
Abstract
This study examined the effects of oral administration of an enzymatic protein hydrolysate from green microalga Chlorella vulgaris (Cv-PH) on the nutritional recovery of malnourished Balb/c mice after a 3-day fasting period. Mice were refed with commercial diet supplemented or not supplemented with Cv-PH (500 mg/kg) for 8 days. Regardless of the diet used during refeeding, animal body weights and serum protein concentrations did not differ between groups. Mice given Cv-PH had a significant increase in hemoglobin concentrations. Most serum amino acid levels were similar in the control and Cv-PH animals. Starved mice refed with Cv-PH showed normal liver functions, as judged by liver weight, protein concentration, and the enzymatic activities of cholinesterase and arginase. Cv-PH increased DNA, protein content, and gut-mucosal weight. In addition, brush-border oligosaccharidase activities were also higher in the Cv-PH group. These findings suggest that Chlorella protein hydrolysate can be used to develop specific formulations suitable for pharmacologic nutrition.
Introduction
B
The unicellular green algae (Chlorophyta, Chlorophyceae) are suitable for protein health foods and food supplements. 4 –6 The enzymatic hydrolysis of cell proteins in green microalgae has been described as a promising method for improving the digestibility of algae protein, which makes the product usable in human nutrition. 7,8
Progress in hydrolysis techniques has led to the production of hydrolysates of many food proteins by using proteolytic enzymes. Because enzymatic protein hydrolysates appear to be more effective than intact protein or free amino acids, they have been widely used in specific formulations with clinical applications. 9,10 In addition, many food-derived peptides possess bioactive properties (e.g., antimicrobial, immunostimulatory, antihypertensive, mineral binding, and opioid) and are often multifunctional. 11 Because of their bioactive properties, some peptides have been claimed to be potential nutraceuticals for food and pharmaceutical applications. 12
We have previously reported on the use of Chlorella vulgaris cell biomass derived from outdoor cultivation in tropical conditions for the production of an enzymatic protein hydrolysate (Cv-PH). 13 Cv-PH enhances host defense activity in vivo by stimulating mechanisms involved in both innate and specific immune responses of malnourished mice. 14 This effect may be due to peptide sequences hidden in Chlorella protein, released by enzymatic hydrolysis. 15
Because most chronic disease states are associated with fasting, which contributes to the establishment of malnutrition, 16 we investigated the effect of an enzymatic protein hydrolysate from green microalga C. vulgaris on the nutritional recovery of malnourished mice. An extended knowledge of algal hydrolysate bioactivity would be useful in understanding the role of hydrolysates in the field of pharmacologic nutrition.
Materials and Methods
Microorganism, cultivation conditions, and biomass processing
Algae samples were obtained by autotrophic outdoor cultivation of C. vulgaris 87/1 in 500-m2 open circulating cascade systems. This strain was isolated from Chalons Dam in Santiago de Cuba, Cuba, in 1987 and is deposited at the Culture Collection of the Solar Energy Research Center. The growth medium contained NH4NO3 (1.2 g/L), MgSO4·7H2O (1.0 g/L) and a foodgrade nitrogen, phosphorous, and potash (8:12:12) fertilizer formula (0.9 g/L). The algal suspension was bubbled with 1% CO2. The algae were harvested by continuous-flow centrifugation (separator, Alfa Laval) up to 10% dry matter in the slurry. The dark-green algae slurry was spray-dried in a Niro Atomizer drier (input, 200–210°C; output, 80–90°C). The powder obtained (moisture content 7%) was preserved in plastic boxes for further use. Dry algae samples of 500 g were extracted with ethanol (2 L) at 45°C for 3 hours via gentle agitation.
Enzymatic hydrolysis of cell biomass
Pancreatin (Merck), with a specific proteolytic activity of 0.47 arbitrary units (AU)/mg of protein, was used for cell protein hydrolysis of the extracted algal biomass. One proteolytic unit was expressed as the amount of enzyme necessary to catalyze at an initial rate of the release of 1 μmol tyrosine from a 2% denatured casein solution at a pH of 7.5 and a temperature of 37°C within 1 minute.
A 10% suspension in water of the ethanol-extracted alga was hydrolyzed at an enzyme/substrate ratio of 30 AU/g, a pH of 7.5, and a temperature of 45°C for 4 hours in a 1,000-mL reaction vessel equipped with a stirrer, thermometer, and pH electrode. The enzyme reaction was stopped by heat treatment at 85°C for 15 minutes. The slurry thus obtained was centrifuged and the resultant solutions were spray-dried.
The yields of the technology were 40–45 g of hydrolysate/100 g of cell biomass. The bulk of the product dry matter consists of soluble hydrolyzed protein and free amino acids, which accounted for 47.7% of the yield. Three main peptides with molecular weights lower than 5,000 Da were identified by size exclusion chromatography. The amino nitrogen/total nitrogen ratio of 26.4% was considered appropriate for protein assimilation. The amino acid pattern was similar to that of the Food and Agriculture Organization reference protein, except for the low content of sulfur amino acids. 13
Animals and diets
Female Balb/c mice, weighing 20 g, were purchased from the National Center for the Production of Laboratory Animals (CENPALAB, Havana, Cuba) and housed individually at 23°C with a 12-hour/12-hour light/dark cycle. In a recent report, we proposed the use of starved female Balb/c mice as an experimental model of malnutrition for evaluating the efficacy of nutritional and immunopharmacologic interventions. 17
Thirty mice that were starved for 3 days and had free access to salted water were studied. After this time, blood was collected from the orbital vein of 10 mice and the animals were killed (M group). The others were refed ad libitum for 8 days with commercial pelleted diet (Ratonina®, CENPALAB) that had a protein content of 22% (M-DC group) or with the commercial diet and the Chlorella protein hydrolysate administered orally at a dose of 500 mg/kg of body weight per day (M-Cv-PH group). A control group of 10 mice was fed with commercial diet throughout the study. All experiments were approved by the institutional ethical committee (University of Oriente, Santiago de Cuba, Cuba) and were performed in accordance with Cuban legislation and the National Research Council Guidelines for the Care and Use of Laboratory Animals.
Biochemical analysis
Serum
Serum was prepared from collected blood and stored at −20°C until required. Total serum proteins were measured by a Biuret colorimetric assay with bovine serum albumin used as a standard, albumin was assessed by the colorimetric reaction with the bromocresol green reagent, and globulins were determined as the difference between total proteins and albumin. 18 Serum free amino acids were determined by reverse-phase high-performance liquid chromatography after derivatization with phenyl-isothiocyanate. 19
Liver
Liver samples were homogenized in ice-cold 0.01 mol/L phosphate-buffered saline with a pH of 7.4 (1:3 w/v). Total protein was measured according to the method of Lowry et al. 20 Cholinesterase and arginase activities were assayed by using the methods of de la Huerga and Mansurova, respectively. 21
Briefly, cholinesterase activity was determined by estimating the remnant substrate acetylcholine in the colorimetric reaction with hydroxylamine in the presence of ferric chloride. For measuring arginase activity, the remnant arginine was determined through the colorimetric reaction with α-naphtol and bromine. The enzyme-specific activities were expressed as micromoles of transformed substrate in 1 minute per mg of protein.
Gut mucosa
After the small intestine was collected, the segment correspondent to jejunum was rinsed thoroughly with ice-cold saline solution, opened, and blotted dry. The mucosa was scraped with a glass slide and weighed separately. Jejunal mucosa was homogenized with ice-cold phosphate-buffered saline with a pH of 6.0 (1:3 w/v). Total protein and DNA were quantified by the methods of Lowry et al. 20 and Burton, 22 respectively. Oligosaccharidases (sucrase and maltase) were determined according to the method of Dahlquist. 23 The enzymatic activities were expressed as micromoles of substrate hydrolyzed per minute, and the results were given per 10 cm of intestine segment.
Statistical analysis
Data are expressed as means±standard error. One-way analysis of variance and post hoc Tukey tests were used to determine mean differences among the groups for all the variables studied (P<.05). All data were analyzed by using SPSS software, version 12.0/ 2003 for Windows (SPSS Inc.).
Results and Discussion
Malnutrition induced by dietary restriction and severe starvation produces a series of metabolic changes that lead to reduction in body weight, depression of immunocompetence, and alteration of digestive system function, particularly of the liver and small intestine. 16,24
Starvation for 72 hours produced an average weight loss of 25%. Mice gradually recovered their weight during refeeding, regardless of the diet fed. Weight increased significantly (P<0.001) to a mean±standard error of 19.9±0.9 g in the M-DC group and of 21.0±1.2 g in the Cv-PH group, from 14.5±1.2 g in fasted animals. In fact, mice gained 0.55±0.1 and 0.6±0.2 g of body weight/ 24 hours during the 8-day refeeding period in the M-DC and M-Cv-PH groups, respectively. The average water and food consumptions were similar in both refed groups (data not shown). The average body weight of the control group was 20.3±0.6 g. Boza et al. 24 reported an average weight loss of 14.5% in 200-g rats starved for 3 days; nevertheless, control and refed rats did not differ in net protein utilization or biological value.
Figure 1 shows hemoglobin concentrations in total blood. The hemoglobin levels decreased after 3 days of fasting (P<.01). Oral refeeding of fasted mice with a Cv-PH diet led to a significant increase in hemoglobin concentration; levels were higher than those of the M-DC and control groups. In a clinical study performed with 32 patients with differing degrees of chronic renal insufficiency, an increase in hemoglobin values (P<.05) was observed after 120-day treatment with 100 g of an oral preparation formulated with a protein hydrolysate of Spirulina pacifica supplemented with natural bee honey as a source of energy. 8 In a previous paper, Morris et al. 14 reported that hemopoiesis was favorably affected at the end of the oral treatment of undernourished mice with Cv-PH.

Effect of starvation and refeeding with commercial diet supplemented or not supplemented with Chlorella vulgaris protein hydrolysate on hemoglobin concentration in total blood of Balb/c mice. All values are given as the arithmetic mean±standard error of 10 mice. abcDifferent letters indicate significant differences among the groups according to the Tukey test (P<.01). C, control mice; M, fasted mice; M-Cv-PH, mice fed the commercial diet and the Chlorella protein hydrolysate; M-DC, mice fed the commercial diet.
Alterations in serum proteins are implicated in protein-energy malnutrition pathogenesis; in particular, albumin has been recognized as a good marker of nutritional status. 25,26 Table 1 summarizes the serum protein concentrations in all groups. Significant hypoalbuminemia was observed in starved mice (P<.05); this finding supports the role of albumin as a temporal reservoir of amino acids and in the transport of amino acids and fatty acids to peripheral tissues. Refeeding restored albumin levels to control group values, regardless of the diet fed. Nonsignificant differences occurred between groups with respect to the total serum protein concentration, globulin concentration, and albumin-to-globulin ratio. Globulins are often increased when albumin concentration is diminished; because these protein fractions had opposite changes, the total serum protein concentration may not reflect the nature and extension of certain alterations. 27
Serum was prepared from collected blood and stored at −20°C until required. Total serum proteins were measured by using a Biuret colorimetric assay with bovine serum albumin as a standard; albumin was assessed by using the colorimetric reaction with the bromocresol green reagent; and globulins were determined as the difference between total proteins and albumin. All values are expressed as the arithmetic mean±standard error of 10 mice.
P<.05.
Different superscripted letters indicate significant differences among the groups according to the Tukey test.
M, fasted mice; M-Cv-PH, mice fed the commercial diet and the Chlorella protein hydrolysate; M-DC, mice fed the commercial diet.
Although the supplementary effect of Cv-PH on serum protein levels was not significant compared with the effect in the M-DC group, it would be possible to measure the concentrations of transport proteins with a lower half-life time than albumin, for which hepatic synthesis may be limited by an inadequate amino acid availability; such proteins include transferrin, prealbumin, and retinol-binding protein. 16
Serum free amino acid concentrations, in general, decreased with fasting (Table 2). After refeeding, the total amino acid concentrations, as well as the concentrations of essential and nonessential amino acids, were significantly higher in Cv-PH–treated animals and reached the values of control mice. Total serum levels of branched-chain amino acids and the ratios of nonessential to essential amino acids were similar in all experimental groups. Regardless of diet, most serum amino acid concentrations increased during refeeding. The concentration of serum glutamic acid plus glutamine was lower and serum arginine and lysine values were higher in Cv-PH–treated mice.
Serum free amino acids were determined by reverse-phase high-performance liquid chromatography after derivatization with phenyl-isothiocyanate, according to the method described by Scholze. All values are expressed as the arithmetic mean±standard error of 10 mice.
P<.05
P<.01
P<.001.
Different superscript letters indicate significant differences among the groups according to the Tukey test.
BCAA, branched-chain amino acids; EAA, essential amino acids; M, fasted mice; M-Cv-PH, mice fed the commercial diet and the Chlorella protein hydrolysate; M-DC, mice fed the commercial diet; NEAA, nonessential amino acids.
The lower serum glutamic acid plus glutamine concentration in the Cv-PH group (P<.001) could be explained by the fact that this amino acid is the main energy substrate for the small intestine during intestinal recovery. 28 In addition, protein anabolism in muscle is enhanced during nutritional recovery after starvation, which leads to lower synthesis of glutamine from alanine. 29,30
On the other hand, the arginine concentration was higher in mice supplemented with Cv-PH (P<.05). Arginine is considered a semi-essential amino acid (mainly during stress), and it has several unique metabolic effects, such as serving as a substrate for nitric oxide synthesis, stimulating the pancreas to secrete insulin and glucagon, improving nitrogen retention, and enhancing cell-mediated immune function. 31
The serum concentrations of the distinct amino acids cannot be generalized because of their different precursor functions in the amino acid pool and the complex metabolic and immunomodulating implications for preserving the homeostasis.
Starvation for 72 hours decreased liver weight (3.43±0.05 g/100 g of tissue). Liver weight in the Cv-PH group was higher than that in the M-DC group (4.90±0.09 vs. 4.37±0.07 g/100 g of tissue; P<.05) and reached the values of control animals (5.26±0.08 g/100 g of tissue). Table 3 presents the results of liver protein content and enzyme-specific activities in all groups. Starved mice showed significantly lower protein concentrations (P<.01) and cholinesterase activity (P<.05) and a greater increase in arginase activity (P<.01) than did control animals. After refeeding in the Cv-PH group, liver function returned to normal and liver measures reached the values of control mice. These findings were not seen in the M-DC animals.
Liver samples were homogenized in ice-cold 0.01 mol/L phosphate-buffered saline with a pH of 7.4 (1:3 w/v). Total protein was measured according to the Lowry method. Cholinesterase activity was determined by estimating the remnant substrate acetylcholine in the colorimetric reaction with hydroxylamine in the presence of ferric chloride. For measuring arginase activity, the remnant arginine was determined through the colorimetric reaction with α-naphtol and bromine. The enzyme-specific activities were expressed as micromoles of transformed substrate in 1 minute per milligram of protein. All values are expressed as the arithmetic mean±standard error of 10 mice.
P<.05.
P<.0.01.
Different superscript letters indicate significant differences among the groups according to the Tukey test.
M, fasted mice; M-Cv-PH, mice fed the commercial diet and the Chlorella protein hydrolysate; M-DC, mice fed the commercial diet.
Cholinesterase activity reflects liver function. Decreases in this measure suggest alterations in protein synthesis and have been associated with protein-energy malnutrition. 32 In addition, modifications of individual amino acid catabolism in relation to protein intake are associated with changes in urea synthesis. In our study, we determined the activity of arginase; this enzyme involved in the catalysis of the final reaction of urea cycle, which operates specifically in hepatocytes. 21 The results indicated the stimulation of liver protein anabolism and the reduction in urea synthesis from amino groups of amino acids, favoring the restoration of normal liver metabolism in Cv-PH–supplemented mice.
The administration of a milk protein hydrolysate to healthy volunteers led to increased expression of glucose-dependent insulinotropic polypeptide. 33 It has been proposed that insulin acts as a signaling mechanism that in hepatocytes initiates the changes induced by protein hydrolysates, favoring the stimulation of protein synthesis and the inhibition of protein catabolism. 34 According to those authors, the association of increased concentrations of serum amino acids and insulin action would explain the advantages of hydrolysates compared with native proteins in stimulating more efficient use of nitrogen.
The intestinal tract is an important interface between the organism and the environment. It helps regulate incorporation of nutrients and defend against pathogens. In protein-energy malnutrition, the adaptive responses and defense mechanisms of the gut mucosa are altered. 35
After 72 hours of food deprivation, fasted mice (group M) showed decreases in mucosal weight, mucosal DNA and protein contents, and jejunal oligosaccharidase activities (Figs. 2 and 3). Refeeding led to a significant increase in mucosal weight (P<.01), similar to the weight of control animals in the M-DC group and higher than the weight in Cv-PH–treated mice. Normal function and turnover of gut mucosa epithelial cells require a high rate of protein synthesis. Recovered M-DC mice exhibited a trend toward increased DNA and protein contents, but only Cv-PH–refed animals reached the mucosal protein concentrations of control animals; in particular they showed higher DNA gut mucosal content (P<.01).
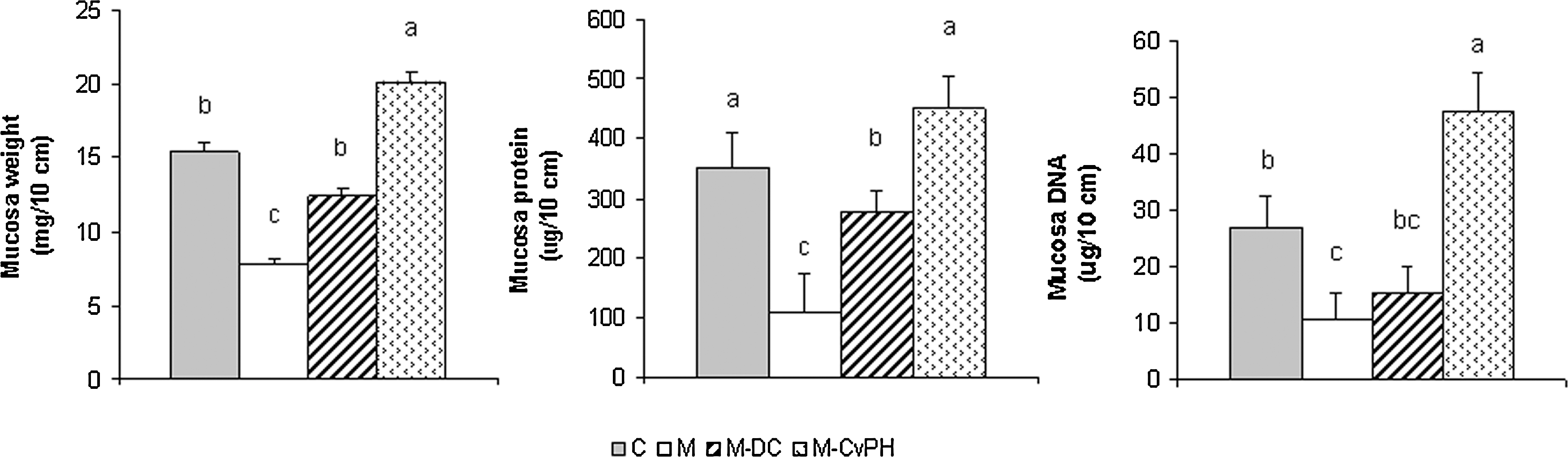
Effect of starvation and refeeding with commercial diet supplemented or not supplemented with Chlorella vulgaris protein hydrolysate on mucosal weight, protein mucosal content, and DNA content in the jejunum of Balb/c mice. After the small intestine was collected, the segment correspondent to the jejunum was rinsed thoroughly with ice-cold saline solution, opened, and blotted dry. The mucosa was scraped with a glass slide and weighed separately. Jejunal mucosa was homogenized with ice-cold phosphate-buffered saline with a pH of 6.0 (1:3 w/v). Total protein and DNA were quantified by the methods of Lowry and Burton, respectively. Results are expressed per 10 cm of intestine. All values are given as the arithmetic mean±standard error of 10 mice. abcDifferent letters indicate significant differences among the groups according to the Tukey test (P<.01). M-Cv-PH, mice fed the commercial diet and the Chlorella protein hydrolysate.
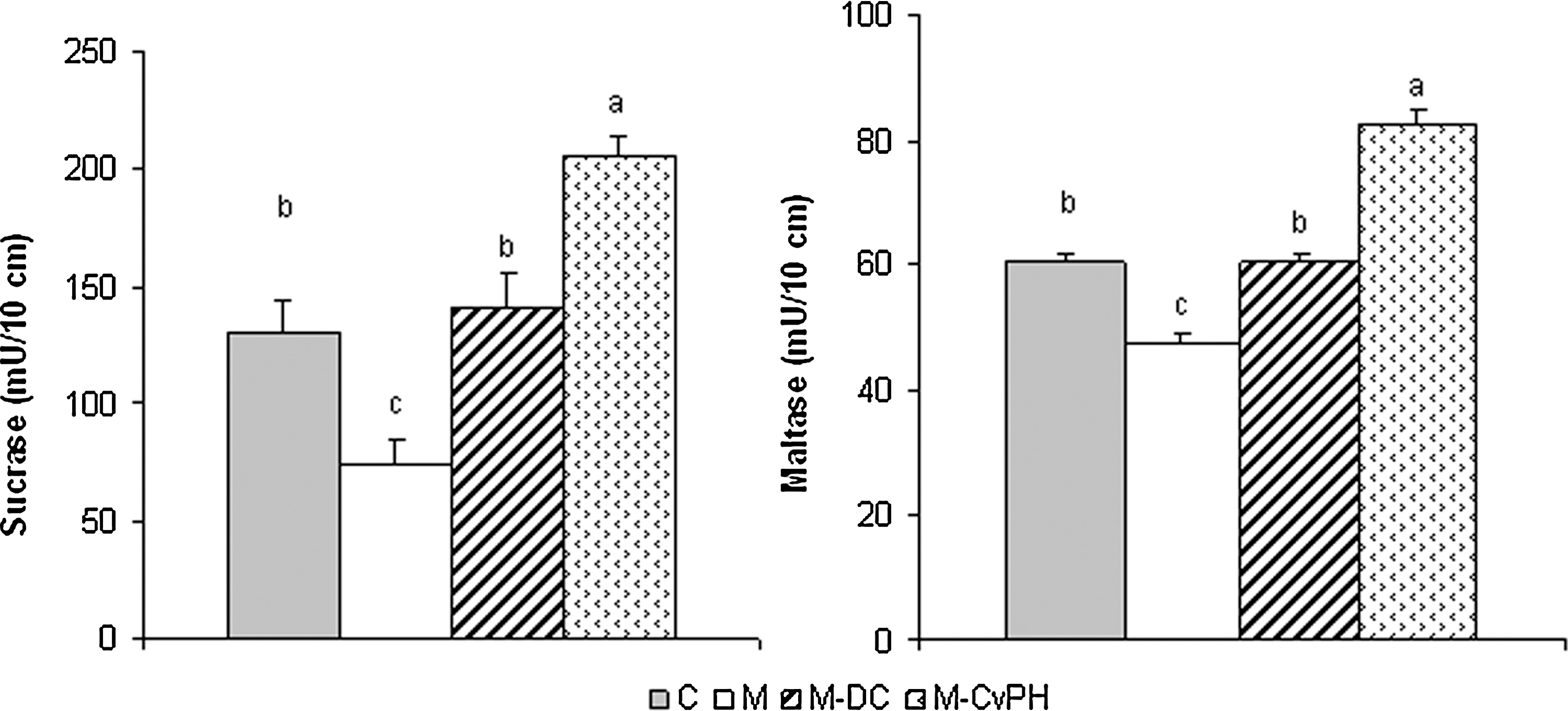
Effect of starvation and refeeding with commercial diet supplemented or not supplemented with Chlorella vulgaris protein hydrolysate on oligosaccharidase activities in jejunum of Balb/c mice. After the small intestine was collected, the segment correspondent to the jejunum was rinsed thoroughly with ice-cold saline solution, opened, and blotted dry. The mucosa was scraped with a glass slide and weighed separately. Jejunal mucosa was homogenized with ice-cold phosphate-buffered saline with a pH of 6.0 (1:3 w/v). Oligosaccharidases (sucrase and maltase) were determined according to the method of Dahlquist. The enzymatic activities were expressed as micromoles of substrate hydrolyzed per minute and the results were given per 10 cm of intestine segment. All values are given as the arithmetic mean±standard error of 10 mice. abcDifferent letters indicate significant differences among the groups according to the Tukey test (P<.01).
The increased DNA content observed in jejunal mucosa of mice supplemented with Chlorella protein hydrolysate might be associated with the stimulation of cell division and turnover of enterocytes. 36 Diets formulated with protein hydrolysates supply high concentrations of specific bioactive peptides that modulate digestive (on absorption and nitrogen retention) and metabolic events in the gastrointestinal tract. 37 Cv-PH administered orally to malnourished mice produced trophic effects on gut mucosa, presumably by means of Chlorella peptides released by enzymatic hydrolysis.
Pancreatic and brush-border proteolytic activities are indispensable for the production of bioactive peptides. After starvation, these activities are depleted; therefore, the production of these peptides would be more difficult for animals receiving an intact protein diet (i.e., Chlorella powder or extracts).
Malnourished mice, as part of the response to a diet deficient in both protein and energy, had reduced brush-border enzyme activities; this change was associated mainly with the nonspecific loss of mucosal proteins instead of the absence of particular substrates. 38
The levels of oligosaccharidases (sucrase and maltase) were significantly increased (P<.01) after refeeding and reached control values; in particular, the Cv-PH group had greater oligosaccharidase activity. Kops et al. 39 reported the stimulation of the brush-border enzyme activities of alkaline phosphatase and maltase in the enterocytic differentiation cell line C2BBe cultured in the presence of soy protein hydrolysates used in enteral nutrition.
Starvation not only severely impairs small-intestine digestive capacity but also increases intestinal permeability to macromolecules, such as ovalbumin. The damaged mucosa may enhance antigen translocation, which leads to immune-induced mucosal damage mediated by inflammatory and allergic reactions. 24,29 On the basis of these considerations, we can hypothesize that during refeeding, the restoration of small-intestine mucosa with Cv-PH helps prevent triggering of an immunogenic response at local or systemic levels. Given the molecular distribution of peptides in Cv-PH (3 main peptides with molecular weights <5,000 Da), 13 its uptake from the gut should not cause major problems.
In sum, the results of this study show that enzymatic protein hydrolysates from Chlorella vulgaris, administered orally to mice recovering from severe starvation, provide nutritional benefits in terms of serum amino acid profiles, hemoglobin concentration, and liver and small-intestine functions. These findings, together with our previous results on the immunostimulant activity of Cv-PH, 14 suggest its potential use in the development of functional foods and specific formulations suitable for pharmacologic nutrition.
Footnotes
Author Disclosure Statement
No competing financial interests exist.