
Abstract
Mushrooms are useful in balancing human diet and treating different health problems. The main determinant of the fluidity of erythrocyte membranes and rheologic properties of blood is the lipid composition of the membrane, which directly depends on lipids present in the diet. Lipid profiles of 4 European wild edible mushrooms, as well as the ability of mushroom lipids to modify the fluidity of erythrocyte membrane, were examined by using gas chromatography–mass spectometry, gas chromatography–flame ionization detector, and electron paramagnetic resonance spin probing technique. Lipids from 2 species—Macrolepiota procera and Collybia platyphylla—provoked an increase in erythrocyte membrane fluidity. Therefore, introduction of these and other wild mushrooms with similar lipid profiles to the human diet could be very beneficial in the treatment of hypertension and other cardiovascular diseases related to decreased fluidity of erythrocyte membranes.
Introduction
M
Previous studies have shown some positive effects of dietary intake of mushrooms on cardiac diseases, the main cause of death in most western societies. 16 However, the association between diet and the fatty acid profile of erythrocyte membranes 17 –20 opens a possibility for further application of mushrooms in clinical nutrition. The lipid composition regulates the physicochemical properties of cellular membranes, such as fluidity and viscosity. 21 –24 Decreased fluidity of erythrocyte membranes affects their deformability 25,26 and flow properties of the blood. 27,28 Earlier studies found that membrane fluidity is significantly lower in spontaneously hypertensive rats and patients with essential hypertension compared with values in normotensive controls. 29,30 Thus, it has been proposed that decreased fluidity of erythrocyte membranes might contribute to the pathophysiology of hypertension and other cardiovascular diseases. 31 –34
Researchers have reported that cell membrane alteration is a source of primary hypertension. 35 Therefore, a diet that can increase fluidity of erythrocyte membranes could be a useful part of hypertension treatment. 36 Selected mushrooms could match demands of an antihypertensive diet, as shown previously for 3 fungal species—Ganoderma lucidum, 37 Grifola frondosa, 38 and Tricholoma mongolicum 39 —that have been reported to reduce arterial blood pressure. In addition, other mushroom species (Agricus bisporus and Volvariella volvacea) showed hypocholesterolemic effects. 40,41 In the present study we evaluated the effects of lipid extracts from 4 wild mushroom species on fluidity of erythrocytes and liposomes by using an electron paramagnetic resonance (EPR) technique with 3 different lipophilic spin probes.
Materials and Methods
Sample preparation
Mushroom samples were collected from the Istra region of Croatia in the late summer of 2008. Macrofungi were identified and classified. Mushrooms fruiting bodies were cleaned up to remove any residual compost. Fresh mushrooms were air-dried and then stored in air-tight plastic bags at room temperature. All dried mushrooms samples were ground in a blender before the extraction. The extraction process was carried out at a laboratory-scale high-pressure extraction plant (HPEP, NOVA-Swiss, Effertikon, Switzerland), as described elsewhere. 42 The main plant parts and properties, by manufacturer specification, were as follows: diaphragm-type compressor (with pressure range up to 1,000 bar), extractor with internal volume of 200 mL (pmax=700 bar), separator (with internal volume of 200 mL, pmax=250 bar), and maximum CO2 mass flow rate of 5.7 kg/h. Ground mushroom samples were placed into an extractor vessel to fill up the vessel. The extraction process was carried out for 4 hours. The flow rate of CO2, expressed under normal conditions, was 97.725 dm3/h. Extraction conditions were pressure of 300 bar and temperature of 40°C. Separator conditions were 15 bar and 23°C. After extraction, obtained extracts were placed into glass bottles; the bottles were sealed and stored at −4°C to prevent degradation.
Chemicals
Commercial CO2 (Messer, Novi Sad, Serbia) was used. Nitroxide spin probes 7DS (2-(5-carboxypentyl)-2-undecyl-4,4-dimethyloxazolidine-3-oxyl, 5DS 2-(3-carboxypentyl)-2-tridecyl-4,4-dimethyloxazolidine-3-oxyl, and 10DS 2-(8-carboxyoctyl)-2-octyl-4,4-dimethyloxazolidine-3-oxyl were purchased from Molecular Probes (Junction City, OR, USA). L-α-phosphatidylcholine was obtained from Sigma-Aldrich (St. Louis, MO, USA). All other chemicals were of analytical reagent grade.
Analytical methods
GC-MS
For GC-MS analysis, the samples (200 mg) were extracted with acetonitrile (1 mL) for 15 minutes in an ultrasonic bath. The acetonitrile extracts were filtered over a 50μ filter and injected into the GC-MS system. GC-MS analysis was run on an Agilent 7890A GC system coupled to a quadrupole mass spectrometer model Agilent 5975C. For mushroom extracts analysis, the GC was fitted with a capillary HP-5MS column with a 0.25-μm film thickness, 30 m length, and inner diameter of 0.25 mm. The operating conditions were as follows: injector temperature, 250°C; split ratio, 20:1; detector temperature, 300°C; carrier gas, helium; and constant pressure, 21.956 psi. The temperature program was 60°C–300°C (3°C/min). The MS conditions were as follows: ionization voltage, 70 eV; ion source temperature, 230°C; and scan range, m/z 35–550. Volumes of 2 μL of acetonitrile extracts were injected. The identification of the individual components was based on computer matching with the Adams and National Institute of Standards and Technology/Environmental Protection Agency/National Institutes of Health version 2.0d mass spectral libraries. The constituents of the extracts were also identified by comparing their retention times to those ini the Adams library.
For methyl esters analysis, the GC was fitted with a capillary DB-23 column with a 0.25-μm film thickness, 30 m length, and 0.25-mm inner diameter. The operating conditions were as follows: injector temperature, 250°C; split ratio, 20:1; detector temperature, 300°C; carrier gas, helium; constant pressure, 37.7 psi. The temperature program was from 200°C to 240°C (5.6°C/min), and held at 240°C for 15 minutes. The MS conditions were the same as those for intact extract analysis.
GC/FID
For GC-FID analysis, the mushroom extracts were transesterified. The extracts (200 mg) were dissolved in 2 mL of 3% H2SO4 dissolved in methanol. The vials that contained the reaction mixtures were sealed and heated in a water bath for 2 hours. After cooling, the mixture was neutralized with an NaHCO3 solution (2 mL), and 2 mL of CH2Cl2 was added. The organic layer was dried-over anhydrous Na2SO4, filtered and evaporated on reduced pressure. The residue was dissolved in CH2Cl2 and injected into the GC system. For GC/FID analysis of methyl esters, a Hewlett-Packard chromatograph model HP 5890 Series II equipped with an FID and fitted with the same column as those for GC-MS analysis (DB-23). The operating conditions were as follows: injector temperature, 250°C; split ratio, 30:1; detector temperature, 300°C; and carrier gas, H2 (flow rate, 1.0 mL/min). The temperature program was as follows: from 150°C to 240°C (4°C/min) and held at 240°C for 10 minutes. A sample (1 μL) dissolved in CH2Cl2 (1:100 v/v) was injected. The percentage composition was computed by the normalization method from the GC-FID peak areas.
EPR spin probing spectroscopy
Liposome fluidity
Effects of the extracts on fluidity were explored by using liposomes as a membrane model. To prepare the liposomes, a chloroform solution of L-α-phosphatidylcholine was evaporated under vacuum to dryness. Then, phosphate buffer (Na2HPO4, 1.2 g/L; NaH2PO4, 0.43 g/L; pH, 7.4) was added to obtain a lipid concentration of 100 mM. Mushroom extracts were dissolved in liposome solution to obtain a final concentration of 1 mg/mL, and the suspension was vortexed. In a control sample, an aliquot of buffer was added. Liposomes solution was applied to dried 7DS (previously dissolved in ethanol) and vortexed. 43 Optimal (spin probe/membrane lipid) ratio of approximately 1:200 was used. 36
Erythrocyte membrane fluidity
Two mushroom extracts that provoked changes in the fluidity of liposomes were studied in further experiments on erythrocytes. Fresh blood was obtained from 4 healthy volunteers age 25–35 years by using tubes containing 0.072 mL of 7.5% K3EDTA as the anticoagulant per 3 mL of blood (Vacuette EDTA, Greiner Bio-One, Austria). Spin-labeling of the erythrocytes (erythrocyte membranes) was performed as described elsewhere. 44 In our experience, it is very important to precisely follow the procedure in order to obtain reproducible results. Fresh blood was washed 3 times with a isotonic phosphate buffer solution (NaCl, 8.8 g/L; Na2HPO4, 1.2 g/L; NaH2PO4, 0.43 g/L; pH was adjusted to 7.4 using 1 M HCl) by centrifugation at 3500 g per 10 minutes at 4°C. The hematocrit in fresh blood was 40%, and the same hematocrit was adjusted in all samples before incubation. Extracts were dissolved in methanol, and those solutions were diluted with buffer to obtain concentration of extracts of 1 mg/mL; the solutions were then left for 2 hours until methanol evaporated. The solutions were added to erythrocytes to obtained final concentration of extracts of 0.2 mg/mL. Ethanol solutions of fatty acid spin probes 5DS and 10DS were supplemented on the walls of tubes. After the ethanol evaporated, sample was added and gently mixed. The amount of doxyl stearates was added to the erythrocytes to obtain the optimal spin-label/membrane-lipid ratio of approximately 1:100. 45
EPR measurements
EPR spectra were recorded by using a Varian E104-A EPR spectrometer operating at X-band (9.1 GHz) and adjusted to the following settings: modulation amplitude, 2 G; modulation frequency, 100 kHz; microwave power, 10 mW; scan range, 100 G; scan time, 4 minutes; and time constant, 0.25 second. The temperature was controlled at 20°C during the measurements. Spectra were recorded and analyzed by using EW software (Scientific Software). The order parameters (S) were calculated as shown elsewhere 46 and used as an indication of membrane fluidity.
Statistical analysis
The data are presented as means±standard deviation of at least 3 separate experiments. Significance of fluidity change provoked by an extract relative to controls was calculated by using the Student t-test. Means were considered significantly different at a P value less than .05. Correlation coefficients determining the relationship between S and the content of each particular fatty acid, as well as ratios of unsaturated to saturated fatty acids (UFA/SFA) and polyunsaturated to monounsaturated fatty acids (PUFA/MUFA), were calculated by means of a Pearson correlation test using Statistica 6.0 (StatSoft Inc, Tulsa, OK, USA). Statistically significant correlations were defined at a P value of .05 or less.
Results and Discussion
Lipid profiles of mushrooms
Table 1 shows results of GC-MS analysis of minor lipid compounds extractable to acetonitrile, present in supercritical mushroom extracts. Results are presented as percentage of total area of the chromatogram (not shown). Predominant “trace” lipid components (up to 5% of total lipid content) of mushroom extracts are free fatty acids and steroids. Triglycerides are not extractable to acetonitrile.
GC-FID was used for quantitative evaluation of fatty acid composition of triglycerides in the extracts. Transesterification was performed before the analysis. Table 2 presents the results. The UFA/SFA and PUFA/MUFA ratios in studied mushrooms were as follows: C. platyphylla, 2.87 and 0.88; L. deterrimus, 0.88 and 0.95; M. procera, 2.55 and 3.20; and A. mellea, 4.09 and 0.99. Among saturated acids, palmitic acid predominated (16:0); the exception was L. deterrimus, in which stearic acid (18:0) was the main component. M. procera exhibited specific fatty acid composition with high content of linoleic (18:2 c9c12) and palmitic acid and low content of oleic acid (18:1 c9). Fatty acid composition of C. platyphylla showed similar levels of linoleic, oleic, and palmitic acids. A. mellea showed similar percentages of oleic and linoleic acid, with a lower level of palmitic acid. Fatty acid composition of M. procera harvested in Asia was described in recent reports. 7,8 Fatty acid profiles of Asian M. procera showed higher UFA/SFA and PUFA/MUFA ratios (3.95 and 3.64) 8 than our results.
Trans-9-octadecanoic acid methyl ester.
Exact structure of the isomer could not be determined.
Cis,cis-9,12-octadecadienoic acid methyl ester.
10,13-Octadecadiynoic acid methyl ester.
MUFA, monounsaturated fatty acid; PUFA, polyunsaturated fatty acid; SFA, saturated fatty acid; UFA, unsaturated fatty acid.
Effects on liposome fluidity
Liposomes were used as a model system for cellular membrane. 47 Doxyl stearates intercalate into the membrane, and specific measures of their EPR spectra depend on membrane fluidity. 46 To obtain data on fluidity of the middle layer of lipid semileaflet of liposomes, 7DS (which has the nitroxide), we used the EPR-active moiety on carbon 7 from the carboxyl group. Figure 1a shows EPR signal characteristic for 7DS in liposome membrane, as well as the method of calculation of S, which is reciprocally proportional to fluidity. 44 Table 3 shows S obtained in liposomes after addition of mushroom extracts. Extracts of some species provoked an increase in liposome fluidity; the most pronounced effects were observed for M. procera. Lipid fractions of 2 species—L. deterrimus and A. mellea—did not provoke statistically significant changes. These 2 species were excluded from the set of experiments performed on erythrocytes.
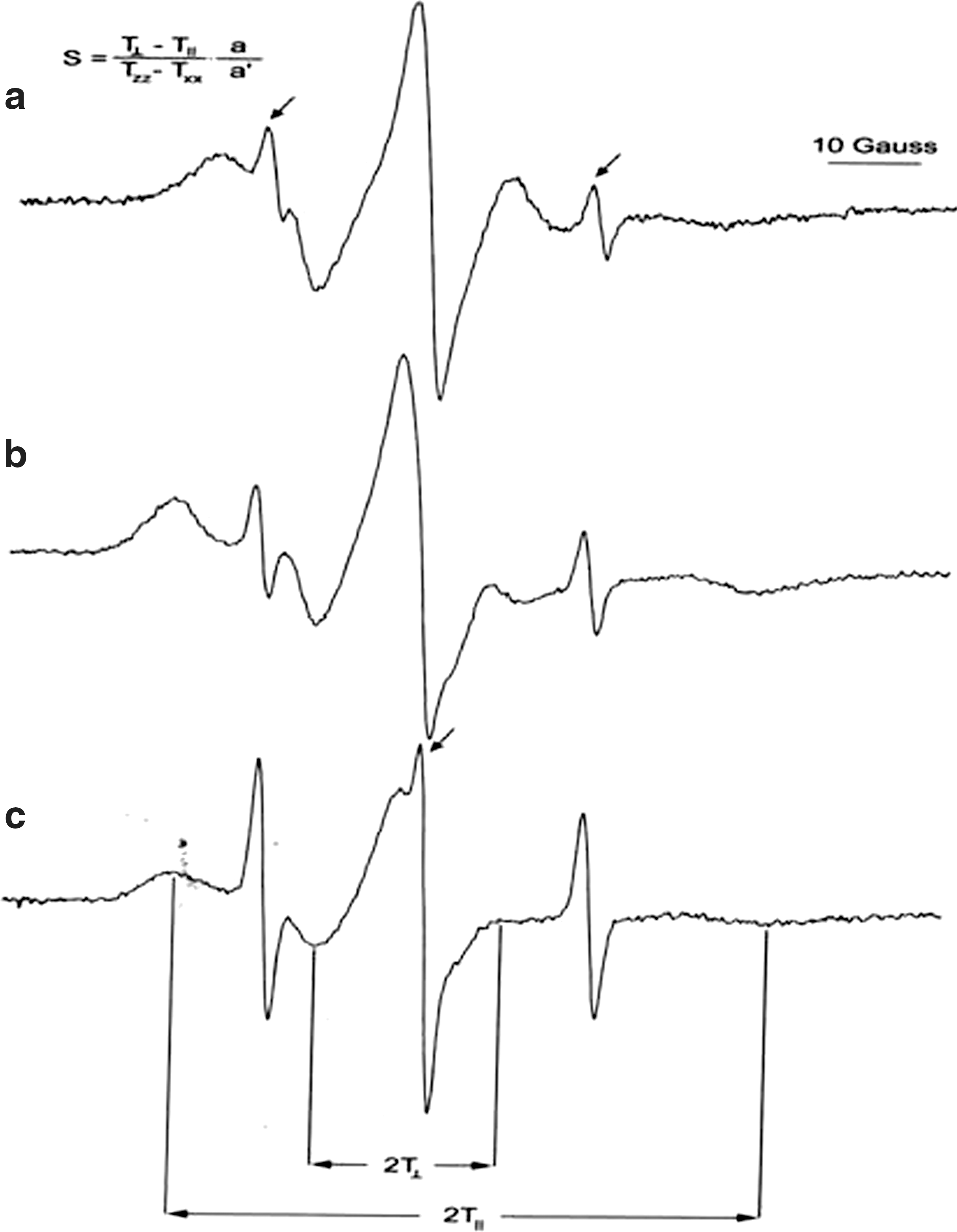
Characteristic electron paramagnetic resonance spectra of (
Data expressed with plus/minus sign are the mean±standard deviation.
Statistical significance relative to control.
Untreated liposomes.
Statisticaly nonsignificant relative to control.
NS, not significant; S, order parameter.
Effects on erythrocyte membrane fluidity
Two spin probes—5DS and 10DS, with the EPR-active moiety on carbons 5 and 10 from the carboxyl group—were used to evaluate S near the membrane surface (5DS) and in deeper membrane layers (10DS). Figure 1b and 1c shows characteristic EPR spectra of erythrocytes labeled with 5DS and 10DS. Table 4 presents S (calculated as illustrated in Fig. 1) of membrane of erythrocytes exposed to extracts of 2 mushroom species.
Data expressed with plus/minus sign are the mean±standard deviation.
Statistical significance relative to control.
Untreated liposomes.
Statisticaly nonsignificant relative to control.
5DS, 2-(3-carboxypentyl)-2-tridecyl-4,4-dimethyloxazolidine-3-oxyl; 10DS, 2-(8-carboxyoctyl)-2-octyl-4,4-dimethyloxazolidine-3-oxyl; NS, not significant; S, order parameter.
As in liposomes, lipids from M. procera provoked the most pronounced increase in fluidity of erythrocyte membrane. These effects were statistically significant for fluidity of both studied membrane layers. Lipids from C. platyphylla had halfway effects, provoking increase in fluidity only in deeper membrane layers. M. procera possesses relatively high antioxidative capacity 48 and thus could prevent a decrease in membrane fluidity related to lipid peroxidation. 49
On the basis of fatty acid composition of 2 mushroom species with positive effects on fluidity, we propose that linoleic acid could be responsible for the observed increase in fluidity of membrane layers close to the surface and that palmitic acid seems to modify mechanical properties of deeper layers.
Linoleic acid could increase membrane fluidity by interrupting the orderly structure of the membrane composed mostly of UFA. 50 Palmitic acid (16:0) should increase mobility of the ends of constituent fatty acids of erythrocyte membrane, which have longer carbon chains (18:0). Therefore, palmitic acid could disturb the organization of inner layers of the membrane, thus provoking increase of fluidity. Pertinent to this, it has been reported that an increased level of palmitic acid improves rheologic properties of erythrocytes. 51 To substantiate proposed mechanisms, we performed a correlation analysis of data obtained on fatty acid profiles of studied mushrooms and their effects on membrane fluidity. A significant positive correlation (r>0.9) was found between erythrocyte membrane fluidity at the surface (determined by using 5DS) and the level of linoleic acid and PUFA/MUFA ratio, as well as between the fluidity of deeper membrane layers (determined by using 10DS) and relative level of palmitic acid. Our results indicate that the PUFA/MUFA ratio of dietary lipids is more relevant for the effects on erythrocyte fluidity than is the UFA/SFA ratio. High PUFA/MUFA ratio also suggests that mushrooms could be used in the diet of people with high blood cholesterol levels 8 and other cardiovascular diseases. 52 It seems that our in vitro results could be extrapolated to in vivo conditions because the composition of the cellular membrane is closely associated with lipids present in the diet. 17 –20 Finally, some other fungal species with lipid profiles similar to those of M. procera (high PUFA/MUFA ratio, linoleic and pamitic acid, and low stearic acid), such as Agaricus arvensis, Agaricus campestris, Suillus granulatus, Helvella crispa, Lentinus squarullosus, Morchella conica, Pleurotus sajor-caju, and Clytocibe nebularis, 7,8,12 –14,53,54 should be further investigated for potential positive effects in hypertension.
Conclusion
Our results clearly demonstrate that the lipids from 2 European wild edible mushrooms—M. procera and C. platyphylla—provoked an increase in erythrocyte membrane fluidity. Hypertension, as well as some other cardiovascular diseases, is related to decreased fluidity of erythrocyte membrane, which is dependent on its lipid composition directly associated with lipids supplemented by diet. Thus, a diet that increases fluidity of erythrocyte membrane should be very beneficial for hypertension treatment. We found that selected wild edible mushrooms could match such dietary criteria. Positive effects on erythrocyte membrane fluidity, high PUFA/MUFA ratio, and high content of essential PUFA linoleic acid indicate that Macrolepiota procera could be used to treat hypertension. Other mushrooms showing similar lipid profiles should also be examined for positive effects in hypertension.
Footnotes
Author Disclosure Statement
No competing financial interests exist.