
Abstract
Cisplatin (CDDP), one of the most active cytotoxic agents against cancer, has adverse side effects, such as nephrotoxicity and hepatotoxicity. The present study was designed to investigate the potential protective effect of pomegranate seed extract (PSE) against oxidative stress caused by CDDP injury of the kidneys and liver by measuring tissue biochemical and antioxidant variables and immunohistochemically testing caspase-3–positive cells. Twenty-four Sprague–Dawley rats were divided into 4 groups: control; CDDP: injected intraperitoneally with CDDP (7 mg/kg body weight, single dose); PSE: treated for 15 consecutive days by gavage with PSE (300 mg/kg per day); and PSE+CDDP: treated by gavage with PSE 15 days after a single injection of CDDP. The degree of protection against CDDP injury afforded by PSE was evaluated by determining the levels of malondialdehyde as a measure of lipid peroxidation. The levels of glutathione and activities of glutathione peroxidase, glutathione S-transferase, and superoxide dismutase were estimated from liver and kidney homogenates; the liver and kidney were also histologically examined. PSE elicited a significant protective effect toward liver and kidney by decreasing the level of lipid peroxidation; elevating the levels of glutathione S-transferase; and increasing the activities of glutathione peroxidase, glutathione S-transferase, and superoxide dismutase. These biochemical observations were supported by immunohistochemical findings and suggested that PSE significantly attenuated nephrotoxicity and hepatotoxicity by the way of its antioxidant, radical-scavenging, and antiapoptotic effects. This PSE extract could be used as a dietary supplement in patients receiving chemotherapy medications.
Introduction
C
The kidneys and liver are very important organs for continuation of homeostasis, and their dysfunction can arise from the administration of various drugs or chemicals, such as some antibiotics and antineoplastic drugs. Cis-diamminedichloroplatinum II (cisplatin, or CDDP) is one of the most important cytostatic agents for the treatment of a broad range of solid tumors. 3,4 Damage to kidney and liver function by CDDP is recognized histologically as necrosis of the terminal portion of the proximal tubule and apoptosis in the distal nephron of the kidney 5 –7 and as sinusoidal dilatation and congestion, hepatocellular degeneration, and inflammatory infiltrations in the liver. 3 –9 This damage becomes the most important dose-limiting factor for CDDP.
Cummings and Schnellmann 10 reported that at least 50% of CDDP-induced apoptosis in proximal tubular cell is mediated by p53 activates caspase 3, whereas 50% of CDDP-induced apoptosis is independent of caspases 3, 8, and 9 and of p53 or mitochondrial dysfunction. Apoptosis is programmed cell death, characterized by activation of caspases, chromatin condensation, cell shrinkage, membrane budding, and phosphatidylserine externalization. Apoptosis was demonstrated by caspase 3 activation, annexin V labeling, chromatin condensation, DNA damage, and cell morphology. Caspase activation is a significant step in the formation of apoptosis. 11 Caspase 3 can be activated by caspase 9, which is activated by the release of cytochrome c from the mitochondria 11,12 or p53-mediated activation of caspase 8 and 9 in the mitochondria. 13
Toxic liver and renal failures are known to induce the excessive production of reactive oxygen species, which are responsible for the induction of kidney and hepatic cell death. 14 –17 However, many studies now indicate that consumption of fruits and vegetables rich in a number of phytochemicals can reduce these side effects of reactive oxygen species and, presumably, of CDDP. 7,18 –20 Research has shown that compounds in fruits have cytoprotective and antioxidant activity and can be effective when exogenously administered. Antioxidants such lycopene, 19 ellagic acid, 21 pomegranate flowers, 22 L-carnitine, 9 caffeic acid phenethyl ester, 23 vitamin C, 24 vitamin E, 25 and resveratrol 26 have shown protective effects against cytotoxic agents in both in vivo and in vitro trials. 27,28
In the present study, we investigated the protective effect of pomegranate seed extract (PSE) against oxidative stress caused by CDDP injury of the kidneys and liver by measuring biochemical variables and conducting immunohistochemical examinations for apoptotic cells in Sprague-Dawley rats.
Material and Methods
Animals and treatments
Adult female Sprague-Dawley rats (mean weight±standard deviation, 180±20 g; 6–8 weeks old) were provided by the Atatürk University's Experimental Research Centre, Erzurum, Turkey. During the study, the animals were kept in metal cages at a temperature of 22°C–24°C and with a 12-hour light/dark cycle; they were fed with standard commercial rat food and tap water. All experiments in this study were approved by the Local Ethics Board of Animal Experiments in Atatürk University. The animals were divided into 4 groups, each with 6 rats, according to their experimental treatment: the control group, the group receiving PSE (PSE group), the group receiving CDDP (CDDP group), and the group receiving both PSE and CDDP (PSE+CDDP group).
The rats in the control group were fed with standard rat food and tap water for 15 consecutive days. The rats in the PSE group were given PSE (300 mg/kg per day orally; Balen Pomegranate Seed Extract Capsule, Arı Mühendislik Co.) through an orogastric tube for 15 consecutive days. The rats in the CDDP group received a single-dose injection of CDDP (7 mg kg/body weight intraperitoneally; Ebewe and Liba) all at once. The rats in the PSE+CDDP group received single oral doses of PSE (300 mg/kg per day orally) for 15 consecutive days after a single-dose intraperitoneal injection of CDDP (7 mg/kg).
Sample Collection and Biochemical Assays
Twenty-four hours after the final PSE and CDDP treatments, all animals were anesthetized with an intraperitoneal injection of 60 mg sodium pentobarbitone per kg of body weight and then sacrificed by cervical dislocation. The livers and kidneys were removed, washed with physiologic saline solution, and stored at −80°C until analysis. All tissues were maintained at 4°C throughout preparation. A portion of the liver and kidney tissues (1:9, w/v) for all assays was homogenized in a 0.9% NaCl solution with an OMNI TH International homogenizer (Warrenton). Tissue homogenates were centrifuged for 15 minutes at 15,000 g, and then the clear upper supernatants were removed for analyses.
Determination of malondialdehyde
Malondialdehyde (MDA) levels in tissues were determined spectrophotometrically according to the method described by Ohkawa et al. 29 A mixture of 8.1% sodium dodecyl sulfate, 20% acetic acid, and 0.9% thiobarbituric acid was added to 0.2 mL of each sample, and the volume was brought up to 4 mL with distilled water. After incubation at 95°C for 1 hour, the tubes were left to cool in cold water; 1 mL distilled water plus 5 mL n-butanol/pyridine (15:1, v/v) were added and mixed. The samples were then centrifuged at 4,000 g for 10 minutes; the supernatants were removed and used for absorbance measurements with respect to a blank at 532 nm. 1,1,3,3-Tetraethoxypropane was used as the standard. Lipid peroxide levels were expressed as nmol/L MDA.
Determination of glutathione
Glutathione (GSH) levels in tissues were assessed by using methods described by Tietze 30 and Anderson. 31 Briefly, 100 μL of each sample was placed into a 3-mL cuvette, and then 750 μL of 10 mM 5-5′-dithiobis-2-nitrobenzoic acid (DTNB) solution (100 mM KH2PO4 plus 5 mM Na2EDTA [pH, 7.5] and reduced GSH, 625 U/L) was added, and the samples were incubated for 3 minutes at room temperature. Then, 150 μL of 1.47 mM β-nicotinamide adenine dinucleotide phosphate (NADPH) was added and mixed rapidly by inversion. The rate of 5-thio-2-nitrobenzoic acid formation (proportional to the sum of reduced and oxidized glutathione) was measured spectrophotometrically for 2 minutes at 412 nm. The reference cuvette contained equal concentrations of DTNB and NADPH but no sample, and values are presented as μmol per g protein.
Determination of GSH peroxidase activity
GSH peroxidase activity in tissues was measured by using the method of Paglia and Valentine. 32 Briefly, 50 μL of sample was combined with 100 μL of 8 mM NADPH, 100 μL of 150 mM reduced GSH, 20 μL of GSH reductase (30 units/mL), 20 μL of 0.12 M sodium azide solution, and 2.65 mL of 50 mM potassium phosphate buffer (pH, 7.0; 5 mM EDTA). The tubes were incubated for 30 minutes at 37°C. The reaction was initiated with the addition of 100 μL of 2 mM H2O2 solution and mixed rapidly by inversion. The conversion of NADPH to NADP was measured spectrophotometrically for 5 minutes at 340 nm. The enzyme activity was expressed as units per g protein using an extinction coefficient for NADPH at 340 nm of 6.22×10−6.
Determination of GSH-s-transferase activity
GSH-s-transferase (GST) activity was assayed by using the electrophilic substrate 1-chloro-2,4- dinitrobenzene (CDNB) according to the procedure described by Habig et al. 33 GST was estimated in 1 mL of incubation mixture containing 905 μL of 0.1 M phosphate buffer (pH, 6.5), 20 μL of 20 mM CDNB reagent, 25 μL of 200 mM of reduced GSH, and 25 μL of TritonX100 (0.66%). The mixture was preincubated at 37°C for 5 minutes. The reaction was started by adding 25 μL of the sample. Enzyme activity was determined by continuously monitoring the change in absorbance at 340 nm for 3 minutes. The optical density change/min was calculated, and GST activity was estimated by using the molar extinction coefficient (9.6 mM−1 cm−1) of GST.
Determination of superoxide dismutase activity
Cu, Zn-super oxide dismutase (SOD) activity in tissues was measured by the method of Sun et al. 34 A 2.45-mL volume of assay reagent (0.3 mM xanthine, 0.6 mM Na2EDTA, 0.15 mM nitroblue tetrazolium, 0.4 M Na2CO3, and 1 g/L bovine serum albumin) was combined with 100 μL of tissue homogenate. Xanthine oxidase (50 μL, 167 U/L) was added to initiate the reaction, and the reduction of nitroblue tetrazolium by superoxide anion radicals, which are produced by the xanthine-xanthine oxidase system, was determined by measuring the absorbance at 560 nm. Cu, Zn-SOD activity was expressed in units of SOD per mg protein, where 1 U is defined as that amount of enzyme causing half-maximal inhibition of nitroblue tetrazolium reduction.
Immunohistochemistry evaluation
For light microscopy, the livers and kidneys were fixed in 10% formalin and embedded in paraffin. The paraffin blocks were cut 5- to 7-μm thick and stained with Mallory's triple stain modified by Crossman. Apoptotic cells were determined with the streptavidin-biotin-peroxidase staining method. For immunohistochemistry examination we used primary antibody monoclonal caspase 3 (dilution: 1/10, Biovision-3015-100) and biotinylated secondary antibody (DAKO-Universal LSAB Kit-K0690); binding sites of antibody were visualized with diaminobenzidine (Sigma). The binding of antibodies was evaluated by high-power light microscopic (Nikon i50) examination.
Thereafter, degenerative changes and caspase-3 immunoreactivity were examined in 10 randomly selected areas of approximately ×20 objective with an image analysis program (Kameram SLR, 1.4.1.0, Mikro Sistem Ltd.). Sections were microscopically scored by a histopathology laboratory technician and histologist. This scale consisted of the following: A=weak in 25% of tissue or less; B=mild in 50% of tissue or less; C=moderate in 75% of tissue or less; and D=very strong in 75% of tissue or more. The average degeneration intensity was calculated as [(A×1)+(B×2)+(C×3)+(D×4)]/(A+B+C+D) and reported as follows: +=0.00–1.00; ++=1.01–2.00; +++=2.01–3.00; and ++++=3.01–4.00. The scores were derived semi-quantitatively by using light microscopy on the preparations from each animal and were reported as follows: −=none; +=mild;++=moderate; +++=severe; and ++++=very strong.
Statistical analysis
For statistical analysis, differences between the groups were tested by analysis of variance followed by the Duncan post hoc test using SPSS software, version 11.0, for Windows XP (SPSS Inc.). A P value less than .05 was considered to represent a statistically significant difference. All data were expressed as means±standard error.
Results
Biochemical results
Table 1 shows the changes in MDA and GSH levels, and GSH peroxidase, GST, and SOD activity in the livers and kidneys. Compared with the control groups, the levels of MDA, a major degradation product of lipid peroxidation, were significantly (P<.05) higher in groups administered CDDP. This increase was attenuated by pretreatment with PSE. Significantly (P<.05) reduced levels of GSH and lower GSH peroxidase, GST, and SOD activities were seen in the liver and renal tissues of CDDP-treated rats compared with the control group. PSE pretreatment alleviated these CDDP-induced decreases (Table 1). Administration of PSE alone had no statistically significant effects on any measures compared to the control group.
Values are expressed as means±standard error.
The footnotes a, b, and c in the same column indicate significant differences between groups (n=6); P<.05.
CDDP, cisplatin; GSH, glutathione; GSHPx, glutathione peroxidase; GST, glutathione S-transferase; PSE, pomegranate seed extract; SOD, superoxide dismutase.
Histologic results
Table 2 presents morphologic and histologic changes. The livers and kidneys of the control animals and the PSE-only groups showed normal histologic structure (Fig. 1 and Fig. 2). In immunohistochemical examination, caspase 3–positive cells were located around of the central vein from the livers (Fig. 1) and in the proximal tubules and loop of Henle from the kidneys (Fig. 2) of the CDDP group. The number of apoptotic cells increased in the livers and kidneys of the CDDP group when compared with other experimental groups. In the CDDP group, histologic alterations included sinusoidal dilatation, congestion, and hepatocellular degeneration. In addition, degeneration and desquamation in the proximal and distal tubules in the kidney sections were seen. In the CDDP+PSE group, decreases in apoptotic cell numbers and sinusoidal dilatations around the central vein were seen compared with the CDDP group. Moreover, in the kidney sections, PSE significantly caused curable degenerative changes and decreased the caspase 3 activity in proximal tubules and loop of Henle (Fig. 2).
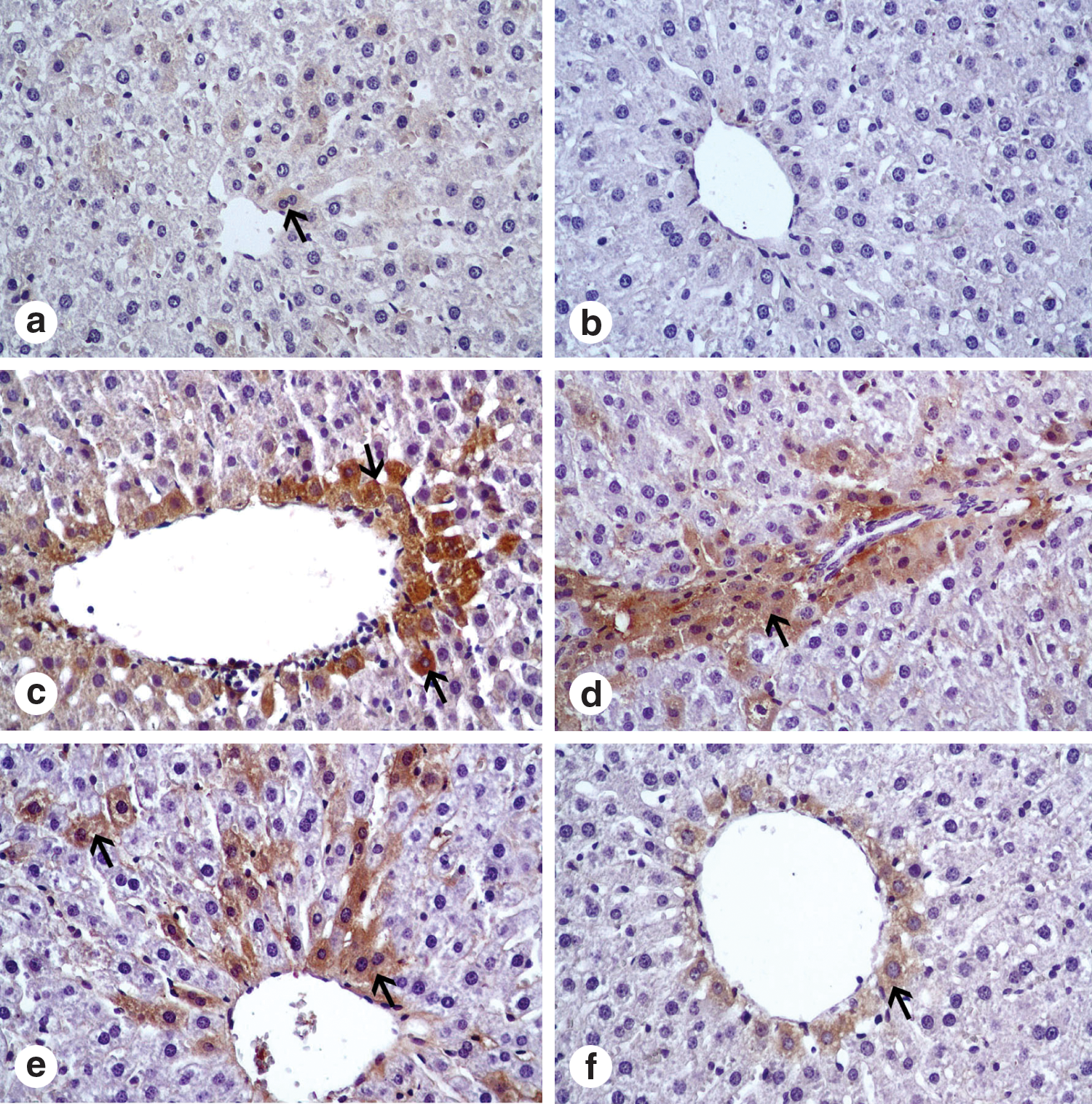
Apoptotic cells in liver. (
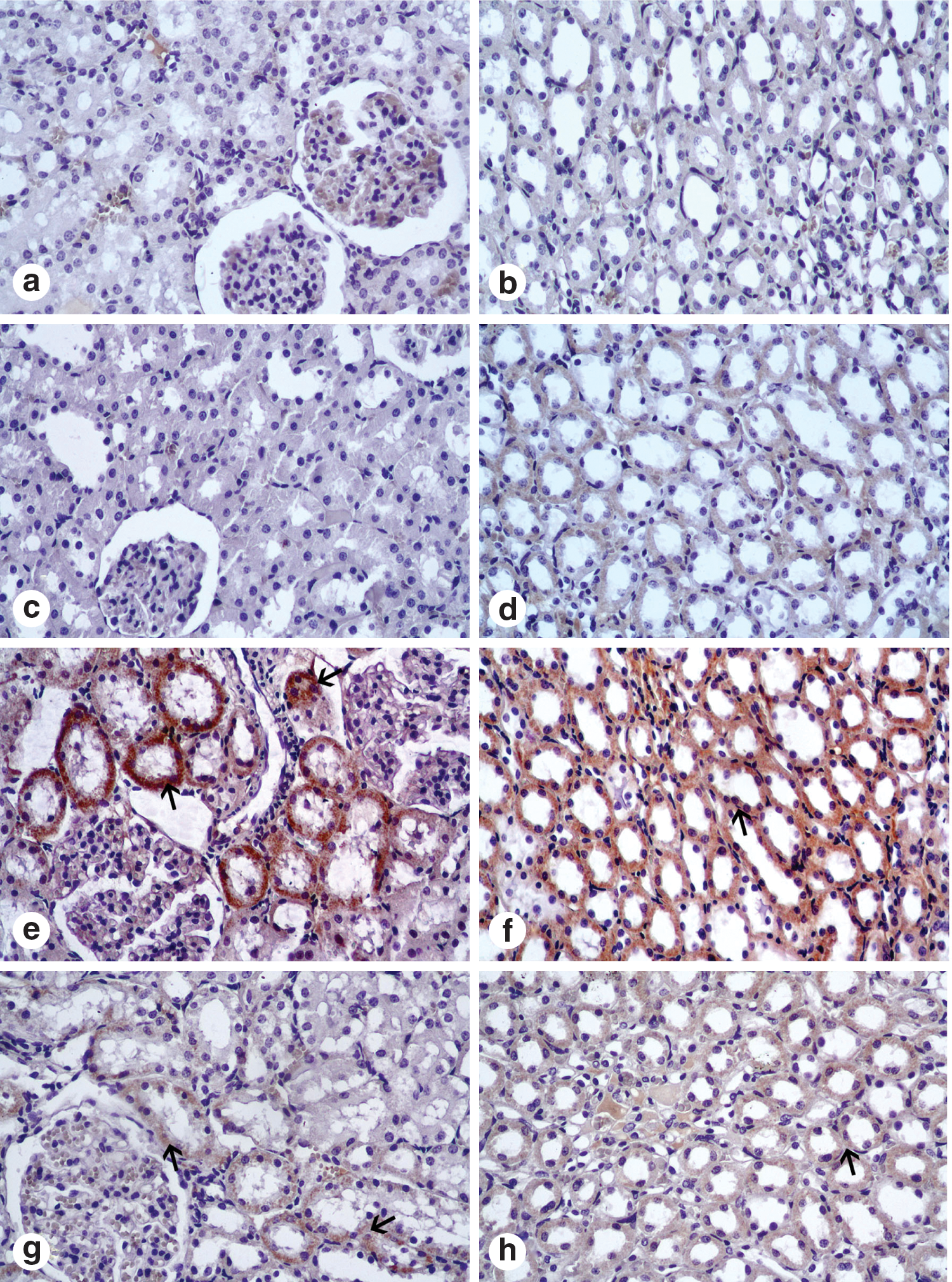
Apoptotic cells in cortex and medulla of kidney. (
Each group consisted of 6 rats. Average degeneration and caspase 3 reaction density were estimated as [(A×1)+(B×2)+(C×3)+(D×4)]/(A+B+C+D) and reported as follows: +=0.00–1.00; ++=1.01–2.00; +++=2.01–3.00; ++++=3.01–4.00. A=weak, ≤25% of tissue; B=mild in ≥25−≤ 50% of tissue; C=moderate in ≤75% of tissue, D=very strong in ≥75%–100% of tissue.
Discussion
CDDP is a platinum-based chemotherapy drug used to treat various types of cancers, such as sarcomas, some carcinomas, lymphomas, and germ cell tumors. Unfortunately, many cancer cells are resistant to this treatment or may become resistant with treatment. Currently, the only way to overcome acquired resistance to CDDP in cancer cells is to increase the dosage, which results in higher toxicity to normal body cells. 4 CDDP is significantly metabolized by liver, making liver toxicity also a major dose-limiting side effect in CDDP-based chemotherapy. 1 Some reports suggest that CDDP-induced hepatotoxicity may be dose-related. Zicca et al. 35 and Koc et al. 3 stated that administration of high doses of cisplatin (7.5 mg/kg and 10 mg/kg, respectively) do result in hepatotoxicity.
In the present study, hepatotoxicity was apparent in rats treated with a high dose (7 mg/kg) of CDDP, as evidenced both by the high readings for biochemical measures such as lipid peroxidation and the low antioxidant activity and by histologic changes. Many studies have reported structural changes in the liver similar to those reported here. 4,8,20 Nephrotoxicity of CDDP has also been documented as a highly important dose-limiting factor in cancer chemotherapy, as evidenced by the high lipid peroxidation and free radical generation in the tubular cells. These effects have been suggested to be responsible for CDDP nephrotoxicity. 4,6,7
Previous studies have reported that free radicals derived from oxidation-reduction reactions or from reactions mediated by reactive oxygen species are responsible for a wide range of chemotherapy-induced side effects, including kidney and liver toxicities. 5,6,9,18 CDDP can cause the generation of oxygen free radicals, such as hydrogen peroxide, superoxide anions, and hydroxyl radicals, in the kidney and liver by increasing the activity of cytochrome P450 enzymes, NADPH oxidase, xanthine oxidase, and adenosine deaminase. 36 These radicals can cause extensive tissue damage as they react with such macromolecules as membrane lipids, proteins, and nucleic acids in the kidneys and liver. For example, the hydroxyl radical is capable of abstracting a hydrogen atom from polyunsaturated fatty acids in membrane lipids to initiate lipid peroxidation. 37,38
In the present study, significantly higher levels of liver and kidney MDA were observed in the CDDP-treated group than in the controls. Increased MDA levels indicated that lipid peroxidation, mediated by reactive oxygen species, was an important contributing factor in the development of the observed CDDP-mediated tissue damage. Pretreatment with PSE significantly prevented CDDP-induced lipid peroxidation in the liver and kidney tissues, indicating an antioxidant effect of this extract. The decrease in lipid peroxidation was probably due to less damage from oxygen free radicals. In previous studies, PSE had antioxidant, 39 hypoglycemic, 40 antiapoptotic, 41 and peroxidative effects on blood and on such tissues as liver, kidney, and prostate.
Many natural products are reported to influence the antioxidant systems and are good cytoprotective agents. In general, GSH, GST, GSH peroxidase, and SOD play important roles in biological systems to protect against oxidative stress. 23,25,26,28 GSH is a ubiquitous thiol-containing tripeptide that plays a key role in cell biology. It modulates cell responses to redox changes associated with the generation of reactive oxygen species, detoxifies the metabolites of drugs (most often formed by cytochrome P450-linked monooxygenase), regulates apoptotic cell death, and mediates transmembrane transport of organic solutes. The significant reduction in GSH levels promoted by CDDP represents an alteration in the cellular redox state, suggesting that the cells could have become more sensitive to reactive oxygen species. This would lead to a reduction in effectiveness of the antioxidant enzyme defense system. 42 In the present study, GSH levels in the liver and kidney tissues were lower in rats treated with CDDP than in the control group. On the other hand, an increase in GSH levels in the liver and kidney tissues indicated that pretreatment with PSE prevented the loss of GSH in response to oxidative stress. The effects of PSE on cellular GSH may therefore be due directly to its antioxidant effects or may represent enhanced biosynthesis of GSH.
GST represents a large family of multifunctional proteins that are essential for disposal of exogenous toxic compounds and for adaptive antioxidant responses to ROS. This enzyme catalyzes the reaction of compounds with the sulfhydryl group of glutathione, creating products that are more water-soluble. GSH peroxidase has a pivotal role in H2O2 catabolism and catalyzes the reduction of H2O2 to H2O and O2 at the expense of GSH. 42 Superoxide radicals are formed in all living cells exposed to oxygen, by various biochemical systems within the cells. The SOD enzyme protects the organism against the toxic effects of superoxide radicals by catalyzing their dismutation to molecular oxygen and H2O2. 43 In the present study, animals treated with CDDP alone showed reduced kidney and liver GST, GSH peroxidase, and SOD activity compared with the control group. These observations suggested that the mechanism of kidney and liver toxicity induced by CDDP in animals is partially related to the depletion of kidney and liver antioxidants. Treatment with PSE immediately after a CDDP challenge significantly prevented this antioxidant depletion. Similar results have been reported by Chidambara Murthy et al., 44 Ahmed and Zaki, 45 Thring et al., 46 Ilbey et al., 47 Bouroshaki et al., 48 and Ahmed and Ali, 49 for kidney, liver, brain, and plasma tissues in which cisplatin, CCl4, ferric nitrilotriacetate, hyperoxaluria, and chlorpyrifos-ethyl treatments caused low GSH, GSH peroxidase, SOD, catalase and nitric oxide levels. In vitro studies 50 –52 also reported similar findings. In the present study, levels of GSH peroxidase and SOD in the PSE-treated groups were higher than those in the CDDP group. These results suggested that PSE has a supportive effect on the antioxidant system because of increases in GSH peroxidase and SOD levels. Pharmacologic properties of pomegranate extracts have been scrutinized, and increased fertility 53 and reduced cytotoxicity 51 and hypoglycemia 54 have been noted. The extracts also demonstrate antimicrobial, 55 anti-parasitic, 56 antiviral, 57 antifungal, 58 and antimutagenic 59 effects. The antioxidant activity of pomegranate extract is inferred to be a result of its component polyphenolics, such as ellagic acid and ellagitannins, 60 which may as act as electron donors in the scavenging of free radicals. 61
Some researchers reported that CDDP has an apoptotic effect on several tissues, such as in the proximal tubule of the kidney and hepatocytes. Apoptosis is characterized by phosphatidylserine externalization, membrane budding, cell shrinkage, chromatin condensation, and activation of caspases 3, 8 and 9. 62 Numerous studies have demonstrated that cisplatin induces both apoptosis and oncosis in kidney, but the mechanisms responsible for caspase 3 activation in apoptotic cells are not fully understood. 11,12,63 Some experiments have shown that p53 activates caspase 3 by mechanisms that comprise the activation of the proapoptotic proteins Bid and Bax and stimulation of cytochrome c release, all of which increase caspase 3, 8, and 9 activities in many cell types in commitment to chemical-induced apoptosis. 64 Hepatocyte apoptosis mediated by caspase 3 is a critical histologic feature in liver injury. 65 In the present study, caspase 3 staining showed that apoptotic hepatocytes increased during the process of liver injury. The liver and kidney caspase 3 activity in the CDDP-treated group was significantly higher than in the control group. These results indicated that Increased caspase 3 activity was an important contributing factor in the CDDP-mediated tissue damage. However, pretreatment with PSE significantly decreased CDDP-induced apoptosis in the liver and kidney tissues.
Hepatocellular degeneration and inflammatory cell infiltration in the portal area from liver and particularly tubular necrosis in the renal cortex from kidneys may have developed in CCDP-treated animals. 66 Yüce et al. 21 reported that structural changes in the kidney and liver sections of CDDP-treated groups may turn back by different agents. Our findings with the CCDP group was similar to those of the above researchers. The CDDP group had a dilatation in Bowman's space and in the collective tubules in the kidney sections. The tubules and liver cells from rats in the CDDP+PSE group had nearly normal histologic structure, except for a minor desquamation of the kidney and liver epithelial cells.
In conclusion, the present results demonstrate that CDDP-induced liver and kidney injury occurs after high-dose CDDP administration but that oral PSE treatment could diminish these side effects by way of its antioxidant and radical-scavenging effects. However, further studies are needed to determine the exact mechanism of PSE on CDDP-induced liver and kidney damage. PSE supplementation may have a potential therapeutic role for decreasing the side effects of liver and kidney toxicity resulting from administration of cancer medicines.
Footnotes
Author Disclosure Statement
No competing financial interests exist.