being investigated as a natural strategy to manage menopausal-related conditions. Our laboratory recently showed, using the established ovariectomized rat model of postmenopausal osteoporosis, that 10% dietary flaxseed, rich in the phytoestrogen lignan secoisolariciresinol diglycoside (SDG), is neither protective nor detrimental to bone mineral density (BMD) or bone strength. 7 It is interesting that a synergistic effect on BMD and bone strength at the lumbar vertebrae was observed when flaxseed was combined with low-dose estrogen therapy. 7 These findings are relevant to postmenopausal women, particularly those concerned with preserving bone health, as there is no ideal treatment for postmenopausal loss of BMD. Moreover, Western women who take prescription drugs such as hormone replacement therapy also commonly consume dietary supplements, many of which contain phytoestrogens. 8,9 Although estrogen therapy is not a mainline treatment for managing bone health, its usefulness in younger postmenopausal women is being re-evaluated, and it may be prescribed in the short term to help alleviate hot flushes and night sweats and to prevent bone loss that is associated with early menopause. 5,6 In addition, the number of food products containing flaxseed has tripled since 2003. 10,11 Examples of foods in which flaxseed has been introduced include breads, cereals, cereal bars, pasta, and yogurt. SDG is also found in appreciable amounts in vegetables, fruits, whole grains, and berries. 12,13 Consequently, the consumption of flaxseed and SDG is widespread in the Western diet.
The mechanisms whereby the combination of flaxseed and low-dose estrogen therapy exerts its greatest protection to bone mass and bone strength at the lumbar vertebrae are currently unknown but may be related to high levels of lignan SDG and the n-3 polyunsaturated fatty acid (PUFA) α-linolenic acid. Our laboratory has previously shown that feeding a 10% flaxseed diet to ovariectomized rats results in higher proportions of n-3 PUFAs, including α-linolenic acid and its longer-chain metabolite eicosapentaenoic acid, in lumbar vertebrae and tibias, 7 but it has not yet been shown whether SDG metabolites (secoisolariciresinol, enterodiol, enterolactone, and other minor metabolites 14,15 ) are accessible to skeletal tissue. If so, lignans may alter the action of low-dose estrogen therapy through direct action on bone cells. Previous studies have reported the deposition of SDG metabolites into various organs 16 –18 but not into the skeleton. Thus, the objective of this study was to determine, using the ovariectomized rat model of postmenopausal osteoporosis and radiolabeled SDG, whether SDG metabolites are accessible to the skeleton. Because the distribution of SDG metabolites varies among tissues, 16 –18 it is not known how the accessibility of SDG metabolites in skeletal tissue compares with that in other tissues. Thus, a secondary objective of this study was to compare the radioactive levels of SDG metabolites in skeletal tissue with those in other tissues (i.e., liver, uterus, vagina, mammary gland, thymus, spleen, kidney, lungs, bladder, heart, and gastrointestinal tissue). By determining whether SDG metabolites are accessible to skeletal tissue, this study will provide insight into whether it is possible that lignans can have a direct role in bone metabolism when flaxseed is combined with low-dose estrogen therapy.
Materials and Methods
Radiolabeling of SDG
Purified SDG was prepared as previously described. 19 The benzyl methylenes of the purified SDG were then labeled with tritium using a gas exchange method by Amersham International (Little Chalfont, United Kingdom), 16 resulting in specific radioactivity of 999 GBq/mol and a radiochemical purity of 98.5%. The benzyl methylene groups were chosen as the labeling site because tritium is more stable at the benzyl methylene groups in comparison with the benzene ring of SDG. 14,16 Moreover, the benzyl methylene groups remain intact during metabolism of SDG by colonic microflora to its metabolites, enterodiol and enterolactone. 14
Animals, diets, and experimental design
Three-month-old ovariectomized Sprague Dawley rats (n=6; mean body weight, 207±3 g) were purchased from Charles River Laboratories (St. Constant, Canada), housed one per cage. and maintained at 22–24°C and 50% humidity with 12:12-hour light–dark cycle. Rats were acclimatized to the basal diet, an AIN-93M 20 formulation (Table 1), for the first 7 days. After this 7-day period, rats were switched to the flaxseed diet, which was the basal diet supplemented with 10% freshly ground flaxseed (Johnson Seeds Ltd., Arborg, Canada) (Table 1) and received a daily 500-μL gavage of tritium-labeled SDG (3H-SDG) (7.4 kBq/g of body weight) in deionized water (n=3) or deionized water alone (n=3). The dose of 3H-SDG represents a level that was previously used by Saarinen et al. 18 and can be detected in rodent tissues without toxicity. This dose of 3H-SDG also contributes a negligible amount of SDG (<1%) to the whole flaxseed diet. 16 The dose of flaxseed chosen for this research is a level that can be attained in the human diet (i.e., 25–50 g/day) as whole or ground seed incorporated into foods such as muffins and cereals. 21 –23 At the end of the 7-day intervention, rats were killed 12 hours following the last oral gavage, and skeletal samples (i.e., lumbar vertebrae, femur, and tibia) and non-skeletal samples (i.e., serum, liver, kidney, bladder, spleen, brain, heart, mammary, uterus, vagina, thymus, and lungs) including gastrointestinal tissues (i.e., stomach, small intestines, cecum, and colon) were dissected, rinsed with phosphate-buffered saline, and frozen at –80°C for later analyses. The time point chosen for sacrifice was based on previous research that showed high levels of radioactivity in tissue and blood at this time after dosing. 16 Animal care and use were carried out in accordance with the Guide to the Care and Use of Experimental Animals, 24 and the experimental protocol was approved by the Animal Ethics Committee at the University of Toronto, Toronto, Canada.
Secoisolariciresinol content, 178.73±62.06 mg/100 g of flaxseed; α-linolenic acid content, 23.55±0.74 g/100 g of flaxseed.
Determination of radioactivity in tissues and serum
Determination of radioactivity in tissues and serum was conducted in both the 3H-SDG group and the control group that received deionized water alone. For scintillation counting of skeletal tissue (n=3 per skeletal site), bone marrow was flushed from femurs and tibias with phosphate-buffered saline. Lumbar vertebrae (200–280 mg), femurs (325–600 mg), and tibias (325–500 mg) were subsequently rinsed three times with phosphate-buffered saline to remove residual blood. Individual bones were dipped in liquid nitrogen and crushed with a hammer, and large bone masses were then pulverized with a mortar and pestle under liquid nitrogen as previously described. 7 Individual samples were then dissolved in a mixture of 1 M hyamine hydroxide in methanol (Packard Bioscience B.V., Groningen, The Netherlands) and deionized water (9:1 vol/vol) and were incubated overnight in a shaking water bath at 45°C. Fifteen milliliters of scintillant (Cytoscint ES, ICN Biomedicals, Costa Mesa, CA, USA) was then added to the dissolved tissue samples, and the samples were mixed vigorously. To minimize chemiluminescence, 0.5 mL of 0.5 M glacial acetic acid was added to each vial. Vials were then shaken and allowed to equilibrate in the dark for 4 days before scintillation counting.
For scintillation counting of non-skeletal tissue, samples were processed according to the methods described earlier. 16 –18 Samples (80–160 mg) were minced and dissolved in a mixture of 1 M hyamine hydroxide in methanol (Packard Bioscience B.V.) and deionized water (9:1 vol/vol) and were incubated overnight in a shaking water bath at 45°C. Dissolved samples (100 μL) were then mixed with 5 mL of scintillant. For tissues with residual blood, 100 μL of 9.7 M hydrogen peroxide was added, followed by heating the samples at 55–60°C for 30 minutes with the vials loosely capped. Once cooled, 0.5 mL of 0.5 M glacial acetic acid was added to the samples to minimize chemiluminescence, and the vials were vigorously mixed before being stored in the dark for 4 days before scintillation counting.
For scintillation counting of serum, 50 μL of serum was dissolved in 300 μL of hyamine hydroxide (Packard Bioscience B.V.) in ethanol (1:2, vol/vol). Samples were then incubated in a shaking water bath at 55–60°C for 1 hour, after which 0.5 mL of 9.7 M hydrogen peroxide was added to prevent color quenching. Vials were incubated for another 30 minutes at 55–60°C and then cooled before addition of scintillant and glacial acetic acid to minimize chemiluminescence. Vials were then mixed vigorously and stored in the dark for 4 days before scintillation counting. 16
All samples were counted for 5-minute periods using a TRI-CARB 2900 TR liquid scintillation analyzer (Packard Instrument Co., Meriden, CT, USA) with a mean counting efficiency of 63.34% for tritium based on unquenched controls. Corrections for chemical and color quenching of the samples were done automatically by the liquid scintillation counter. All results were corrected for tissue-specific counting efficiencies and possible losses during processing as previously described. 16 All results were converted to picomole equivalents of 3H-SDG by dividing them by the specific activity of the radioisotope (27 mCi/mL) and were adjusted for the weight or volume of tissues and serum.
Statistical analyses
To detect differences in radioactivity within a particular tissue between the control and 3H-SDG groups, an unpaired Student's t test was performed. Differences in radioactivity among various tissues within a group (i.e., control or 3H-SDG-treated rats) were determined by one-way analysis of variance followed by a Student–Newman–Keul's test as the post hoc test for data that were normally distributed and had equal variances. For data that were not normally distributed (gastrointestinal tissues) or did not have equal variances (non-skeletal tissue), a Kruskal–Wallis one-way analysis of variance followed by Dunn's post hoc test was performed. All statistics were performed using SigmaStat (version 3.5, Systat Software Inc., Chicago, IL, USA). The significance level was set at P<.05. All data are presented as mean±SEM radioactivity per gram of tissue weight.
Results
Final body and organ weights
There were no significant differences in body weight between the control and 3H-SDG groups at either the beginning (control, 206±7 g; 3H-SDG, 207±3 g) or end (control, 215±10 g; 3H-SDG, 216±4 g) of the 1-week study. Final tissue weights were similar between the control and 3H-SDG groups (data not shown).
Radioactivity in tissues and serum
Administration of 3H-SDG resulted in detectable radioactivity in all collected tissues (Fig. 1A). Radioactivity in gastrointestinal tissues (i.e., cecum, colon, stomach, and small intestine) represented 65.2% of the total radioactivity detected, which was primarily due to concentrations of radioactivity in the cecum and colon (Fig. 1A).
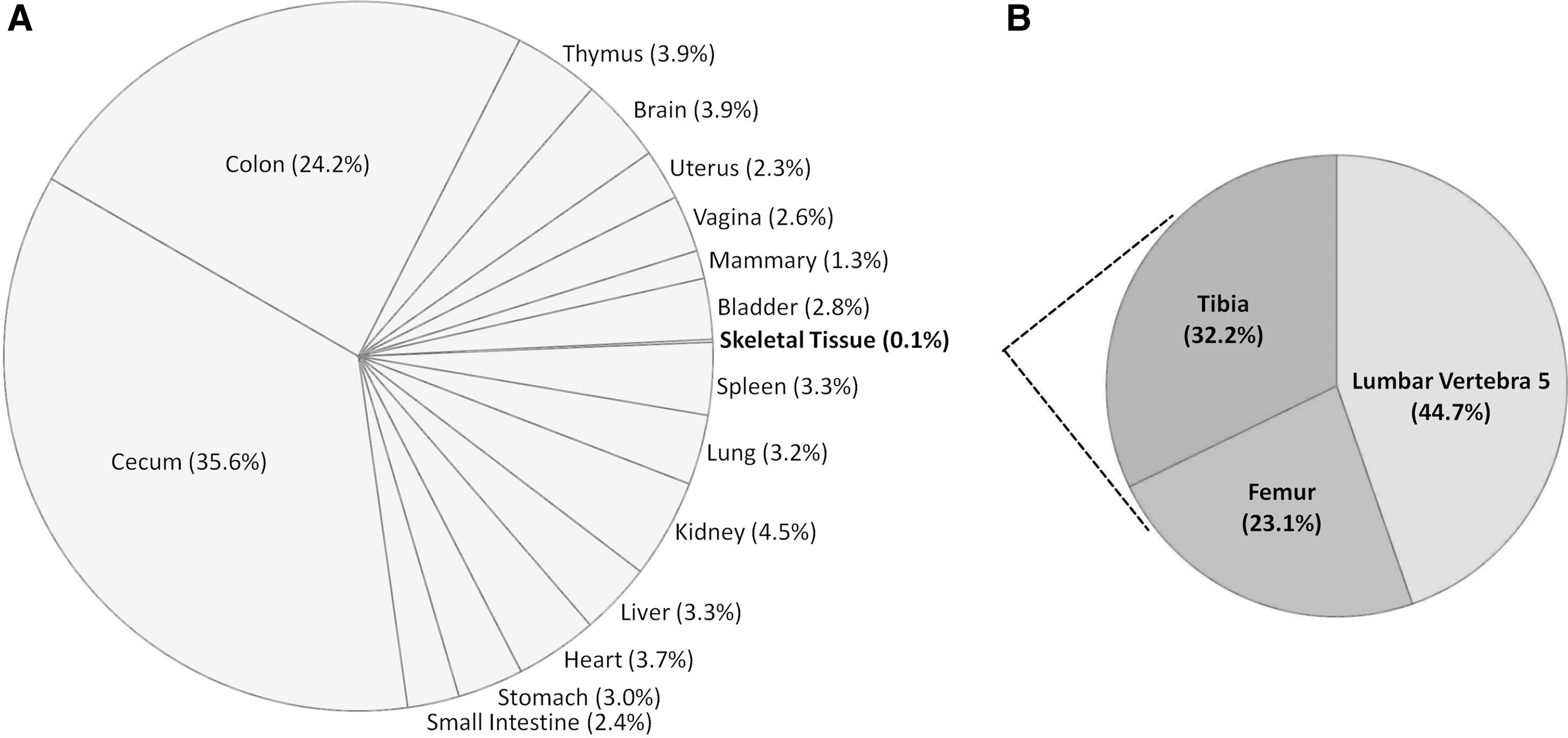
Percentage distribution of 3H-secoisolariciresinol diglycoside metabolites in (
In non-gastrointestinal tissues, kidney had highest level of radioactivity (4.5% of the total measured radioactivity), whereas skeletal tissue levels had the lowest (0.1% of total radioactivity) (Fig. 1A). In skeletal tissue, radioactivity in the lumbar vertebra, tibia, and femur represented 44.7%, 32.2%, and 23.1% of the total radioactivity measured in the skeleton, respectively (Fig. 1B). Overall, the radioactivity concentrations from highest to lowest were cecum>colon>kidney>thymus>brain>heart>spleen>liver>lung>stomach>bladder>vagina>small intestine>uterus>mammary>lumbar vertebra>tibia>femur. As expected, no radioactivity was detected in the tissues of control rats that received deionized water alone.
Radioactivity in the lumbar vertebra was 1.4 and 1.9 times higher than in the tibia and femur, respectively, but they did not differ significantly (Fig. 2A). Radioactivity in the total skeletal tissue was significantly lower than the thymus, heart, brain, liver, and kidney (P<.001) (Fig. 2B). No differences in radioactivity were observed between total skeletal tissue and mammary, uterus, vagina, spleen, bladder, and lung tissues (Fig. 2B). Excluding skeletal tissue, the highest level of radioactivity was observed in the kidney, which was 1.1–3.5 times higher than all other non-gastrointestinal tissues. Radioactivity in the kidney was significantly higher compared with mammary tissues (P<.001), but it did not differ significantly from other non-gastrointestinal tissues (i.e., thymus, brain, uterus, vagina, bladder, spleen, lungs, liver, and heart) (Fig. 2B). Cecum radioactivity was 1.5–15.1 times higher than in other gastrointestinal tissues (colon, stomach, and small intestine). Radioactivity in the cecum and colon were significantly higher than in the small intestine (P<.001), but these values did not differ from each other (Fig. 2C). Radioactivity of the stomach did not differ from that of other gastrointestinal tissues (Fig. 2C). The radioactivity in serum was 7.8±0.51 pmol/mL. In all tissues combined, the percentage of radioactivity recovered from the total administered dose was 5.6±1.0%, mainly due to the radioactivity present in the whole liver, cecum, and colon (1.1–1.3% of the administered dose), whereas the radioactivity recovered in all other whole tissues was below 1% of the total administered dose.
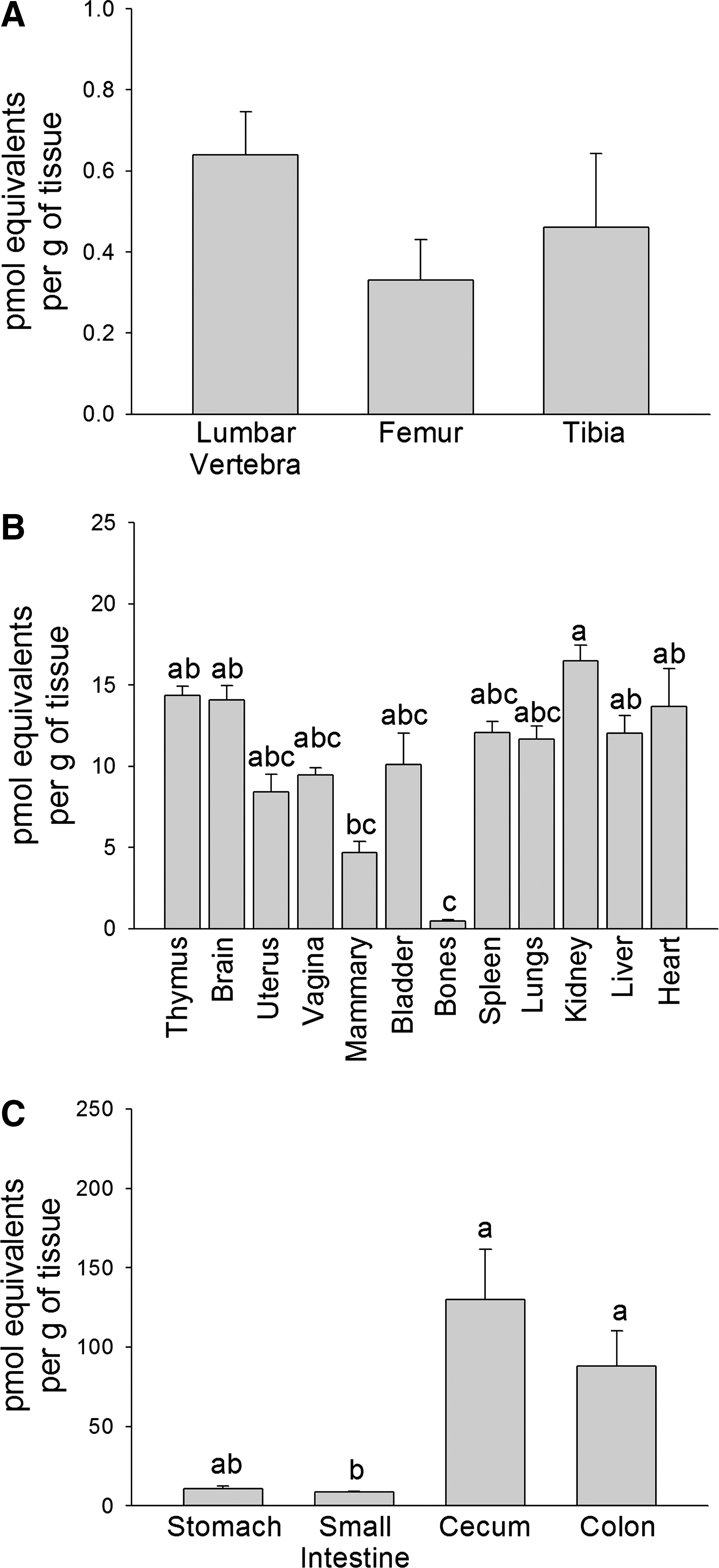
Mean levels of radioactivity in (
Discussion
This study has shown for the first time that SDG metabolites are accessible to skeletal tissue (i.e., lumbar vertebrae, femurs, and tibias) in the ovariectomized rat model of postmenopausal osteoporosis. Although SDG metabolites are accessible to skeletal tissue and thus may have the potential to exert direct action on skeletal metabolism when combined with low-dose estrogen therapy, it is unknown which metabolites (i.e., secoisolariciresinol, enterodiol, enterolactone, etc.) may be responsible for these potential effects, and they are present at much lower levels than in most other tissues. This present study distinguishes itself from previous work because it is the first to determine whether SDG metabolites can be incorporated into skeletal tissue. Moreover, the current study was the first of its kind to administer 3H-SDG in combination with a 10% flaxseed diet instead of unlabeled SDG, which is more reflective of the human diet.
Flaxseed and the metabolites of its major lignan (SDG) and n-3 PUFA (α-linolenic acid), have been shown to favorably modulate skeletal metabolism. 25 –27 The up-regulation of proteins involved with bone formation (i.e., osteonectin, alkaline phosphatase, and collagen I) was observed in MG-63 osteoblastic-like cells treated with the SDG metabolites enterolactone and enterodiol. In contrast, epidemiologic, in vivo, and clinical studies have reported conflicting findings on the effectiveness of flaxseed or its purified lignans on bone metabolism, particularly in the postmenopausal situation. 7,21,28 –31 Variability in experimental systems (i.e., animal vs. humans; healthy adult rats vs. ovariectomized rats representing a postmenopausal osteoporotic situation) may explain the conflicting findings regarding the effectiveness of flaxseed or its components on bone metabolism. Even so, the only previous study investigating the effect of flaxseed on bone metabolism in an ovariectomized rat model of postmenopausal osteoporosis observed that flaxseed alone does not modulate BMD or bone strength. 7 It is interesting that the greatest benefit to vertebral BMD and strength was observed when flaxseed was combined with low-dose estrogen therapy. 7 These findings suggest that even though flaxseed lignans and n-3 PUFAs are accessible to skeletal tissue, their biological potency and level are not sufficient to induce a functional response on BMD or bone strength in ovariectomized animals when administered alone.
Whether the amount of SDG metabolites that reached the skeletal tissue is sufficient to modulate bone cell function when combined with low-dose estrogen therapy requires further investigation. The level of radioactivity observed in the skeletal tissue analyzed in the current study represents 0.035% of the daily administered radioactivity. Based on previous research that has shown that ovariectomized rats consume approximately 18 g of food/day 7 and on the amount of secoisolariciresinol aglycone present in our 10% flaxseed diet (178.73 mg/kg of food) used for this study, 7 we calculate that approximately 3.22 mg of secoisolariciresinol aglycone was consumed daily by the rats in the present study, resulting in the accessibility of secoisolariciresinol aglycone metabolites to skeletal tissue at a level of 7.08 μg/g of tissue throughout the 7-day study.
Although the mechanisms whereby flaxseed combined with low-dose estrogen therapy preserves bone tissue in the ovariectomized rat model of postmenopausal osteoporosis are unknown, findings from the present study suggest that it may be possible for lignan metabolites to exert direct local action on bone metabolism. Like estrogen, the mammalian lignans (enterolactone and enterodiol) have the ability to bind to estrogen receptors, with preferential binding to estrogen receptor-α. 32 Estrogen receptor-α has been identified in osteoblasts, osteoclasts, and their precursors; it regulates the transcription of several genes involved with bone turnover (osteoprotegerin and macrophage colony-stimulating factor). 33 However, enterolactone can also compete with estrogen for binding to the estrogen receptor and thus may interfere with the effect of estrogen on skeletal metabolism. 34 It is important to note in the present study that radioactive levels of SDG metabolites in skeletal tissue were relatively low, representing 4.0–8.2% of the concentration detected in the serum. This finding is similar to previous studies investigating the tissue distribution of selective estrogen receptor modulators such as tamoxifen and raloxifene in which lower or minimal levels of these selective estrogen receptor modulators are reported in skeletal tissue compared with other selected tissues. 35,36
In the present study, a small percentage of the administered radioactivity was found in tissues, which agrees with previous work that has shown that after 12 hours of 3H-SDG administration, most radioactivity is present in the cecal and colon contents. 16 In contrast to others, 16,17 the current study observed that the level of 3H-SDG metabolites in the serum was generally lower than those in most tissues, excluding the skeletal and mammary tissues. These findings conflict with a previous study in which serum radioactivity was higher compared with all other tissues in 8-week-old intact female and male rats that were gavaged for 1 or 7 days with 3H-SDG and unlabeled SDG. 17 Discrepancies between our findings with the previous study 17 may be due to the level of 3H-SDG gavaged in the aforementioned study (3.7 kBq/g of body weight), which is half of the dose that was used in the present study (7.4 kBq/g of body weight). We specifically chose to give a previously studied dose 18 that was higher than that used by Saarinen and Thompson 17 to ensure that skeletal incorporation could be measured if indeed this did occur. In addition, unlabeled SDG (5.3 μg/g of body weight) in the previous study was gavaged to reflect the amount of SDG present in a 5% flaxseed diet, 17 which contrasts to the present study, in which a 10% whole flaxseed diet was administered. Thus, the higher dose of 3H-SDG and SDG from the 10% flaxseed diet in the current study, along with other lignans (i.e., matairesinol, lariciresinol, and pinoresinol) present in whole flaxseed, could have resulted in greater intestinal absorption of 3H-SDG metabolites and radioactivity uptake in the tissues compared with serum. In fact, tissue levels of radioactivity in the current study are four- to eightfold higher than those reported in the aforementioned study 17 with a similar distribution of radioactivity in commonly measured tissues. This suggests that tissues of both adult intact and ovariectomized female rats respond similarly to lignan intake. The previous study, in which both unlabeled and 3H-SDG were gavaged daily for 1 week in female adult rats, measured radioactivity only in non-skeletal tissues. 17
Tritium labeling of SDG on the benzyl methylene groups as opposed to the phenolic groups was performed because tritium is more stable at this site and the benzyl methylene groups are present only in known SDG metabolites. 16 Previous research using scintillation counting of high-performance liquid chromatography fractions of urine from rats fed 3H-SDG demonstrated that the radioactivity administered was only detected at the retention times for enterodiol, enterolactone, secoisolariciresinol, and their other minor metabolites, 14 which provides evidence that no hydrogen exchange occurred. Future study using scintillation counting of high-performance liquid chromatography fractions from skeletal tissue is needed to determine whether this is also true in the bone.
In conclusion, our study has shown for the first time that SDG metabolites are accessible to the lumbar vertebrae, femurs, and tibias of ovariectomized rats fed a 10% flaxseed diet while being gavaged daily with 3H-SDG. However, whether SDG metabolites can exert direct or indirect action on bone cells and thus may play a role in the mechanism whereby flaxseed and low-dose estrogen therapy exerts a synergistic action on bone mass and strength in the ovariectomized rat model of postmenopausal osteoporosis requires further study.
Footnotes
Acknowledgments
We extend our gratitude to the Natural Sciences and Engineering Research Council of Canada for funding through a Discovery Grant to W.E.W. and a Doctoral Post-Graduate Scholarship to S.M.S. We thank Brian Johnson of Johnson Seeds Ltd. for generously providing the flaxseed for this study and Dr. Niina Saarinen for her helpful technical guidance.
Author Disclosure Statement
No competing financial interests exist.