
Abstract
Resveratrol inhibits endothelin-1, a vascular tension regulator. We synthesized the resveratrol analogue 4,4′-dihydroxy-trans-stilbene with 2 hydroxyl groups in the 4 and 4′ position to obtain a molecule more active than resveratrol (3,4′,5-trihydroxy-trans-stilbene). The results demonstrate that 4,4′-dihydroxy-trans-stilbene led to a significant decrease in total endothelin-1 secretion and in endothelin-1 messenger RNA (mRNA) levels in human endothelial cells. In addition, resveratrol and its analogue decreased endothelin-converting enzyme-1 mRNA levels and further reduced the activity of the enzyme. 4,4′-dihydroxy-trans-stilbene was more active than resveratrol because the new molecule exerted greater activity at the level of endothelin synthesis and conversion, even at a lower concentration. Although 4,4′-dihydroxy-trans-stilbene and resveratrol inhibited formation of reactive oxygen species and lipid peroxidation, the treatment of cells with different oxidant agents did not modify the endothelin-1 release. This finding suggests that the inhibition of endothelin-1 secretion is independent of the antioxidant properties of the 2 compounds. On the basis of these results, the resveratrol analogue 4,4′-dihydroxy-trans-stilbene could be a promising chemopreventive agent against cardiovascular diseases.
Introduction
A
Corder et al. have shown that red wines and cranberry juice strongly inhibit the synthesis of endothelin-1 (ET-1), a 21–amino acid peptide with vasoactive properties. 3 The mechanism of inhibition was proposed, taking into account the formation of metabolites of red wine and cranberry juice constituents with vasoactive activity at nanomolar concentrations. ET-1 was originally described as a highly potent vasoconstrictor produced by endothelial cells in both cell cultures and in vivo. 4 The main step in the ET-1 pathway is the conversion of the inactive precursor “big-endothelin” to the biologically active peptide in endothelial cells, which is catalyzed predominantly by endothelin converting enzyme-1 (ECE-1). 5 –7 If endogenously generated, ET-1 appears to play a modest role in the healthy organism, whereas increased plasma levels of the vasoconstrictor are associated with some forms of hypertension or heart failure. This finding corroborates a major role of the peptide in the etiology of these condition. 8 Indeed, endothelin antagonists inhibit the development of early stages of atherosclerosis, such as endothelial dysfunction or fatty-streak formation, 7 and reduce myocardial infarction in established diseases. 9
Pharmacologic studies indicated an effect of RSV on vascular smooth muscle cells. Ruef et al. 10 showed that the addition of RSV resulted in the inhibition of H2O2–induced Pre-pro-ET-1 (PpET-1) expression through an antioxidant mechanism. Moreover, RSV potently inhibits stress-induced (including oxidative stress) ET-1 gene expression, partially by interfering with the ERK1/2 pathway through attenuation of the formation of reactive oxygen species (ROS). 11
Recent experimental evidence suggests that a strong correlation between antioxidant and vasoprotective activity may not be so straightforward. 12 We have reported that the number and the position of the hydroxylic groups play a crucial role in some biological activities of RSV and its stilbenic derivatives, such as antioxidant activity (in vitro and in vivo) and antiproliferative effect in normal and cancer cell lines. 2
To enhance the biological activity of natural stilbenic phytochemicals, 4,4′-dihydroxy-trans-stilbene (DHS), a resveratrol analogue with 2 hydroxyl groups in the 4 and 4′ positions, was synthetized. 13 This molecule has powerful antioxidant activity, 14 and we have shown that this molecule inhibits cell proliferation with higher efficiency but via a different mechanism from resveratrol. 15
However, the vasoactive effect, a property of resveratrol, has not yet been examined with DHS. In this study, we therefore investigated whether DHS modulates ET-1 secretion as occurs with RSV. In particular, we first assessed secretion and subsequently gene expression of ET-1 after treatment with DHS on human umbilical vein endothelial cells (HUVEC), comparing these activities with those of RSV. To fully elucidate the mechanism of action on the ET-1 system, we investigated the effect of DHS and RSV on ECE-1 expression and activity. In parallel, the antioxidant activity of DHS and RSV was assessed in HUVEC cells.
Materials and Methods
Synthesis of 4,4′-dihydroxy-trans-stilbene
4,4′-dihydroxy-trans-stilbene was obtained by Perkin condensation of 4-hydroxybenzaldehyde and acetic anhydride in the presence of 4-hydroxyphenylacetic acid and triethylamine, as described elsewhere. 13
Reagents
Trans-resveratrol (99% purity), N-(α-rhamnopyranosyloxy-hydroxyphosphinyl)-Leu-Trp disodium salt (phosphoramidon disodium salt; PH) (97% purity) and all other chemicals of reagent grade were obtained from Sigma (St. Louis, MO, USA), unless otherwise specified.
Cell culture and treatments
HUVEC cells, provided by Professor J.A.M. Maier (University of Milan, Italy), were cultured in 199 medium supplemented with 10% fetal bovine serum (both from Invitrogen, Carlsbad, CA, USA), 5 μg/mL endothelial cell growth factor (Roche, Basel, Switzerland), and 5 U/mL heparin on gelatin-coated dishes in a 5% CO2 atmosphere. HUVECs (70% confluent) seeded in 6-well dishes were growth-arrested in serum-free medium for 24 hours. Cell treatments with RSV or DHS were performed by incubating cells for 24 hours with the substances dissolved in dimethyl sulfoxide, with the solvent never exceeding 0.1% in the culture medium. The final RSV and DHS concentrations ranged from 1 to 90 μM. Cell treatments with oxidizing agents were conducted by incubating cells with 10 μM hemin, 200 μM/1 mM Fe2+/ascorbate, and 10 μM CdCl2 dissolved in O.1 M NaOH (hemin) or ultrapure H2O (Fe2+/ascorbate and CdCl2) for 6 hours.
Cytotoxicity assays
Cell toxicity was determined by using the 3-[4,5-di-methylthiazol-2-yl]-2,5-diphenyl-tetrazoliumbromide (MTT) colorimetric assay. 2 The maximum nontoxic concentrations of 30 μM for RSV and 7.5 μM for DHS were used for all subsequent experiments.
Measurement of ET-1 concentration
After treatment with RSV or DHS, the ET-1 concentration in culture medium was determined, as reported elsewhere. 16
Reverse transcriptase polymerase chain reaction expression of Pp-ET-1, ECE-1, and β-actin genes
Reverse transcriptase polymerase chain reaction expressions of ET-1 and β-actin were determined, as reported elsewhere.
16
Primers for the ECE-1 gene were as follows: 5′-TCTGTGCTTCTGGGAAGTGTGAGC-3′ (sense primer) and 5′-GCTCACACTTCCCAGAAGCACAGA-3′ (anti-sense primer). Gel images of the bands were acquired by a Canon Canoscan 9950F transparencies scanner (Canon, Tokyo, Japan), and the densitometric analysis was performed on a Macintosh G3 computer using the public domain NIH Image program (developed at the U.S. National Institutes of Health and available at
Inhibition assay of recombinant ECE-1 activity
Recombinant human ECE-1 was purchased from R&D Systems (Minneapolis, MN, USA); the protein was reconstituted in 25 mM Tris and 0.15 M NaCl (pH, 8). The assays were performed at 37°C with samples containing 0.1 M 2-(N-morpholino) ethanesulfonic acid; 0.1 M NaCl (pH, 6); 5 ng enzyme; and concentrations of RSV, DHS, or PH ranging from 0.1 to 100 μM. Reactions were initiated by the addition of 100 ng of big bovine ET-1. After a 30-minute incubation, the production of ET-1 in each sample was measured by the Imubind-ET-1 enzyme-linked immunosorbent assay kit (Biomedica, Vienna, Austria). Data were plotted as percentage activity relative to uninhibited control reactions versus inhibitor concentration. The 50% inhibiting dose (ID50) values were calculated by Dixon plots of the experimental data, as reported elsewhere. 2
Oxidant conditions and antioxidant activity
Oxidant conditions were induced by treating HUVECs with 10 μM hemin or 200 μM/1 mM Fe2+/ascorbate or 10 μM CdCl2. Oxidative damage was assessed by determining lipid peroxidation aldehydic end products, namely hexanal, by high-performance liquid chromatography. 17 Detection of intracellular ROS was recorded by monitoring changes in 2′,7′-dichlorodihydrofluorescein diacetate (Invitrogen) fluorescence with flow cytometry in a Coulter Epics XL cytometer (BD Biosciences, San Jose, CA, USA). 17
Antioxidant activity was assessed by measuring lipid peroxidation and ROS production induced by hemin (the most effective oxidant in our experiments) in endothelial cells treated with 7.5 μM DHS or 30 μM RSV for 24 hours, as described elsewhere. 16
After the induction of oxidative stress by hemin, Fe2+/ascorbate, or CdCl2, ET-1 release was also measured.
Statistical analysis
Data are presented as means±standard deviation. Statistical analysis was performed by using the Student t-test. MS Excel™ (Microsoft, Redmond, WA, USA) was used for statistical analysis; significance cutoffs were 95% (P<.05) and 99% (P<.01).
Results
Figure 1 shows the chemical structure of RSV and DHS.

Chemical structures of trans resveratrol (RSV) and 4,4′-dihydroxy-trans-stilbene (DHS).
Preliminary experiments were performed to study the cytotoxic effect of the 2 compounds by the MTT test (Fig. 2) after 24 hours of treatment with each compound. RSV did not significantly affect cell growth or survival at up to 30 μM, whereas a statistically significant (P<.01) overall 40% reduction of living cells compared with control (untreated) cells was observed at the highest concentration used (90 μM). When compared with RSV, DHS appeared to be more potent. In fact, it induced a cytotoxic effect by 32% (P<.05 compared with control cells) at 15 μM and by 60% (P<.01) at 90 μM. For this reason, all subsequent experiments were performed by treating HUVECs with 30 μM RSV or 7.5 μM DHS (i.e., at their respective maximum nontoxic concentrations).
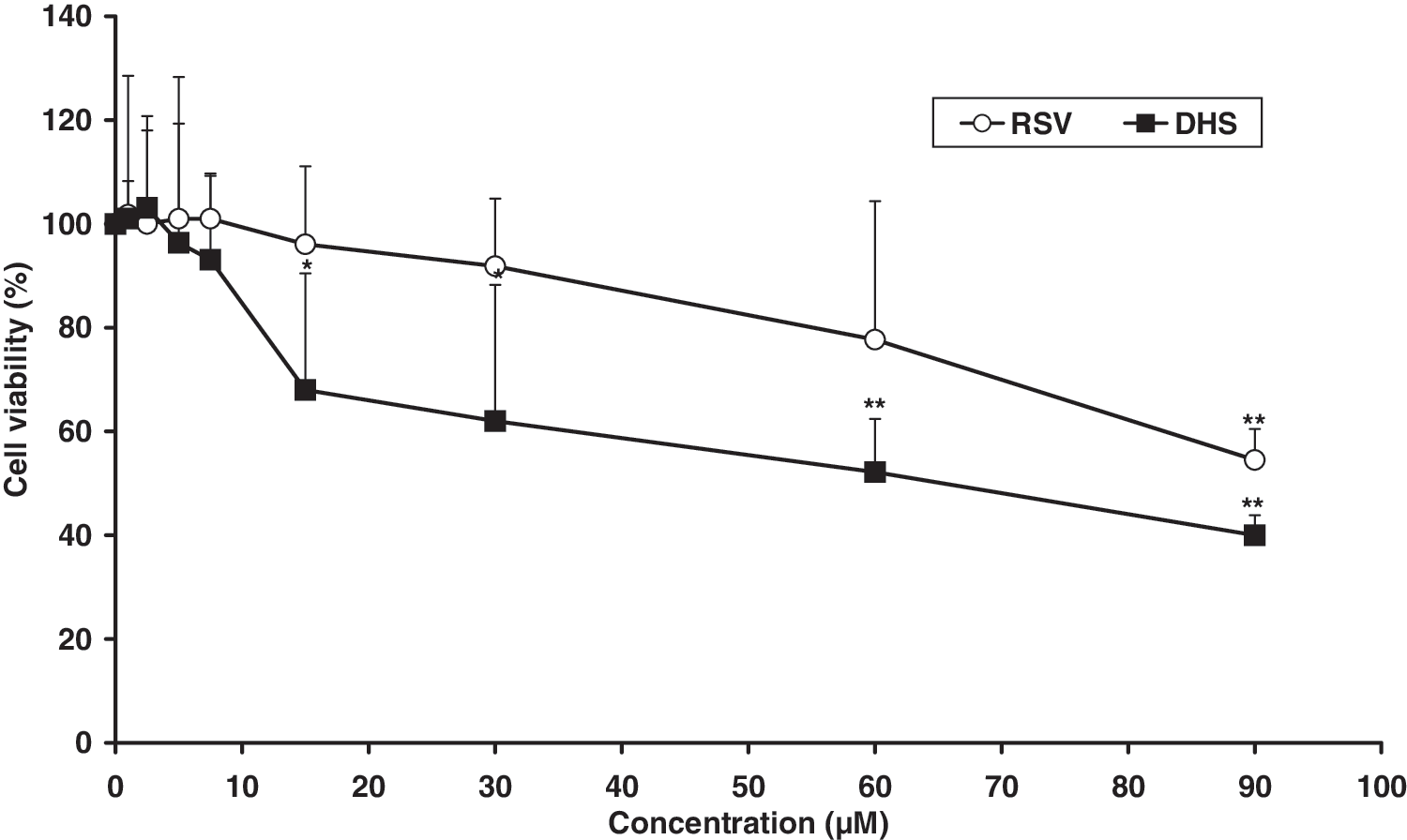
Effects of 24 hours of incubation with different concentrations of trans resveratrol or 4,4′-dihydroxy-trans-stilbene on cell viability. The cytotoxicity was determined by 3-[4,5-di-methylthiazol-2-yl]-2,5-diphenyl-tetrazoliumbromide assay. The data are expressed as mean±standard deviation percentages for at least 3 independent experiments and are referred to untreated control cells. The mean of at least 3 experiments is shown. *P<.05, **P<.01 versus control (untreated) cells.
Effects of RSV or DHS on ET-1 release
Culture media containing ET-1 from untreated cells or cells treated with the 2 compounds were collected and analyzed by enzyme immunoassay. As reported in Figure 3, the stilbenic molecules significantly (P<.01) inhibited ET-1 release from 3.89 fmol/mL per 1×106 in untreated cells to 2.01 and 2.09 fmol/mL per 1×106 in RSV- and DHS-treated cells after 24 hours of treatment, with a reduction of 48.29% and 46.12%, respectively.
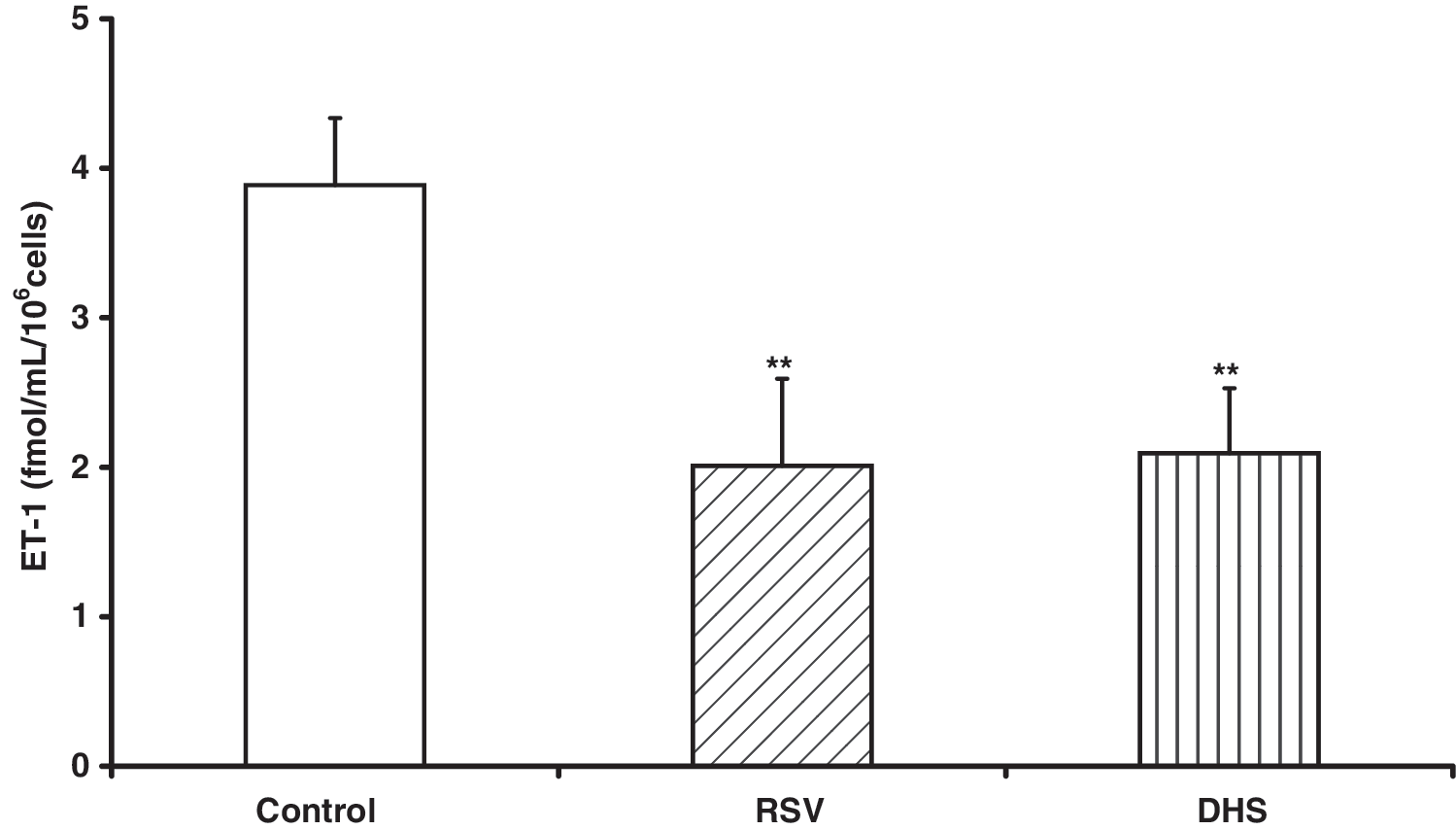
Inhibition of endothelin-1 release by RSV or DHS in human umbilical vein endothelial cells after 24 hours of incubation. The analysis was performed by using a quantitative immunoassay as reported in the Materials and Methods section. Cells were treated with RSV or DHS at the concentrations of 30 μM and 7.5 μM, respectively. The data are expressed as mean±standard deviation. The mean of 5 experiments is shown. **P<.01 versus control cells. ET-1, endothelin-1.
Effects of RSV or DHS on PpET-1 mRNA levels
The sequence analysis of the human ET-1 complementary DNA revealed a single-copy gene that encodes for a precursor, PpET-1. This molecule is proteolytically cleaved by proteases (furin-like enzymes) to produce the big-ET-1; big-ET-1 is subsequently cleaved into the active molecule ET-1. To investigate the possible effect of 2 compounds at the molecular transcriptional level, cells were treated for 24 hours and then collected and analyzed by using semi-quantitative polymerase chain reaction. The results showed that the effect of the treatment with RSV or DHS occurs at the first step (i.e., at PpET-1 transcription) (Fig. 4). The densitometric analysis showed that the exposure of endothelial cells to DHS, but not to RSV, decreases the amount of PpET-1 mRNA (P<.01).
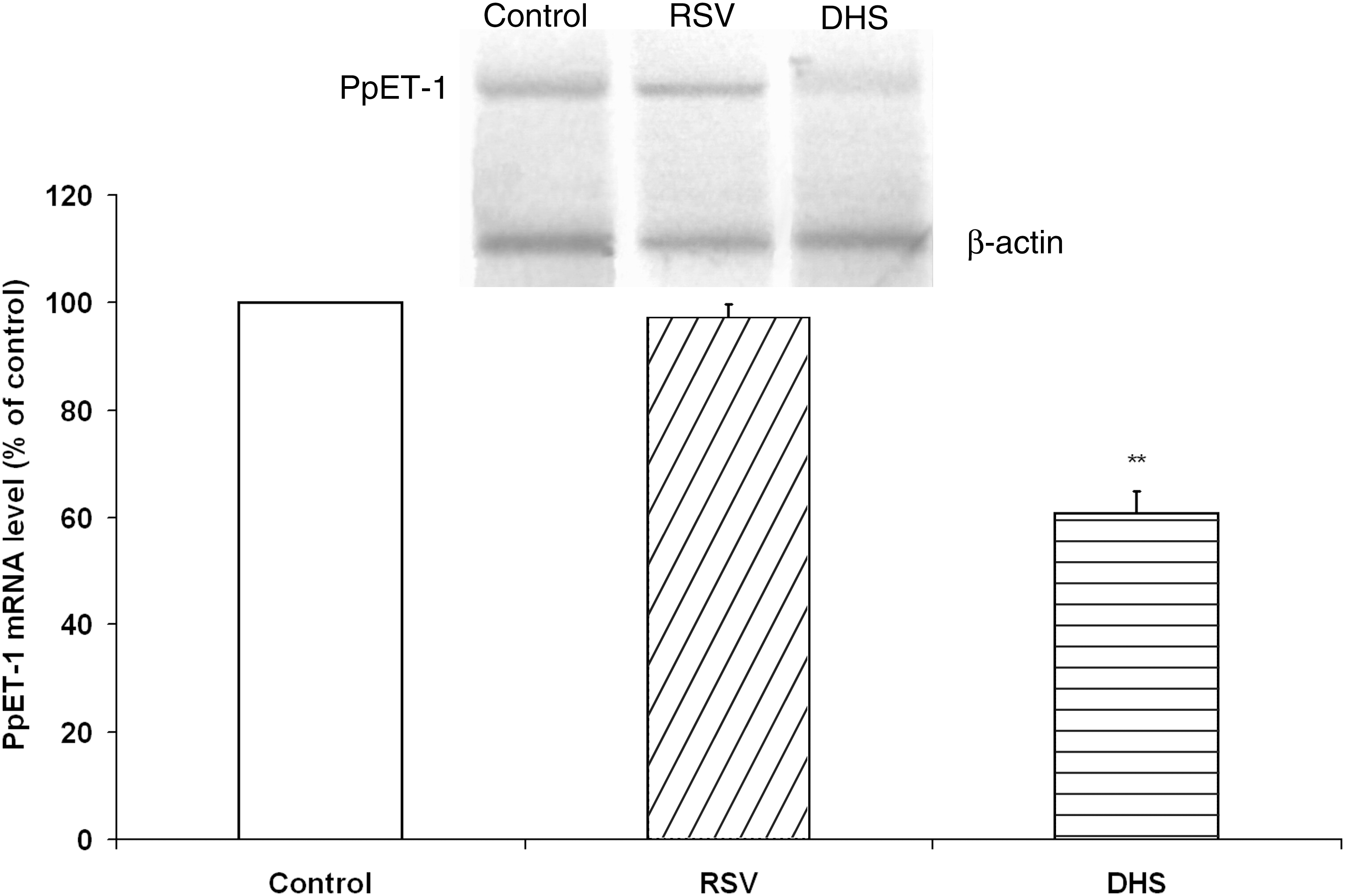
Effects of RSV or DHS on PpET-1 mRNA levels. The blot shown is representative of 3 independent experiments with similar results. β-actin RNA was used to normalize the RNA applied in each lane. The histogram shows the densitometric analysis of 3 different blots. The results (means±standard deviation) are expressed as a percentage of basal expression from 3 independent experiments. **P<.01 versus control cells. mRNA, messenger RNA; PpET-1, Pre-pro-endothelin-1.
Effects of RSV or DHS on ECE-1 mRNA levels and on ECE-1 activity
To investigate the different mechanisms by which the above stilbenes inhibit ET-1 release, we examined the effect of both compounds on ECE-1 gene transcription and enzymatic activity. As shown in Figure 5, the treatment of endothelial cells with RSV or DHS for 24 hours decreases the amount of ECE-1 mRNA (P<.01). This reduction at the mRNA level was similar to that observed when HUVECs were treated with PH (P<.01), the selective ECE-1 inhibitor. Furthermore, DHS was more effective than RSV (P<.05) in decreasing ECE-1 mRNA levels, even at a lower concentration.

Inhibition of ECE-1 mRNA levels by RSV, DHS, or phosphoramidon disodium salt on human umbilical vein endothelial cells. The blot shown is 1 representative out of 5 independent experiments with similar results. β-actin RNA was used to normalize the RNA applied in each lane. The histogram shows the densitometric analysis of the 5 different blots. The results (means±standard deviation) are expressed as a percentage of basal expression from 5 independent experiments. The results of 5 experiments are shown. **P<.01 versus control, • P<.05 versus RSV. ECE-1, endothelin-converting enzyme-1; PH, phosphoramidon disodium.
The results obtained in the in vitro assay on recombinant ECE-1 activity (Fig. 6) showed that DHS strongly inhibits ECE-1 activity, whereas RSV is inactive. The corresponding ID50 values for the compounds are listed within Fig. 6: the stronger the inhibitor, the smaller the ID50 value.
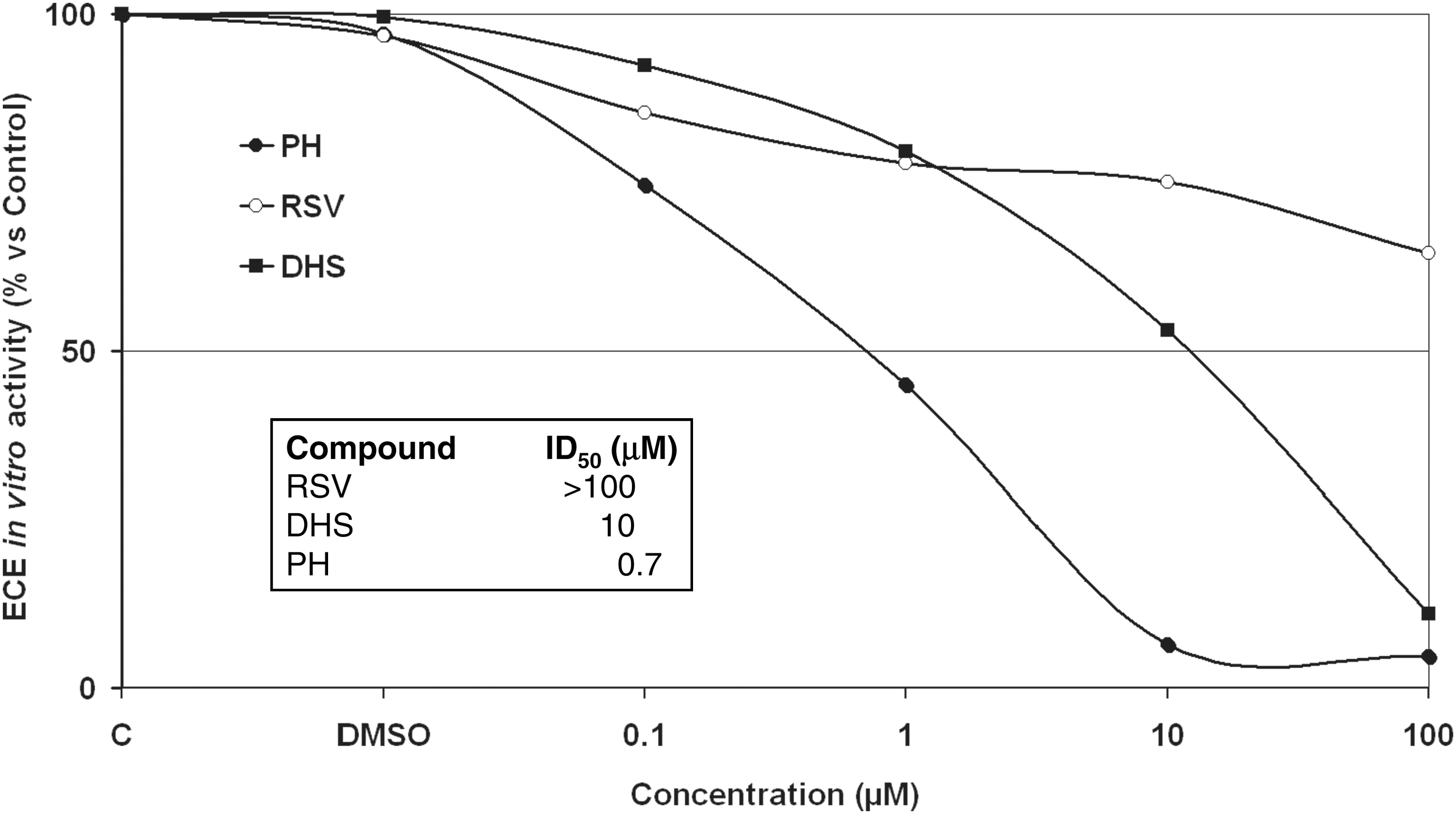
Effects of RSV, DHS, or phosphoramidon disodium salt on ECE-1 activity measured in vitro by using recombinant protein in the absence (control) and presence of compounds. The assay was performed, as described in the Materials and Methods section, in the presence of different concentrations of each substance (ranging from 0.1 to 100 μM). DMSO, dimethyl sulfoxide; ID50, 50% inhibiting dose.
Oxidant conditions and antioxidant activity
To ascertain whether the RSV and DHS effect in inhibiting ET-1 production is exclusively dependent on oxidative stress counteraction, HUVEC cells were previously treated with 10 μM hemin or 200 μM/1 mM Fe2+/ascorbate or 10 μM CdCl2. Hemin was the most effective stressor in terms of lipid peroxidation and ROS production and was chosen thereafter (Fig. 7). RSV and DHS were effective as antioxidants by lowering lipid peroxidation and ROS production induced by hemin (Fig. 8). Nevertheless, the results showed that no statistically significant variation in ET-1 release from HUVEC cells occurred by treating cells with the all the above oxidative stressors (Fig. 9).
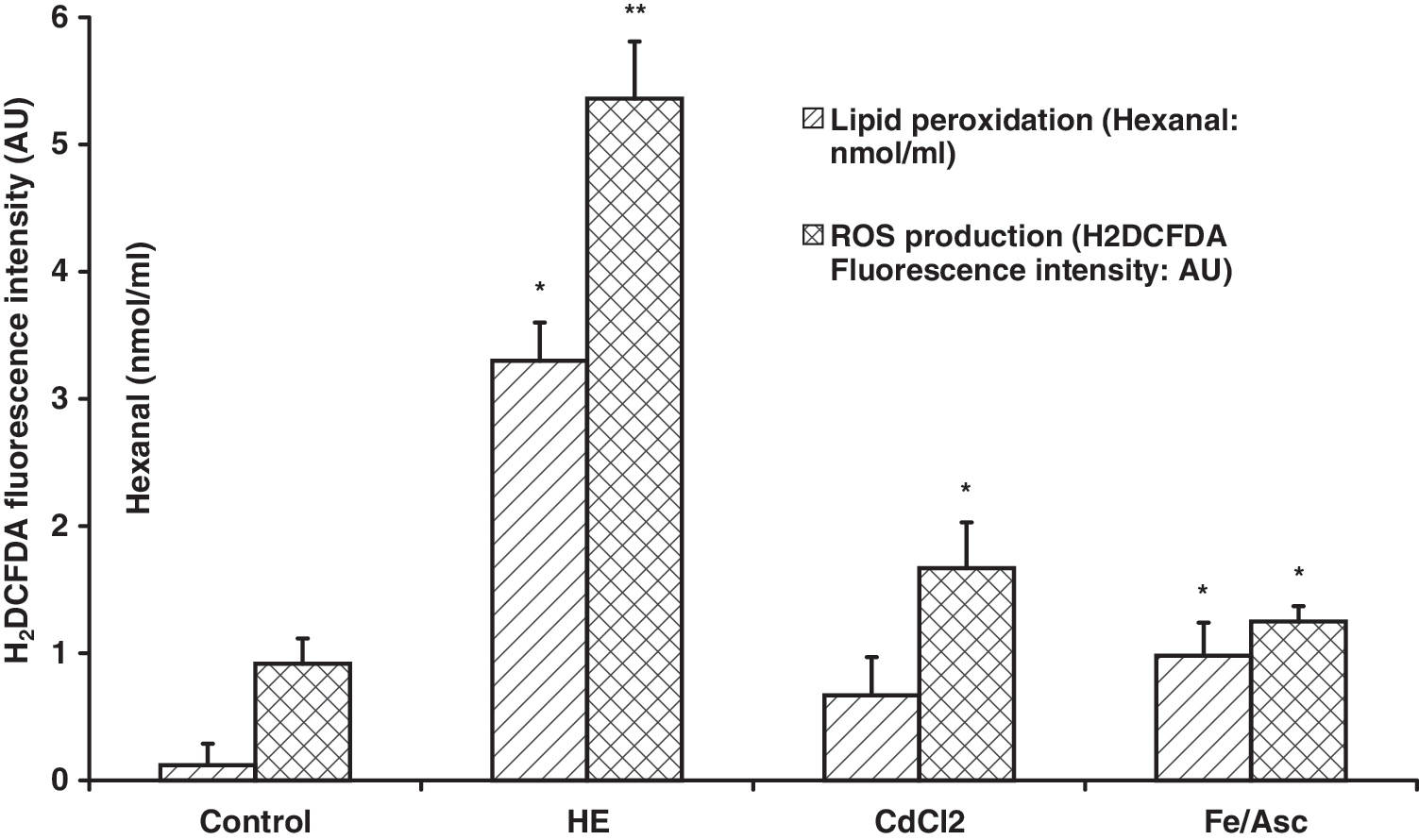
Oxidant conditions induced by treatment with hemin, CdCl2, or Fe/Asc. Lipid peroxidation was assessed by hexanal production; intracellular levels of reactive oxygen species were revealed by fluorescence intensity by H2DCFDA by flow cytometry in human umbilical vein endothelial cells. The data are expressed as mean±standard deviation. The results of 5 experiments are shown. *P<.05 versus control cells; **P<.01 versus control cells. AU, arbitrary units; H2DCFDA, 2′,7′-dichlorodihydrofluorescein diacetate; HE, hemin; ROS, reactive oxygen species.

Effects of preincubation with 30 μM RSV or 7.5 μM DHS on lipid peroxidation and production of reactive oxygen species induced by hemin. The data are expressed as mean±standard deviation. The results are mean values of at least 3 independently-reproduced experiments; *P<.05 versus hemin-induced oxidative stress; **P<.01 versus hemin-induced oxidative stress.
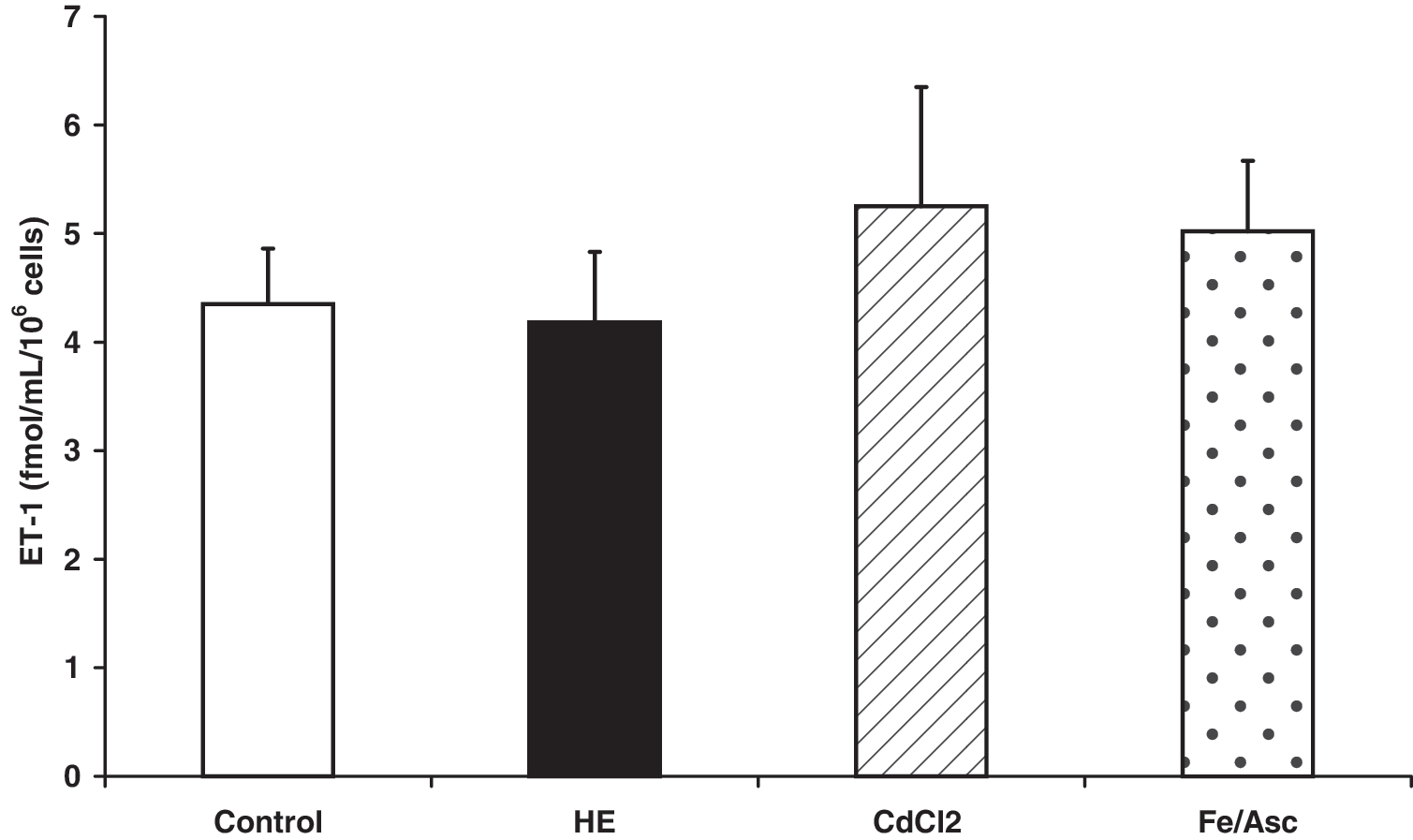
Effects of different oxidative stressors (hemin, CdCl2, and Fe/Asc) on ET-1 release. The mean of 5 experiments±standard deviation is shown.
Discussion
Epidemiologic evidence has suggested that a moderate consumption of red wine may be cardioprotective, although the precise mechanism responsible remains poorly understood. A possible explanation could be developed by taking into account the fact that RSV strongly decreases ET-1 synthesis in cultured aortic endothelial cells. 11 Such an effect was previously observed for anthocyanidins contained in red berries as well as in red wine. 16
Oxidant conditions induced by ROS can significantly alter vascular function; in particular, elevated ROS levels are involved in the release of ET-1. Some authors showed that the ROS may act as second messengers in cells exposed to various stimuli, 18,19 such as hemodynamic forces (including shear flow and cyclic strain) in order to stimulate ET-1 gene expression. 20
The vascular protective effect by phenolic compounds could therefore be attributed to their antioxidant properties. However, in the past few years, the fate and mode of action of phenolics, including RSV and its analogues and derivatives, became more complex than originally expected. 21 In fact, the protective effect of polyphenol components in red wine on vascular endothelial function has recently been demonstrated to be independent of antioxidant activity. 21
Several RSV analogues have been synthesized to improve the biological activity of the molecule. 15,22,23 In our previous studies, different trans RSV derivatives revealed a structure–activity relationship. In particular, we have demonstrated that the hydroxyl group in the 4′ position is required for the antioxidant activity; in contrast, the presence of 4′-OH together with stereoisometry in the trans conformation (4′-hydroxystyryl moiety) was required for the inhibition of cell proliferation. 2 On the basis of these earlier results, we have synthesized and reported the biological effects of DHS, an RSV analogue with more selective antioxidant and antiproliferative activities. 15
In this study, we investigated and compared the biological activity of RSV and DHS toward the ET axis, taking into account different mechanisms of action, including antioxidant activity. Preliminary toxicity tests indicated that DHS is more cytotoxic than RSV in HUVEC cells. For this reason, the molecules were used in subsequent experiments at their maximum nontoxic concentrations.
DHS exerted a significant decrease in ET-1 release and in PpET-1 mRNA levels, and RSV showed a significant decrease only at the protein secretion level (i.e., only at the last stage of the ET-1 pathway), with no effect at the transcriptional level. Although inhibition of ET-1 release by DHS was not significantly different from that of RSV, even if it also decreased PpET-1 mRNA levels, DHS was used at a 4-fold lower concentration, while RSV at the equimolar 7.5 μM concentration had no effect on ET-1 release (data not shown).
Synthesis and release of endothelin depend not only on ET-1 transcription and translation but also on 2-step conversion of its inactive form to the active form. 24 The key enzyme responsible for this conversion is ECE-1, a zinc-binding metalloendopeptidase located on the plasma membrane of endothelial cells. 25 ECE-1 mRNA levels were inhibited by both stilbenic compounds tested. Here again, DHS was significantly more active than RSV. As far as ECE-1 enzymatic activity is concerned, only DHS showed to be active as an inhibitor, whereas RSV was ineffective. These data denote a double mechanism of action: the first one at the transcriptional level and the second one involving activity of the enzyme. With respect to the reference ECE-1 inhibitor PH, 26 DHS showed a greater activity even at a 100-fold lower concentration. Therefore, in our experiments, DHS proved to be a more powerful ECE-1 inhibitor than the best-characterized ECE inhibitor available. 27
Although it is believed that ET-1 can increase oxidative stress, several observations suggest that ROS can increase ET-1 production in cultured endothelial cells and vascular smooth muscle cells. 10,28 –30 The relevance of these observations is unclear given that Saito et al. observed the contrasting finding that H2O2 actually decreases ET-1 mRNA and protein synthesis in endothelial cells. 31 In our experiments, no correlation between oxidative stress and ET-1 release was found. In fact, whereas RSV and DHS were active in protecting against hemin-induced lipid peroxidation and ROS production, oxidative stressors did not induce ET-1 release. These data support the hypothesis that oxidative stress plays a marginal role in the ET-1 axis.
In summary, the results presented here show the following: DHS inhibits the ET-1 release and lowers PpET-1 mRNA levels, whereas RSV lowers only ET-1 release in HUVECs (being ineffective at the transcriptional level); DHS is more active than RSV in diminishing ECE-1 mRNA levels and in inhibiting ECE-1 activity; and oxidative stress may play only a marginal role in the ET-1 axis.
Further studies on the structure–activity relationships of resveratrol analogues could lead to the development of a promising class of naturally derived compounds active as chemopreventive substances against cardiovascular diseases and cancer.
Footnotes
Acknowledgments
We thank Professor A. Rossi (Dip. Biochimica) and Dr. C. Scotti (Dip. Medicina sperimentale, Università di Pavia, Italy) for their help, Dr. G. Maga (IGM – CNR, Pavia, Italy) for conducting the ECE-1 activity calculations, and Professor E. Gherardi (MRC, Cambridge, United Kingdom) for his critical review of the manuscript.
Author Disclosure Statement
Authors declare that no conflict of interest exists.