
Abstract
Effects of dietary fibers on human postprandial energetics remain undefined. The objective of the present study was to explore effects of whole yellow pea flour (WPF) and fractionated pea flour (FPF) on postprandial energy expenditure, substrate utilization, and hepatic triglyceride synthesis rate. Using a crossover-diet controlled design, 23 overweight men and women received muffins containing WPF, FPF, and white wheat flour (WF) for 28 days, followed by 28-day washout periods. Subjects received 50 g/day WPF and WF. Given that FPF is approximately 84% fiber, the amount of FPF administered to volunteers was equivalent to the amount of pea-derived fiber in the WPF treatment. Four weeks of FPF consumption reduced (P=.007) total postprandial energy expenditure (333.0±3.6 kcal/330 minutes) compared with WF (349.3±3.6 kcal/330 minutes). When values were normalized to the level of food energy consumed, FPF (4.6±0.3%) decreased (P=.018) the thermic effect of food (TEF) compared with WF (5.7±0.3%). Carbohydrate oxidation tended to be lower (P=.075) with FPF (44.7±2.1 g/330 minutes) versus WF (51.2±0.1.9 g/330 minutes). WPF had no effect on total energy expenditure, TEF, or carbohydrate oxidation. Only after 370 minutes was cumulative oxidation of [1-13C]palmitic acid higher (P=.045) in the WPF group (0.96±0.05%) compared with FPF (0.81±0.05%). Neither treatment had any effect on hepatic triglyceride synthesis rate. In conclusion, chronic ingestion of different fractions of yellow peas imposes distinctive effects on postprandial energy expenditure and substrate utilization.
Introduction
D
Currently the U.S. Department of Agriculture recommends that ½ cup of pulses be consumed daily as part of a healthy diet. 5 However, recommended levels of intake are difficult to attain because, compared with wheat, pulse crops are rarely used domestically and by the food industry as ingredients for formulating novel food products. Poor variety and unfavorable sensory characteristics that are unappealing to certain populations, especially in North America, have limited extensive integration of pulse crops into foods. 6 However, with the development of pulse-derived flours, novel functional foods that use pulses as the primary ingredient can be seamlessly integrated into diets devoid of pulse crops.
Although whole yellow peas (Pisum sativum L.) are one of the most widely cultivated pulse crops, the health benefits of whole yellow peas and their fractions remain largely undefined. Given that yellow peas provide a good source of protein and fiber, their incorporation into food products may increase or decrease postprandial energy expenditure. Investigating and identifying the thermogenic effects of functional foods that incorporate whole and fractionated yellow peas as the foremost ingredient could provide cost-effective options for harnessing the health benefits associated with pulse consumption.
The objectives of the present study were to investigate the effects of whole yellow pea flour (WPF) or fractionated yellow pea flour (FPF) on postprandial energy expenditure, substrate utilization, meal-derived fat oxidation, and hepatic triglyceride (TG) synthesis.
Materials and Methods
Subjects
Twenty-nine hypercholesterolemic, overweight men (body mass index 25–40 kg/m2) and women (body mass index 24–38 kg/m2) residing in the city of Winnipeg, MB, Canada, were recruited using radio and newspaper advertisements. Baseline characteristics of study subjects are summarized in Table 1. Exclusion criteria included smoking, use of prescription and natural lipid-lowering therapies, history of myocardial infarction, coronary artery bypass, angina, congestive heart failure, inflammatory bowel disease, pancreatitis, renal disease, diabetes, and a history of chronic alcohol usage (>2 drinks/day). Individuals with cancer or eating disorders and those who reported expending over 4,000 kcal/week through exercise were also excluded from participation. Participants taking medications for other ailments were included providing their daily dose of medication remained stable for the duration of the study. Prior to initiating the study, all subjects read and signed an informed consent form. This study was approved by the University of Manitoba's Biomedical Research Ethics Board.
n=23. Data are mean±SD values.
BMI, body mass index; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; TC, total cholesterol, TG, triglycerides.
Treatments
Treatments consisted of WPF, FPF (hulls only), and white wheat flour (control) (WF). Treatment flours were administered to subjects in muffins. Muffin recipes were developed at the Richardson Centre for Functional Foods and Nutraceuticals clinical kitchen. WPF and FPF flours were obtained from Best Cooking Pulses Inc. (Portage la Prairie, MB, Canada), while the WF was obtained from a local supermarket. Pea treatment flours were analyzed by Medallion Labs (Minneapolis, MN, USA) to determine their nutrient composition (Table 2). The composition of WF was determined by available food analysis tables. 7 Current U.S. Department of Agriculture recommendations for pulse consumption are ½ cup/day, which is approximately 50 g of dry whole yellow peas. 5 Given that each WPF treatment muffin was formulated to contain approximately 25 g of WPF, subjects were required to eat two treatment muffins per day. The FPF treatment muffins were formulated based on total pea-derived fiber content of the WPF muffin because the pea hull flour used in the present study was approximately 84.2% fiber (Table 2). Hence, the amount of fractionated flour added to the FPF muffin recipe was adjusted to equal the amount of fiber in the WPF treatment (Table 2); the balance of the muffin weight was WF. The WF muffin was solely composed of WF.
Treatment flours were analyzed at Medallion Labs.
Flour composition determined using available nutritional tables. 7
CHO, carbohydrate; WPF, whole yellow pea four; FPF, fractionated yellow pea flour; WF, white wheat flour.
Study design
The present study was a triple crossover, controlled-diet design with three 28-day study phases, each followed by a 28-day washout. In addition, the present study was single-blinded whereby volunteers were not informed as to which treatment they were receiving during a given treatment phase. To determine the treatment order within each subject, participants were randomly assigned to one of the six possible treatment sequences.
Diet and treatment protocol
For the duration of each study phase, subjects were instructed to consume only food provided by the Richardson Centre for Functional Foods and Nutraceuticals clinical research kitchen. The nutrient composition of the study diet was analyzed using Food Processor version 7.71 software (ESHA Research, Salem, OR, USA). The background diet had a nutrient composition of 55% CHO, 15% protein, and 30% fat. Diets were designed to contain no pulse products besides the treatment muffins. To prevent body weight change for the duration of each study phase, daily energy requirements for each subject were determined by calculating individual resting metabolic rates using the Mifflin equation 8 and then applying an activity factor of 1.7. The amount of food each participant received was based on individual calculated energy requirements. The average caloric content of two treatment muffins (approximately 400 kcal) was subtracted from the total calculated energy requirement to compensate for the energy intake from the treatments. Body weights were assessed daily. If a subject's weight increased or decreased during the first week of each study phase, energy intake was adjusted accordingly. Each morning, study participants were required to visit the Richardson Centre for Functional Foods and Nutraceuticals Clinical Research Unit to consume breakfast and one treatment muffin under supervision. Participants would then leave the unit with a cooler containing the remainder of their meals and snacks alongside a treatment muffin to consume with dinner. Empty, unwashed food containers were used to monitor compliance.
Energy expenditure protocol
On a single day during the first and last week of each study phase, energy expenditure was determined using open circuit indirect calorimetry (Vmax Encore, Summit Technologies Inc., Burlington, ON, Canada) fitted with a ventilated canopy. The flow sensor and gas sensors were calibrated daily prior to initiating respiratory measurements. The flow sensor was calibrated using a calibration syringe. Gas sensors were automatically calibrated by Vmax Encore software (Summit Technologies Inc.) using two reference gasses, with the first containing 16% O2, 4% CO2, and 80% N2 and the second containing 26% O2, 0% CO2, and 74% N2. All indirect calorimetry data was collected using Vmax Encore software. Finally, commencement of premenopausal women into the study was planned such that they did not undergo indirect calorimetry measurements during their menstrual cycle.
After a 12-hour fast, subjects arrived at the Richardson Centre for Functional Foods and Nutraceuticals. After being allowed to rest for a period of 20 minutes, subjects were comfortably installed in a bed in a supine position. The ventilated hood was placed over the subject's head, and resting energy expenditure (REE) was measured for 30 minutes prior to their breakfast meal. Subjects were given 40 minutes to consume their calorically controlled breakfast accompanied by one treatment muffin. In addition, the breakfast corresponding to day 1 of the 3-day menu cycle was always consumed on the day that subjects underwent energy expenditure measurement (Table 3). Volunteers were instructed to consume the treatment muffins prior to ingesting any other foods in the breakfast meal. Postprandial energy expenditure was measured in 30-minute increments immediately following breakfast and again 60, 120, 180, 240, and 300 minutes after the breakfast meal. For all measurements, subjects remained motionless in bed in the supine position. At times when energy expenditure was not being measured subjects were required to remain in the supine position. Washroom breaks were permitted during the break periods.
For each time increment, only respiratory gas exchange values recorded during the last 15 minutes of each measurement period were used to calculate TEE (in kcal/min), CHO oxidation (in g/min), and fat oxidation (in g/min). TEE was calculated using equations described by Lusk:
9
where 4.686 represents the kcal/L of O2 consumed with 100% fat oxidation, 0.707 is the respiratory quotient (RQ) (RQ=VCO2/VO2) corresponding to 100% fat oxidation, 0.293 corresponds to the difference in RQ between 100% CHO oxidation and 100% fat oxidation, and 0.361 is the difference in Kcal/L of O2 between 100% CHO oxidation and 100% fat oxidation. Non-protein (np) RQ was calculated based on the assumption that nitrogen is excreted at a constant rate of 0.14 g/kg of body weight/24 hours:
10
where 6.03 and 4.88 represent the volume (in L) of CO2 and O2 consumed per gram of nitrogen metabolized.
11
CHO and fat oxidation were calculated using the following equations:
where 0.746 and 2.019 represent the volume of O2 (in L) consumed per gram of glucose and TG oxidized, respectively. 12,13
Incremental area under the curve (AUC) values were calculated for each interval using the trapezoidal rule. TEE and total CHO and fat oxidation represented the total AUC. The TEF was calculated by subtracting the area below REE from the total AUC.
Determination of dietary fat oxidation
During week 4, dietary fat oxidation was assessed on the same day as the indirect calorimetry session. Immediately following the measurement of REE, a baseline breath sample was collected into 12-mL evacuated glass tubes (Exetainer®, Labco, High Wycombe, United Kingdom) using breath collection bags fitted with a gas collection port (EasySampler™, Quintron Instrument Co., Milwaukee, WI, USA). During the breakfast meal, subjects consumed warm margarine containing [1-13C]palmitic acid (CAS 57677-53-9, Cambridge Isotope Laboratories Inc., Andover, MA, USA), at 10 mg/kg of body weight, which was spread over the treatment muffin. Additional breath samples were taken immediately following the breakfast meal and again at 30, 90, 150, 210, 270, and 330 minutes following each indirect calorimetric session.
Breath samples were analyzed using continuous flow isotope ratio mass spectrometry (IRMS) (ABCA, SerCon Ltd., Crewe, UK) and analyzed using ABCA breath analyzer software version 500.1.12 (SerCon Ltd.). The IRMS instrument was calibrated against a reference gas containing 5% CO2. Samples were normalized against Pee Dee Belemnite limestone (PDB) where 13C/12C=0.0112372. 13C enrichment in the sample was calculated as: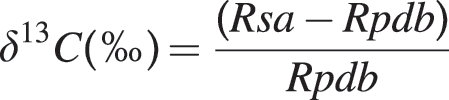
where δ13C represents the parts per thousand 13C/12C ratio of the sample relative to Rpdb, Rsa is the 13C/12C ratio in the sample, and Rpdb is the constant ratio of PDB.
14
The 13C atom percent (AP13C) was calculated as:
14
Atom percent excess (APE) was calculated as the difference between AP13C at each time interval and AP13C at baseline.
14
Percentage dose recovered per minute (PDR) was determined using the following equation:
15
where mmol of CO2 expired/minute was calculated multiplying mmol of CO2=22.7 mmol/L by the volume of CO2 expired/min as determined by indirect calorimetry.
15
The amount of 13C administered (mmol) was calculated using the following formula:
15
where 0.99 accounts for 99% labeling efficiency of 1-13C in administered palmitic acid, while 0.01 accounts for the 1% of naturally occurring 13C in administered palmitic acid. AUC of PDR was calculated using the trapezoidal rule and represents cumulative oxidation of dietary [1-13C]palmitic acid.
Determination of endogenous hepatic TG synthesis
On day 28 of each study phase, immediately following a fasted blood sample, subjects ingested 1.2 g/kg of body water deuterium oxide (APE 99.9%) (Cambridge Isotopes Inc.; obtained from ACP Chemicals Inc., Montreal, PQ, Canada). Body water was considered to be 70% total body weight. Deuterium oxide was microfiltered prior to ingestion. On the morning of day 29, approximately 24 hours after subjects ingested deuterium oxide, another fasted blood sample was taken. The exact time of deuterium ingestion and the day 29 blood sample was recorded.
Because very low-density lipoprotein (VLDL) particles are the richest source of newly synthesized fatty acids, VLDL-derived palmitic acid was used as an indicator for endogenous hepatic fatty acid synthesis. 16 VLDL from day 28 and 29 blood samples was separated from plasma using ultracentrifugation. In brief, 1 mL of plasma was added to 3 mL of saline (d=1.006 g/mL) in 4-mL polycarbonate ultracentrifuge tubes (Beckman-Coulter, Mississauga, ON, Canada). Samples were centrifuged at 306,000 g (RCFmax) for 4 hours. The VLDL was collected from the supernatant, and heptadecanoic acid was added to VLDL as an internal standard. VLDL-derived lipids were extracted using chloroform:hexane (4:1 vol/vol). Samples were dried under nitrogen and derivatized via methylation using boron trifluroide (Sigma Aldrich, Oakville, ON, Canada), hexane, and methanol (7:6:7 by volume) at 100°C for 55 min. Hexane and double distilled water were added to samples, which were then vortex-mixed and centrifuged. The supernatant was collected, dried under nitrogen, resuspended in hexane, and transferred to a gas chromatography vial for analysis.
Deuterium enrichment of methyl-palmitate was used to determine the fractional synthesis rate (FSR) of TG over a 24-hour period and was measured using gas chromatography (model 6890 chromatograph, Agilent Technologies, Palo Alto, CA, USA) fitted with a SAC-5 column (Sigma Aldrich) and coupled to a pyrolysis reactor and a Delta V Plus IRMS instrument (Thermo Fisher Scientific Inc., Bremen, Germany). After separation by gas chromatography, fatty acids were pyrolyzed at 1,450°C. The 2H/1H enrichment of methyl-palmitate was determined by IRMS. Plasma water was used as the precursor pool from which 2H2O is acquired for fatty acid synthesis. In brief, plasma water was isolated from whole plasma using centrifugal filtration (Millipore Ultrafree-0.5®, Fisher Scientific, Ottawa, ON, Canada) at 12,000 g for 45 min. Plasma water was collected, and 2H/1H enrichment was determined using a high temperature conversion/elemental analyzer coupled to a Delta V Plus IRMS instrument. Methyl-palmitate and plasma water samples were analyzed in conjunction with a working reference gas consisting of a known 2H/1H enrichment. Samples were normalized against the working reference gas and reported as δ(‰)16:0 and δ(‰)H2O, respectively. All IRMS data were analyzed using Isodat software (Thermo Fisher Scientific Inc.). FSRs were calculated using the following formula: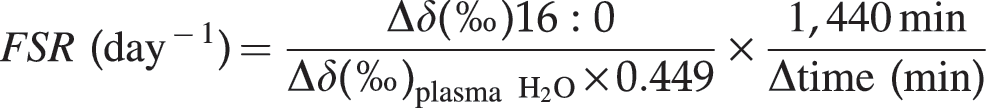
where Δδ(‰)16:0 and Δδ(‰)plasma H2O is the difference in palmitate and plasma water 2H enrichment between day 28 and 29, respectively. 17 To standardize all data over a 24-hour period, FSR data were multiplied by 1,440 min/Δtime (min), where 1,440 min is the number of minutes in 24 hours and Δtime (in min) is the difference in time between deuterium ingestion and the day 29 blood sample. Finally, 0.449 represents the correction factor for deuterium incorporation into endogenous palmitic acid and is based on previous research demonstrating that a maximum of 0.87 g of 3 H g-atom/g-atom of C is incorporated into newly synthesized fatty acids during adipose tissue lipogenesis. 18
Net synthesis of VLDL-palmitic acid (in g/day) were calculated using previously described methods, using the assumption that 300 mg of VLDL-TG fatty acids/kg of body weight is synthesized daily. 16
Statistical analysis
All statistical analyses were completed with SPSS version 11.5 (SPSS Inc., Chicago, IL, USA) using linear mixed model analysis of covariance with subject as a random factor. Repeated measures were used to determine the existence of effects of time and a time×treatment interaction. For thermogenic data and cumulative oxidation, total AUC and incremental AUC at each time point were compared between treatment groups. When appropriate, other factors were tested as covariates, including body weight, body mass index, total lean mass, total fat mass, REE, and resting substrate oxidation. For thermogenic data, the interaction day×treatment was also tested as a covariate. In addition, the effect of treatment, gender, phase, and treatment sequence were included in the model as fixed factors when their effect on the independent variable was significant. Comparisons between treatments were assessed using Bonferroni's post hoc test. All data are presented as mean±SEM values except for baseline criteria, which are presented as mean±SD values. A level of significance at P<.05 was used for all analysis.
Results
Subject dropout and compliance
Altogether, 29 subjects were recruited for the study. Four subjects dropped out for personal reasons, and two subjects left the study for medical reasons. Of the 23 volunteers who completed the entire 12-week trial, seven were men, and 16 were women. Statistical analysis for all measurements revealed no effect of gender, age, or an age×gender interaction.
Postprandial energy expenditure, TEF, and substrate utilization
Results for postprandial energy expenditure are summarized in Figure 1. Treatment had no effect on REE during week 1 (WF, 0.90±0.03 kcal/min; FPF, 0.90±0.03 kcal/min; WPF, 0.84±0.03 kcal/min) or week 4 (WF, 0.89±0.03 kcal/min; FPF, 0.85±0.03 kcal/min; WPF, 0.93±0.03 kcal/min). For TEE, no difference between treatments was observed during week 1 (Fig. 1A). After 4 weeks, TEE was 5% lower (P=.007) in the FPF group (333.0±3.6 kcal/330 min) compared with WF (349.3±3.6 kcal/330 min) (Fig. 1B). Analysis of incremental AUC indicate that postprandial energy expenditure was reduced shortly following the breakfast meal. At 30 min after breakfast, FPF (29.0±0.2 kcal/30 min) and WPF (29.1±0.3 kcal/30 min) demonstrated lower (FPF, P=.004; WPF, P=.025) incremental energy expenditure compared with WF (29.8±0.3 kcal/30 min). After 90 min, FPF (64.9±0.8 kcal/60 min) tended (P=.054) to maintain a reduced level of postprandial energy expenditure compared with WF (67.5±0.9 kcal/60 min). Energy expenditure was positively correlated with calories consumed during the breakfast meal (r=0.869, P<.001).

Effect of WF, FPF, and WPF on total energy expenditure during (
No differences in CHO oxidation were observed across treatments during week 1 (Fig. 2A). After 4 weeks, CHO oxidation tended to be lower (P=.075) in the FPF group (44.7±2.1 g/330 min) compared with the WF group (51.2±0.1.9 g/330 min) (Fig. 2B). Similar to TEE, week 4 CHO oxidation was positively correlated with calories during breakfast (r=0.732, P<.001). No differences in total fat oxidation were observed across treatment groups during week 1 or week 4 (Fig. 3).
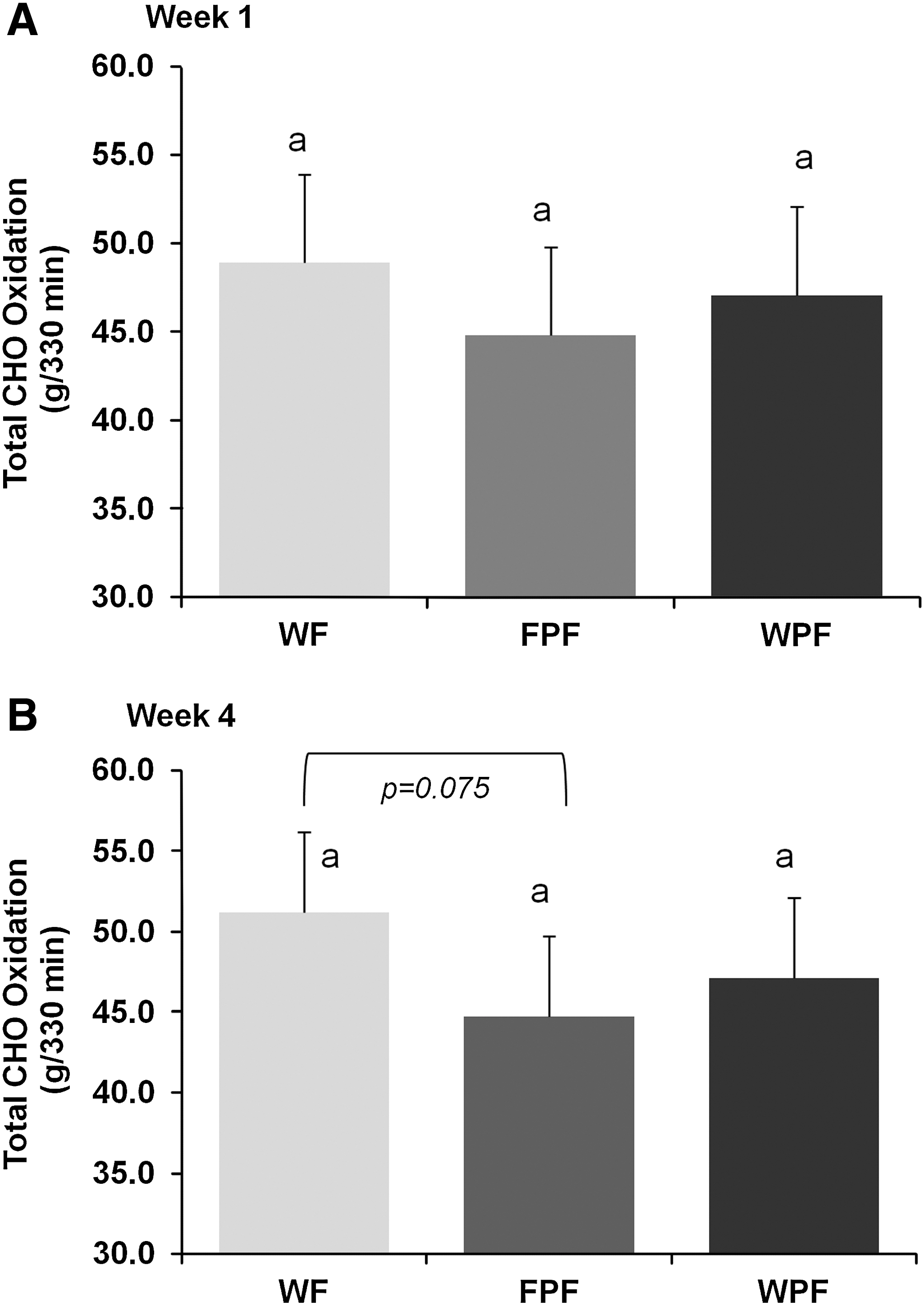
Effect of WF, FPF, and WPF on total CHO oxidation during (
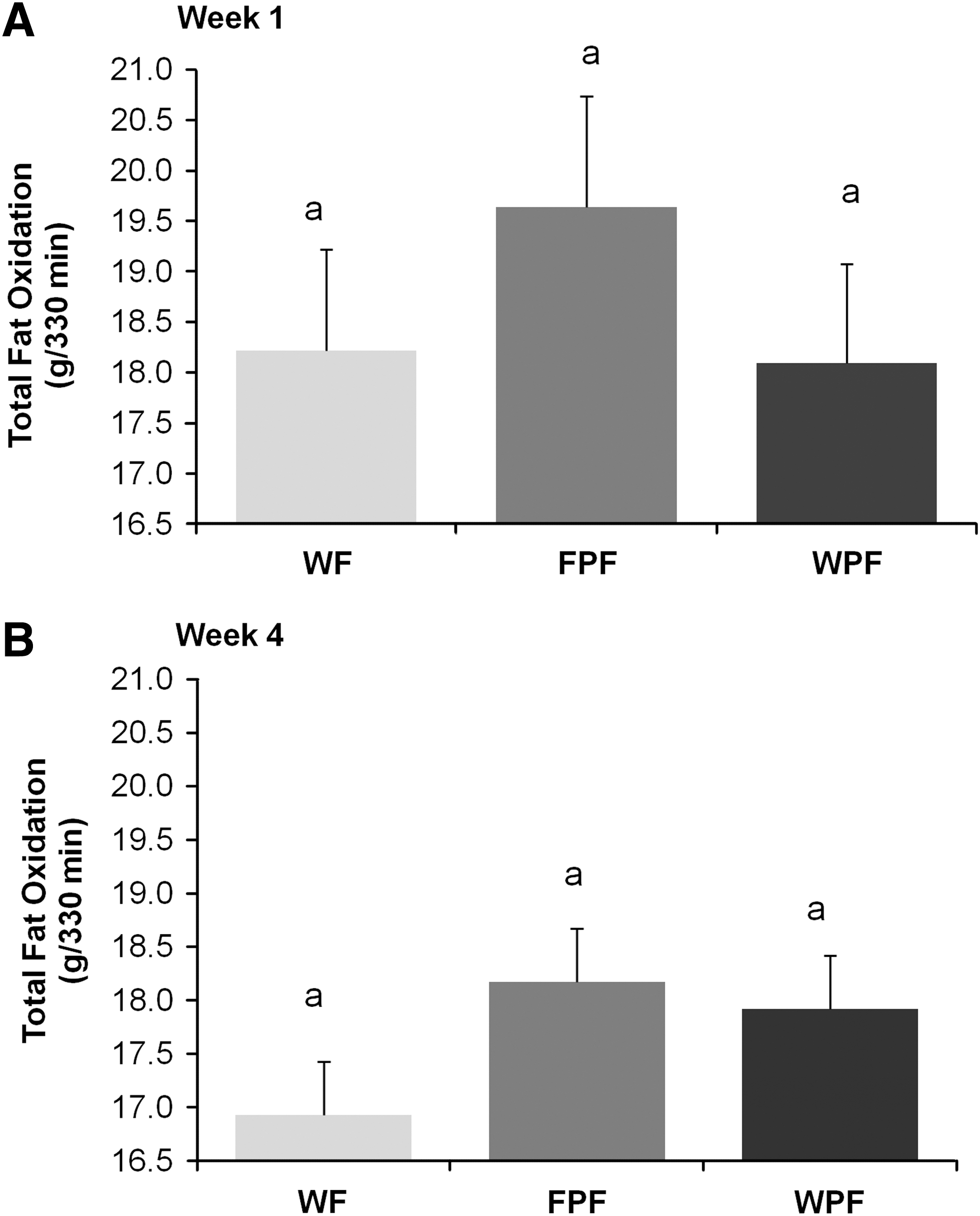
Effect of WF, FPF, and WPF on total fat oxidation during (
Neither treatment demonstrated any effect on TEF during week 1. However, TEF tended to be lower (P=.092) during week 4 in the FPF group (51.0±3.5 kcal/330 min) compared with the WF group (62.3±3.2 kcal/330 min) (Fig. 4A). When TEF data were normalized to energy consumed during the breakfast meal, a lower (P=.018) proportion of calories was expended during diet-induced thermogenesis with FPF (4.6±0.3%) compared with WF (5.7±0.3%) (Fig. 4B). The proportion of calories expended during diet-induced thermogenesis with WPF (4.9±0.3%) did not differ (P > .05) from other treatments.
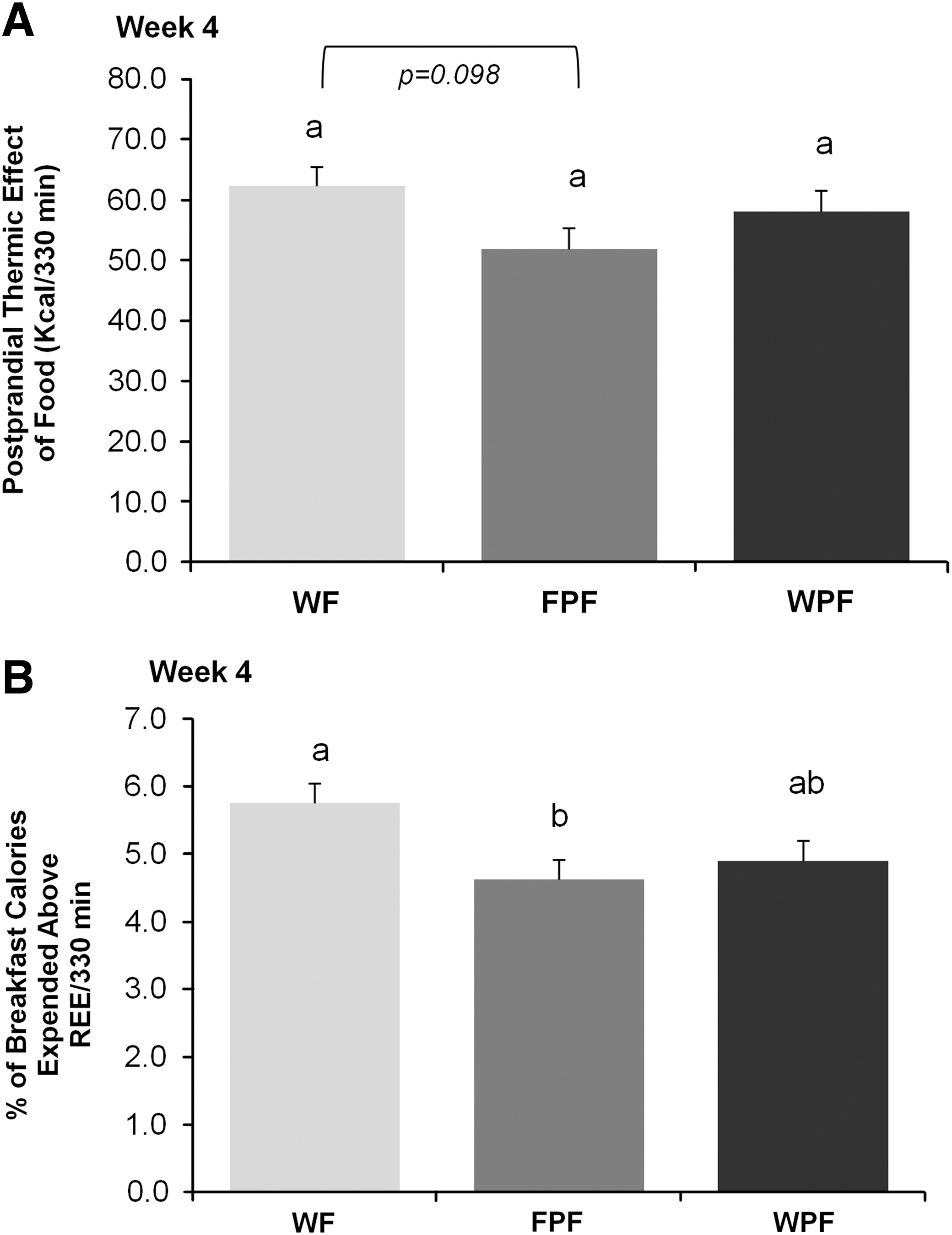
Effect of WF, FPF, and WPF on postprandial thermic effect of food (
Dietary fat oxidation
Cumulative oxidation of [1-13C]palmitic acid is presented in Figure 5A. Cumulative oxidation of dietary [1-13C]palmitic acid tended to be lower (P=.069) in the FPF group (3.0±0.2%) compared with the WF group (3.4±0.2%). Analysis of incremental AUC data demonstrated that 370 minutes after consumption of [1-13C]palmitic acid, a divergence in dietary fat oxidation was observed, with percentage of 13CO2 recovered in the breath being higher (P=.045) in the WPF group (0.96±0.05%) compared with the FPF group (0.81±0.05%) (Fig. 5A).

Effect of WF, FPF and WPF on the recovery of 13CO2 recovered in breath secondary to oxidation of dietary [13C]palmitic acid: (
Hepatic fractional TG synthesis
Deuterium incorporation into endogenous VLDL-derived palmitic acid was used as an index of TG synthesis. No differences in FSR for palmitic acid were found among WPF, FPF, and WF (Table 4). Calculation of net TG synthesis indicated that the amount of VLDL-derived palmitic acid produced was also similar among treatments (Table 4).
Data are mean±SEM values (n=23).
FSR, fractional synthesis rate; VLDL, very low-density lipoprotein.
Discussion
The main finding of the present study was that regular consumption of WF increased postprandial TEE, TEF, and CHO oxidation, compared with FPF. In addition, the effects of FPF and WF on TEE, postprandial thermogenesis, and CHO oxidation were not demonstrated during week 1, indicating that longer-term intake of WF is needed to elicit the observed changes in thermogenesis in the fed state.
WPF is a source of soluble and insoluble fiber (Table 2). Although few studies have examined the effects of fiber on postprandial thermogenesis, similar results were observed by Scalfi et al. 4 whereby high fiber meals reduced postprandial thermogenesis compared with a low fiber meal. The effect of fiber on postprandial thermogenesis was attributed to the concomitant reduction in postprandial glucose and insulin secretion. 4 Delayed glucose absorption, alongside a decrease in insulin release, would delay the exposure of glycolic pathways to glucose, thus reducing CHO oxidation and overall energy expenditure. In the present study, glucose levels were measured after each time increment, and no differences in glucose response between treatment groups were observed (data not shown). Nonetheless, the increase in energy expenditure shown in the present study with WF alongside the decrease in energy expenditure with FPF is unlikely to be secondary to postprandial glucose/insulin responses because the effects of WF and FPF on energy expenditure only occurred after 4 weeks of treatment (Fig. 1). Alternatively, these energetic responses may stem from mechanisms secondary to chronic ingestion of WF and FPF treatments. Although small, the ability for FPF to decrease energy expenditure is noteworthy given that the primary component of FPF treatments was WF and that the daily surplus of available carbohydrate from WF was minor at approximately 8 g/day above that of FPF treatments. Hence, future studies that utilize dietary interventions with higher doses of FPF may produce more pronounced hypothermogenic responses compared with conventional North American diets composed of high levels of WF. In addition, compared with week 1, WPF, which contains the same amount of pea-derived fiber as FPF treatments, had decreased energy expenditure from 356.9 kcal/330 minutes to 342.3 kcal/330 minutes during week 4, suggesting that the fiber component of WPF has a similar effect as observed with FPF. The need for chronic ingestion of high fiber meals to modulate energy expenditure is supported by Khossousi et al., 19 in whose study a single meal containing 15 g of fiber elicited no effect on energy expenditure after 6 hours compared with the same meal containing 3 g of fiber. Data from the present study indicate that chronic ingestion of low doses of FPF prevents increases in energy expenditure secondary to chronic intake of white flour.
The data demonstrate that the increase in energy expenditure observed with WF was due to an increase in CHO oxidation (Fig. 3). Again, it is difficult to delineate biological mechanisms by which FPF would inhibit an increase in CHO oxidation compared with WF, given the high levels of WF within the FPF treatments. Baseline data suggest that men in the study are somewhat insulin resistant with fasted glucose levels of 5.7 mmol/L (Table 1). Insulin resistance has been shown to reduce postprandial thermogenesis independent of obesity, 20 but previously published homeostasis modeling assessment data from the present study demonstrated no difference in insulin resistance after 4 weeks of receiving WPF and FPF. 21 Moreover, both WPF and FPF were shown to decrease insulin resistance 25% after 4 weeks of treatment compared with WF. 21 Given that insulin resistance was reduced, we hypothesized that postprandial thermogenesis would have increased in the FPF and WPF groups. Nonetheless, this was not observed, and we can only speculate as to why chronic intake of FPF prevented the observed increase in CHO oxidation compared with WF. Production of short-chain fatty acids—specifically, butyrate—as a result of microbial fermentation of fiber in the large intestine is a plausible mechanism to explain the FPF-related decrease in CHO oxidation. Butyrate has been shown to modulate hepatic CHO metabolism via increased postprandial hepatic glycogen synthesis 22 and reduced the rate of glycolysis in rat hepatocytes. 23 Enhanced glycogen synthesis would manifest itself as a reduction in CHO oxidation during indirect calorimetric measurements because glycogen synthesis pathways do not consume O2 or produce CO2. Nonetheless, pea fiber-induced butyrate production as a means of modulating CHO is only speculative and requires further evaluation.
Given that yellow peas are high protein foods and high protein diets are thermogenic, we had proposed that yellow peas and other pulse crops could be used as ingredients that modulate energy expenditure. However, results from the present study do not support this notion. Nonetheless, in order to fully dismiss yellow peas as non-thermogenic ingredients, future studies should evaluate and compare pulses and isolated pulse-derived proteins and bioactives without the addition of a background diet. Also, given the range of body mass indexes of those recruited for the present study, it cannot be overlooked that the doses of WPF used could have been insufficient to elicit a thermogenic response. Nonetheless, the observation that chronic intake of WPF yielded energy expenditure and CHO oxidation profiles that were comparable to those induced by WF could be important for overweight individuals implementing dietary lifestyle changes. Replacing WF with WPF would incorporate fiber, antioxidants, and protein not found in WF and thereby improve the overall quality of the diet.
Finally, neither treatment had any effect on hepatic TG FSR or net VLDL-palmitic acid synthesis. Moreover, results from net VLDL-palmitic synthesis correspond to previously published data. 16 The dietary factors most renowned for their effects on TG synthesis are marine-derived ω-3 fatty acids via the modulation of transcription factors such as peroxisome proliferator-activated receptor. 24 However, to our knowledge, no studies have concluded that pulse crops increase or decrease hepatic TG lipogenesis in humans. One would expect that because FPF modulated CHO oxidation, TG synthesis would have been affected. However, studies indicate that glycogen storage is favored over CHO-derived TG synthesis. Moreover, CHO-derived TG synthesis only occurs with excess CHO ingestion and overfeeding for ≥2–3 days. 25,26 In the present study, the amount of food each subject received was precisely controlled to prevent weight loss or weight gain. In addition, the present study used a crossover design; thus, if a subject was overfed and created metabolic conditions that favor TG synthesis, this effect would have been demonstrated within energetic measurements in subsequent phases. We conclude that the addition of ½ cup of yellow peas/day to the diet does not modulate hepatic TG synthesis.
Conclusions
In summary, chronic dietary incorporation of FPF at a dose equivalent to ½ cup of whole yellow peas/day produced lower TEE, TEF, and CHO oxidation compared with WF. Conversely, chronic intake of WPF was able to negate the effects of pea-derived fiber on postprandial energetics. Although data from the present study suggest that WPF could increase the oxidation of exogenous lipid, studies that investigate the affect of WPF on postprandial dietary fat oxidation beyond 6 hours are needed to confirm these results. The present study demonstrates that chronic ingestion of different fractions of yellow peas imposes distinctive effects on postprandial energy expenditure and substrate utilization and justifies further exploration of the effects of other pulse crops on postprandial energetics.
Footnotes
Acknowledgments
We would like to thank Pulse Canada for funding this study. We would also like to thank Kim Robinson, Xin Wang, Katherine Leung, Darren Speziale, and Yen Ming Chan and the kitchen staff for their assistance with the clinical trial, Gerber® for their generous donation of the baby food used for flavoring the treatment muffins, and Best Cooking Pulses Inc. for providing the whole and fractionated pea flours. Finally, we acknowledge Dr. Stephen Cunnane and Mélanie Fortier for their mass spectrometer expertise in breath 13CO2 analysis as well as Dr. Edward Kesselman for his medical expertise.
Author Disclosure Statement
No competing financial interests exist for C.P.F.M. and P.J.H.J.