
Abstract
Cranberry extract has been reported as a therapeutic agent, mainly in urinary tract infections due to its anti-adhesive capacity. In order to compare the effects of proanthocyanidin (procyanidin) (PAC)-standardized cranberry extracts and commercial PAC A2, we first investigated the presence of genes encoding known adhesins on 13 strains of uropathogenic strains coming from patients with cystisis. After this characterization, the anti-adhesive effects of PAC A2 were assayed on selected uropathogenic Escherichia coli strains before testing cranberry extracts. Before checking inhibitory effect on bacterial adhesion to cells, we showed that neither PAC A2 or three cranberry extracts (A, B, and C) specifically inhibited the growth and did not supply any potential nutrient to E. coli strains, including the unrelated control strain. PAC A2 exhibited an inhibitory effect on the adhesion of two selected uropathogenic strains of E. coli. This work also showed that a preliminary exposure of bacteria to PAC A2 significantly reduced the adhesion. This phenomenon has been also observed with a lesser impact when uroepithelial cells were pretreated with PAC A2. Moreover, the assays were more robust when bacteria were in fast growing conditions (exponential phase): the adhesion to uroepithelial cells was greater. Significant reduction of adhesion to urepithelial cells was observed: around 80% of inhibition of adhesion with the cranberry extracts at equivalent PAC concentration of 50 μg/mL. The effects of the different assayed extracts were not obviously different except for extract B, which inhibited approximately 55% of adhesion at an equivalent PAC concentration of 5 μg/mL.
Introduction
S
In E. coli, four main types of adhesins have been identified that adhere to various receptors. Type 1 fimbriae bind to
Only the American cranberry has been studied for its biological properties for use in clinical practice (urology) to prevent or treat UTIs. Some studies have shown effects indicating the inhibition of bacterial adhesion to the urethra and bladder walls. 10,11 Other studies on human cohorts have shown a reduction in relapse rate associated with consumption of cranberry juice. 12 –14 Recently, the exact mechanisms of action of cranberry have been discovered: (1) inhibition of the synthesis of P fimbriae resulting, in the case of prolonged exposure, in complete disappearance of these adhesins; (2) elongation of the bacterial cell, 15 which prevents the adhesion of E. coli to the bladder wall; and (3) change in the bacterial cell surface property. 16
In 1989, Zafriri et al. 17 identified two compounds in cranberry that inhibit adhesins of E. coli: fructose and proanthocyanidins (procyanidins) (PACs), which contain A-type linkages in cranberry. The P fimbriae involved in pyelonephritis are inhibited only in the presence of cranberry or blueberry juice in a dose-dependent manner. 17 In vitro studies have shown that fructose inhibits the adhesion of type 1 fimbriae (mannose sensitive) by saturating the bacterial binding sites. 17,18 PACs are components produced by plants in response to environmental stresses and during bacterial infections. 19 One of the characteristics of PACs is their ability to bind to proteins. 20 An analysis by mass spectrometry has shown that purified cranberry PACs with a biological anti-adhesion activity have at least one type A double interflavanic linkage. 21
The aim of this study is to confirm the anti-adhesion effect of various PAC-standardized cranberry extracts. Preliminary, optimal conditions were determined using commercial PAC A2 in order to serve as reference. The anti-adhesion effects of PAC A2 and cranberry extracts have been evaluated on E. coli strains isolated from UTIs under different conditions concerning bacterial growth and preincubation of either uroepithelial cells or bacteria.
Materials and Methods
Chemicals and cranberry extracts
PAC A2 (reference number 0985S) was obtained from Extrasynthèse (Genay, France). The sample purity was verified by high-performance liquid chromatography (vide infra).
Three commercial cranberry powders were used, termed A, B, and C. Extract A was produced using purification steps adapted from the method described by Fuleki and Francis; 22 extract B corresponded to a mix between extract A and dried concentrated cranberry juice. Extract C was a dried cranberry juice concentrated extract. Stock solutions of cranberries were produced in dissolving powder in a water/ethanol (1:1 vol/vol) medium to an equivalent concentration of PACs equal to 500 μg/mL. The solutions were sterilized by filtration.
Bacterial strains
Thirteen uropathogenic strains of E. coli, numbered S1–S13, taken from patients with cystitis or pyelonephritis were provided by the Pontchaillou University Hospital in Rennes, France. The E. coli strain CSH120 (strain C) was used as a control for all procedures.
Uroepithelial cells
The T24 cell line of human bladder cancer was used in these experiments. The cells were cultured in an atmosphere of CO2–air (5:95 [vol/vol]; O2 partial pressure of 19.998 kPa) at 37°C. The medium used was RPMI 1640 medium containing 1% penicillin/streptomycin and 10% fetal calf serum. For all experiments, T24 cells were seeded at a density of 2×105 cells per well in a 24-well plate.
PAC assay and analysis
The PACs were completely hydrolyzed in a hydro-ethanolic acidified medium to release catechin and epicatechin monomers. The monomers were then oxidized by the action of the oxygen present in the medium. The oxidation product (cyanidin) was assayed using a spectrophotometer at 545 nm. This method is adapted from the European Pharmacopoeia method (Pharmacopoeia 6.0 01/2008:1220). The optical density (OD) at 545 nm corresponding to anthocyanins previously present in the extract was subtracted from results obtained after acid hydrolysis according the method of Bate-Smith. 23
In order to analyze the purity of the PAC A2 used in this study, high-performance liquid chromatography was performed. The chromatographic conditions used were an XTerra® (Waters Corp., Milford, MA, USA) C18 column (particle size, 3.5 μm; 3.0×150 mm) fitted with a precolumn of 20 mm filled with the same phase. The elution was done in gradient mode with the system water +0.01% trifluoroacetic acid/acetonitrile. The detection was done using a diode array spectrophotometer, and the profile was recorded at 280 nm.
Polymerase chain reaction detection of adhesin genes
To classify the strains according to the genes coding for the different adhesins, amplifications were performed using primer pairs targeting the genes afa, papC, papGII, and sfa. 24 The amplification reaction was performed using the GoTaq® (Promega, Madison, WI, USA) system according to the manufacturer's instructions with the following temperature cycles: 95°C for 5 minutes, 30 cycles of 30 seconds at 95°C, 30 seconds at the appropriate hybridization temperature, and 90 seconds at 72°C for polymerization. The polymerase chain reaction fragment was then analyzed by horizontal electrophoresis in 1% agarose gel prepared in 1×TAE buffer (40 mM Tris, 20 mM acetic acid, and 10 mM EDTA, pH 7.9). 25 After migration, the gel was immersed in ethidium bromide to detect the DNA band under ultraviolet light.
Growth and counting
The E. coli strains were transferred to Luria–Bertani (LB) solid medium 24 and then cultured in CFA medium, 26 which promotes the production of fimbriae and thus bacterial adhesion. The LB culture was grown at 37°C, and the CFA culture was grown at 35°C.
Growth was monitored by measuring OD at a wavelength of 570 nm (OD570nm), and counts were taken to establish a correlation between the OD570nm and the bacterial concentration (colony-forming units/mL) of all strains tested. Establishing this relationship is essential for testing adhesion of the E. coli strains to T24 bladder epithelial cells.
In order to test the nutrient ability of the different extracts of cranberry, E. coli strains were inoculated in M63 containing either various concentrations of cranberry extracts or glucose (1 mM) as the control, and the growth was monitored by measuring OD570nm.
Minimum inhibitory concentrations
Agar medium (Mueller–Hinton) was inoculated by flooding with a bacterial solution (OD570nm=0.01). Then filters (Durieux®, Torcy, France) pre-impregnated with solutions of cranberry (10 μL) and either undiluted (20 μg of PAC) or diluted 1:2 (10 μg of PAC), 1:4 (5 μg of PAC), or 1:8 (2.5 μg of PAC) were deposited on the agar. The plates were incubated at 37°C overnight.
Adhesion assays
The different adhesion tests with E. coli were performed with cultures in CFA medium. The strains of E. coli were grown in a volume of 100 mL at an initial OD570nm of 0.05. The culture was incubated at 37°C with stirring (140 rpm). For exponential-grown bacteria, the culture was monitored to 0.6 OD570nm corresponding to 3×108 colony-forming units/mL. For stationary-grown cells, the culture was monitored to 3.6 OD570nm corresponding to 9×109 colony-forming units/mL. Adequate dilutions were performed in fresh CFA medium to obtain bacterial solutions of 2×106 bacteria in a volume of 50 μL in order to inoculate one well containing T24 cells. The adhesion was performed for 2 hours, and three washing steps with phosphate-buffered saline solution were done in order to eliminate nonadhesive bacteria. The final step consisted of pouring 200 μL of phosphate-buffered saline containing 0.1% Triton X-100 in order to detach adhered bacteria. These bacterial solutions were enumerated on LB agar plates.
Different assays were performed: (1) no pretreatment of either T24 cells or bacteria (BnTCnT); (2) bacterial solutions (BTCnT); or (3) T24 cells were pretreated with either PAC A2 or cranberry solutions at different concentrations for 30 minutes (BnTCT). The results are expressed in percentages of adhesion corresponding to 100×[number of bacteria (colony-forming units) adhered to 2×105 uroepithelial cells/total bacteria added]. The relative inhibition (%) corresponds to 100×(1 – [number of adhered bacteria at a given PAC concentration/number of adhered bacteria when there was no PAC]). The data are presented as mean±SD values, and significance level was set at P<.05. Independent Student's t tests were used to determine if there were significant differences. The adhesion experiments conducted in the presence of PAC A2 or cranberry extracts are recorded in equivalent PAC concentration in Results.
Results
Characterization of the E. coli strains
According to the results of the polymerase chain reactions (data not shown), five groups of strains could be determined based on combinations of adhesins: strains S4, S9, S10, and S11 did not possess genes coding for the desired adhesins; strains S1, S7, S12, and S13 had the gene coding for the adhesin Afa; strain S5 had the gene coding for PapGII; strains S2, S3, and S8 had the genes coding for PapC, PapGII, and Afa; and, finally, strain S6 had the genes coding for PapC, PapGII, Sfa, and Afa.
For the following experiments, strains S2 and S5 and the control strain C have been chosen because Pap adhesins are most frequently encountered in cases of cystitis and/or pyelonephritis and Afa adhesins are responsible for urinary tract problems in only 10% of cases.
Analysis of minimum inhibitory concentrations and nutrient ability of cranberry extracts
The high-performance liquid chromatography profile in Figure 1 shows that the PAC A2 used in this study is pure. The three samples of cranberry (A, B, and C) were analyzed, and their respective PAC concentrations are 512 mg/g, 282 mg/g, and 29 mg/g, respectively.
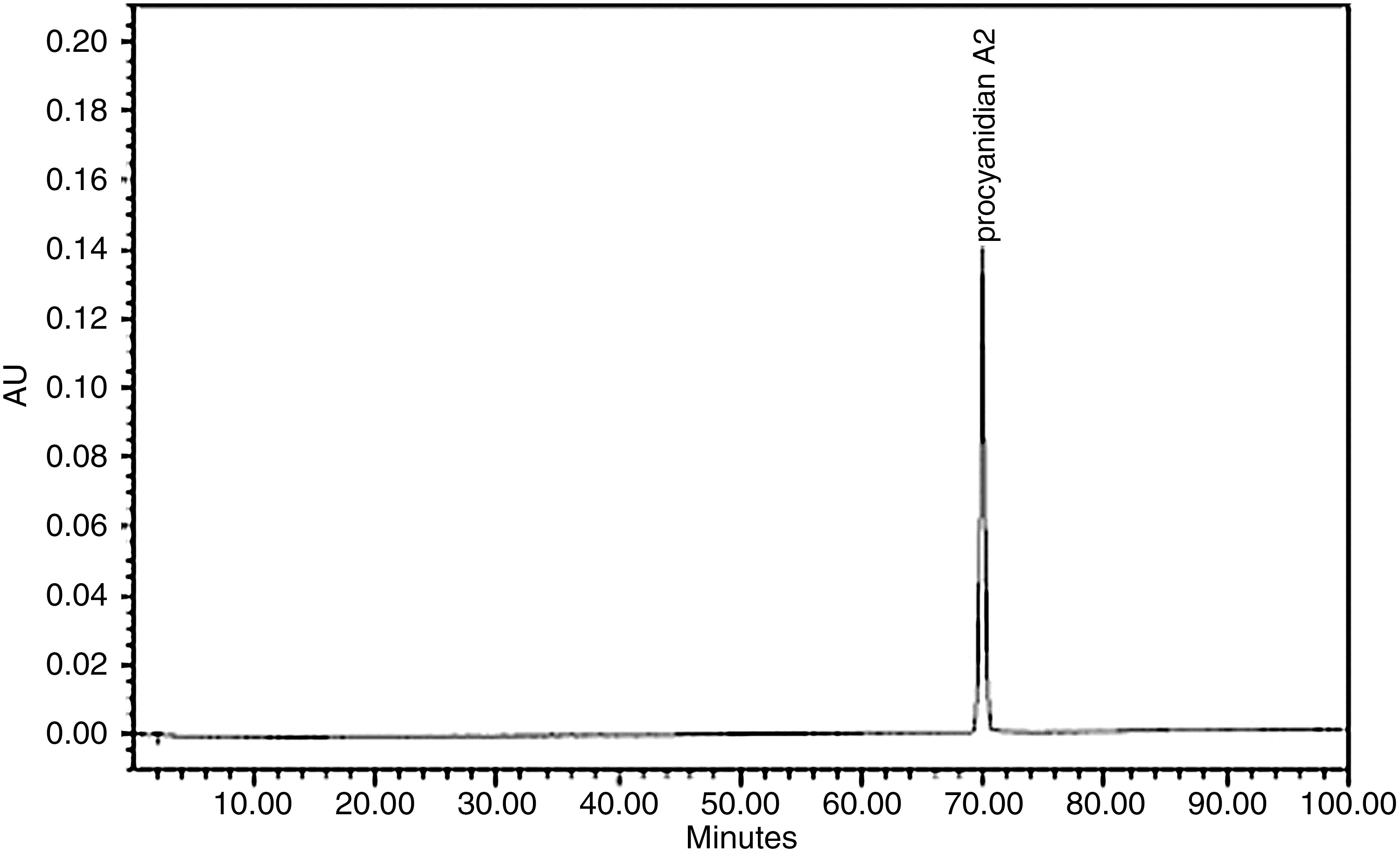
High-performance liquid chromatography profile of procyanidin A2. Conditions are given in Materials and Methods. AU, arbitrary units.
The minimum inhibitory concentration has been determined to ensure that none of the products contained in the cranberry samples inhibits the growth of bacteria. Extracts of cranberry, whatever the concentrations, did not inhibit the growth of bacteria (data not shown). In some cases better growth has been observed around the filters, which could be explained by the presence of compounds metabolized by bacteria in the samples. The assays concerning the nutrient abilities of either PAC A2 or cranberry extracts have shown that the E. coli strains did not grow in minimal medium supplemented with either PAC A2 or cranberry extracts compared with the assays performed with glucose (data not shown). These results have shown that the cranberry extracts were not nutritious.
Adhesion of E. coli strains to uroepithelial cells in the presence of PAC A2
The objectives of this section were (1) to determine the dose effect associated with the anti-adhesion properties of PAC A2 in the bacterial adhesion to T24 bladder cancer cells to compare the results with those observed with extracts of cranberry, (2) to analyze any differences between strains S2 and S5, (3) to consider the physiological state of bacteria (exponential phase or stationary phase of growth), and (4) to determine the effect of prior contact with PACs on bacterial adhesion to uroepithelial cells.
The anti-adhesion tests of PAC A2 were performed on three strains of E. coli: strain C, control strain; strain S2, carrier of the genes coding for PapC, PapGII, and Afa fimbriae; and strain S5, carrier of the genes coding for Pap GII fimbriae.
The results for all the strains in stationary phase and exponential phase are presented in Tables 1 and 2.
Different assays were performed: (1) no pretreatment of either T24 cells or bacteria (BnTCnT); (2) bacterial solutions (BTCnT); or (3) T24 cells were pretreated with either proanthocyanidin (PAC) A2 or cranberry solutions at different concentrations for 30 minutes (BnTCT). All experiments were performed in triplicate. The % of adhesion was defined as 100×(adhered colony-forming units/added colony-forming units in well).
Significant difference between adhered bacteria at a given PAC concentration and adhered bacteria without PAC with the significance level set at P<.05.
All experiments were performed in triplicate. The % of adhesion was defined as 100×(adhered colony-forming units/added colony-forming units in well).
Significant difference between adhered bacteria at a given PAC concentration and adhered bacteria without PAC with the significance level set at P<.05.
In the stationary phase (Table 1), strains S2 and S5 carrying fimbriae adhered in larger quantities (by a factor of 2–3): the percentage of adhesion for strains S2 and S5 were, respectively, 6.13±1.41% and 7.31±0.66% compared with 2.64±0.93% of control strain C. The presence of PAC A2 in the medium without pretreatment induced a 28.2–63.3% decrease in the quantity of adhesive bacteria (S2 and S5) starting at 5 μg/mL. There was a decrease in adhesion of the control strain of up to 45.5%. When bacteria were pretreated with PAC A2 (BTCnT), the inhibition of adhesion was enhanced—from 37.6% to 77.1%—with a superior effect on strain S5. For the control strain, the inhibition was also enhanced, up to 78.1%.
The pretreatment of uroepithelial T24 cells (BnTCT) also favored the inhibition of adhesion compared with the assay with no pretreatment. For strain S2 the maximum inhibition reached was about 52.0%, and for strain S5, 75.5% of inhibition of adhesion was obtained with 100 μg/mL PAC A2. In the case of control strain C, the inhibition of adhesion was only observed when PAC A2 was at higher concentrations (50 and 100 μg/mL). This control strain adhesion inhibition was also observed when the bacterial or uroepithelial cells were pretreated (BTCnT or BnTCT, respectively, yielding 41.9–77.1% and 0–41.9% inhibition). However, the inhibition is not dose dependent.
With or without pretreatment of bacteria or uroepithelial cells, the reduction in adhesion of the adhesive test bacteria can approach nearly 75%. However, according to the results obtained with the control strain, it is difficult to state conclusively that PAC A2 has an anti-adhesion effect when the bacteria are in stationary phase.
When the uropathogenic E. coli bacteria are in exponential phase (Table 2), the quantity of bacteria adhering to uroepithelial cells is five to eight times greater than in stationary phase. Moreover, strains S2 and S5 carrying fimbriae adhered six to eight times more in quantity than control strain C under the experimental conditions. In the absence of pretreatment (BnTCnT), the adhesion inhibition of strains S2 and S5 was, respectively, 71% and 77.8% at a concentration of 100 μg/mL PAC A2. However, the nonspecific inhibition observed for the control strain was >57.3%.
When uroepithelial cells were pretreated (BnTCT), adhesion inhibition at a concentration of 100 μg/mL was 90% for strain S2 and 76.6% for strain S5. The nonspecific inhibition observed for control strain C was 67.8%. It is therefore very difficult to make comparisons and draw conclusions on the specific efficacy of PAC A2.
When the bacteria were pretreated (BTCnT), the adhesion inhibition of strains S2 and S5 was, respectively, 93.5% at a PAC concentration of 50 μg/mL and 94.8% at 20 μg/mL, whereas the nonspecific inhibition observed for control strain C was between 18.8% and 44.9%. The anti-adhesion effect observed for strain S5 (93%) started at 10 μg/mL PAC A2, and it remained stable up to 100 μg/mL. When the pretreatment was applied to bacteria, nonspecific inhibition was limited. Compared with the control strain, the adhesion of strains S2 and S5 was inhibited starting at a concentration of 10 μg/mL PAC A2 (44.9%, 70.7%, and 93%, respectively, inhibition of adhesion). This effect appears to be dose dependent, with maximum inhibition at 93.5% and 94.8% for strains S2 and S5, respectively.
These results support the conclusion that PAC A2 has an inhibitory effect on adhesion of E. coli bacteria to T24 uroepithelial cells. This inhibitory effect is specific for uropathogenic strains carrying P fimbriae when the bacteria are in the exponential phase of growth. The exponential phase is therefore appropriate for demonstrating the anti-adhesion effect of PACs.
In addition, treating the bacteria with PAC A2 prior to contact with uroepithelial cells induced greater inhibition on bacteria that carry the genes coding for the PapGII adhesin, such as strain S5, in the exponential growth phase. A concentration of 50 μg/mL PACs appears to be appropriate for demonstrating the anti-adhesion effect.
The S5 strain, which carries a single type of PapGII fimbriae, was taken as the model strain for the subsequent experiments.
Adhesion of strains of E. coli to uroepithelial cells in the presence of PAC-standardized cranberry extract
The objectives of this section were (1) to demonstrate, in the experimental model previously defined, the anti-adhesion action of cranberry extracts enriched with PACs, (2) to compare the results with those described previously from experiments with PAC A2 to assess the consistency between the observed effects and the PAC content in these extracts, and (3) to compare the anti-adhesion action of cranberry extract enriched with varying amounts of PACs, at equivalent final PAC concentration.
Table 3 presents the results obtained with the model strain S5, which carries the PapGII fimbriae, in exponential growth phase, placed in contact with the cranberry extracts A, B, and C, at PAC equivalent concentrations of 0–180 μg/mL. The bacteria were pretreated as described previously. The experiments of this study were also performed in triplicate.
All experiments were performed in triplicate. The % of adhesion was defined as 100×(adhered colony-forming units/added colony-forming units in well).
Significant difference between adhered bacteria at a given PAC concentration and adhered bacteria without PAC with the significance level set at P<.05.
Strain C was used as the negative control because it has no adhesion fimbriae. Under these experimental conditions, we did not observe any nonspecific inhibition by cranberry extracts on adhesion of the control strain C.
In the model strain S5, the inhibitions of adhesion approached 80% at both equivalent PAC concentrations of 20 μg/mL and 50 μg/mL. Extracts A and B have already inhibited adhesion at equivalent PAC concentration of 5 μg/mL. Global analysis of the relative inhibitions of adhesion by the different extracts has shown that the difference between extracts A and B was not significant (P=.191), but the differences between extracts A and B versus C were significant (A vs. C, P=.001; B vs. C, P=.015).
Discussion
Adhesion is a major virulence factor in the complex pathogenesis of UTIs. In this work, a genotypic approach has been used to detect and identify the presence of adhesin-encoding operons. Of the 13 strains obtained from patients with cystisis, which have been randomly chosen in this work, four did not carry any gene encoding adhesin, only one was positive for the sfa operon, 12 of them were afa +, five possessed the allele papGII, and four possessed the allele papC. Pap adhesins are most frequently encountered in cases of cystitis and/or pyelonephritis: 74.7% of strains causing pyelonephritis and 44.1% of strains associated with cystitis. Afa adhesins are responsible for urinary tract problems in only 10% of cases. 23 For these reasons, the experiments on the inhibition of adhesion were performed on strains S2 and S5. Previous analyses were performed to be sure that either PAC or cranberry extracts did not inhibit or provide any supplementary nutrient. The results are consistent with the literature indicating that the mode of action of cranberry is not bactericidal.
The anti-adhesion tests of PAC A2 were performed on strains that were in either stationary or exponential phase. The adhesion conditions were optimal when the bacteria are in exponential growth phase (five times more bacteria adhered on the same uroepithelial cell matrix), facilitating the identification of the anti-adhesion effects. These results could be related to the intricate cross-talk between different adhesin gene clusters and their relative expression during different stages of growth or when subjected to stressful conditions. 27
However, our results support the conclusion that PAC A2 has an inhibitory effect on the adhesion of E. coli carrying adhesin genes to bladder cells. The application of PAC A2 reduced up to around 95% of adhesion of strain S2 and S5. Notable effects were also observed at relatively low concentrations of PAC A2 (5 μg/mL). These observations are in agreement with the results of Pinzon-Arango et al., 28 who demonstrated that PAC treatment resulted in a decrease of attachment to uroepithelial cells of an E. coli P-fimbriated variant. The differences with our study consist of the assayed E. coli strains and in the relatively high level of PAC that inhibited adhesion to eukaryotic cells. The results obtained with control strain C shown that there was some difficulty in making comparisons and drawing conclusions on the specific efficacy of PAC A2. Nevertheless, the variations were consistent with variations due to differences in concentration of bacteria that have adhered to uroepithelial cells, especially for strain C in either exponential or stationary growth phase. Indeed, there was probably a nonspecific inhibition due to the natural interaction occurring between the PACs present in the medium and the surface proteins of the bacteria and uroepithelial cells. However, the anti-adhesion effect is greater with the test strains than with the control strain, in terms of the large quantity of bacteria that have not adhered compared with the results in the absence of PAC A2.
The experimental model developed with PAC A2 highlighted the in vitro inhibitory effect of cranberry extracts on adhesion of clinically isolated uropathogenic E. coli bacteria that carry PapGII fimbriae. Whatever the concentration of PACs in the tested extracts (512 mg/g PACs for extract A, 282 mg/g for extract B, and 29 mg/g PACs for extract C), the effects were quite similar when the final equivalent PAC concentrations were equivalent in the experimental setting. This confirms that the PACs are responsible for the observed inhibitory effects. Cranberry extracts have also been shown to alter the bacterial surface properties such as fimbriae and lipopolysaccharides. 27 Moreover, the anti-adhesion effectiveness of cranberry extracts tested at constant PAC concentrations is quite comparable to the anti-adhesion activity of PAC A2: 70–80% inhibition of the adhesion of uropathogenic E. coli bacteria was observed using concentrations of 20 and 50 μg/mL PACs.
Many publications have suggested that the anti-adhesion effect of PACs from cranberry is related to the presence of specific linkages, known as type A bonds. 21 Recently, another role was attributed to cranberry extracts and more precisely to the nondialyzable material, which contained 65.1% PACs, that has been shown to interfere in bacterial cell–cell signaling systems. 29 PACs are polymers composed of flavanol subunits (catechin and epicatechin), which can be linked by two types of bonds: type B linkages, for which the two monomers are mono-covalently linked between carbons C4 and C8, and type A linkages, involving a second interflavanic bond between the carbon C2 and the oxygen of carbon C7 (Fig. 2).
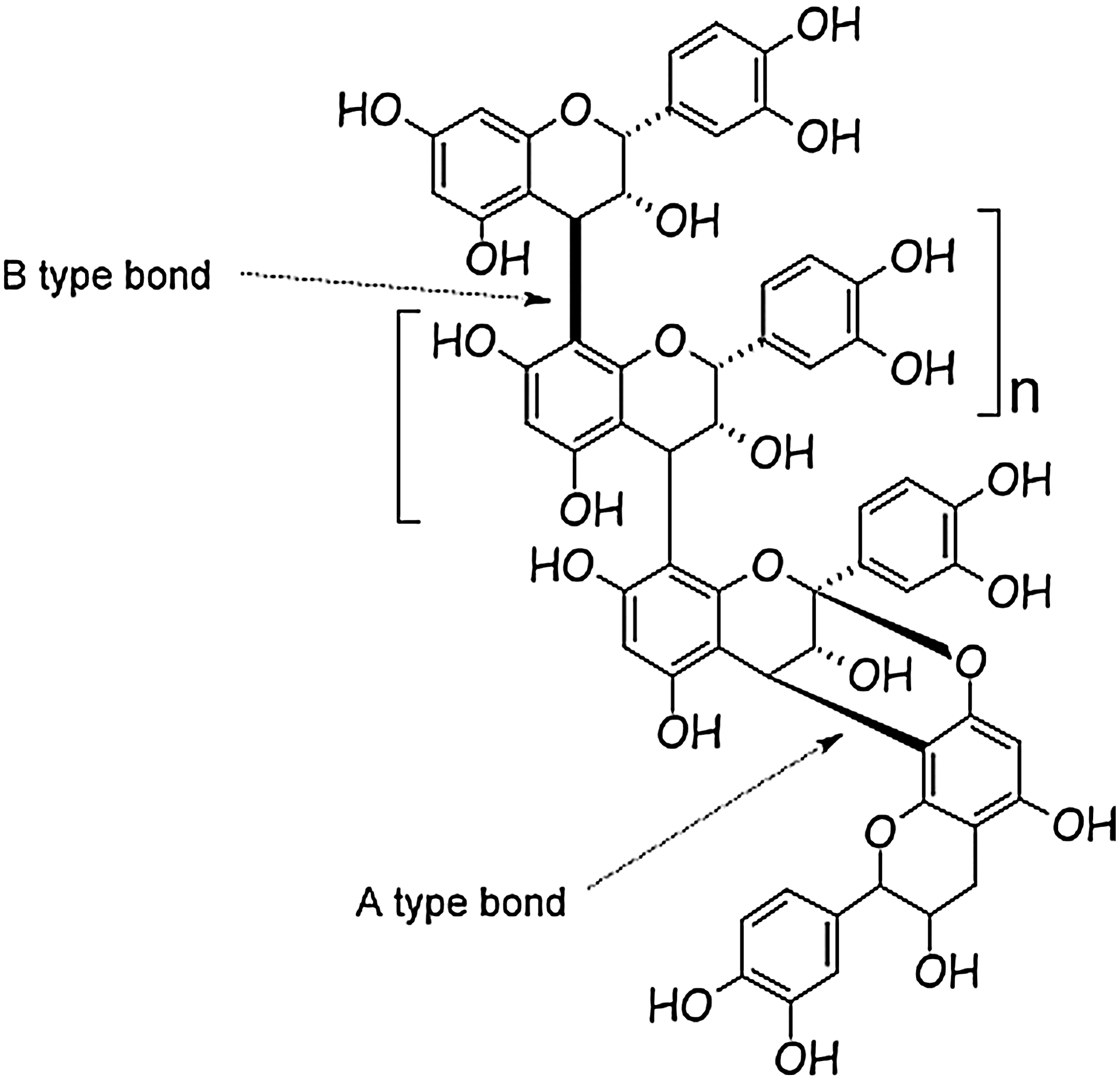
Structure of proanthocyanidins showing type A and type B interflavanic bonds.
Thioacidolysis of the polymers performed on the different cranberry extracts showed that type A bonds represent around 30% of total interflavanic bonds present in PACs, compared with about 70% of type B bonds. During thioacidolysis, the type B linkages are hydrolyzed, whereas the type A linkages are not cleaved, and flavanols linked by these bonds are present in the medium after acid hydrolysis. Acid depolymerization can also determine the mean degree of polymerization of PACs. The purified extracts of cranberry showed a mean degree of polymerization of six to 13 units.
After thioacidolysis the solution contains only monomers and type A dimers. There are no trimers or other polymers in the medium. To our knowledge there is no example of exclusively type A trimer or polymer in the literature. This clearly indicates that there is no succession of several A linkages along the polymer chains of PACs.
Accordingly, cranberry PACs are composed of a succession of subunits linked by both A and B bonds, never with two or more successive A bonds.
Studies have demonstrated the absence of an anti-adhesion effect for PACs from other plant sources containing no interflavanic A linkage. 30 These observations, in conjunction with the analytical results obtained on the structure of PACs and the biological effects described in this study, demonstrate that the presence of a single A-type linkage in a polymer is necessary and sufficient to confer anti-adhesion activity versus uropathogenic E. coli bacteria. These results must be confirmed from a biological perspective by further studies on these cranberry extracts, including conducting a human clinical trial to confirm the results obtained in this study in vivo.
Footnotes
Acknowledgments
We thank Christine Baysse for helpful comments on the manuscript. A part of this work was financially supported by Europe–Feder. This work was supported by a grant from the research division of Diana Naturals. UMR CNRS 6026, University of Rennes 1 contributed to the experimental work and interpretation of the data.
Author Disclosure Statement
All authors declare competing financial interests. C.I. and M.B. are employees of Diana Naturals. The research was funded by Diana Naturals, and they have participated in the chemical characterization of the extracts, the establishment of the study design, the analysis of results, and writing of the manuscript. G.E. and S.G. of the University of Rennes 1 have collected the E. coli strains, have acquired the uroepithelial cells, have performed the biological experiments, and have participated to the establishment of the study design, the analysis of results, and the writing of the manuscript. All authors read and contributed to the finalization of the manuscript.