
Abstract
Soybean has many compounds with a variety of biological properties that potentially benefit human health; among them, isoflavones have inhibitory effects on lipid oxidation in adipose tissue. In this study, we examined two Korean traditional fermented soybean products—doenjang (DNJ) and cheonggukjang (CGJ)—for their ability to suppress redox-sensitive nuclear factor κB (NF-κB) activation in the kidney of rats fed a high-fat diet. Sprague–Dawley rats, 4 weeks old, were fed soybean, DNJ, or CGJ (1 g/kg/day) with a 20% fat diet for 6 weeks. Body weight and food intake were carefully monitored. NF-κB-related activities of genes for inflammatory proteins, such as inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), and vascular cell adhesion molecule-1 (VCAM-1), were determined. The soybean products exhibited antioxidative action by maintaining redox regulation, suppressing NF-κB activation, and modulating the expression of genes for NF-κB-induced inflammatory proteins such as COX-2, iNOS, and VCAM-1. Based on these results, we conclude that Korean traditional soybean fermented products, especially CGJ, suppress the generation of reactive species, NF-κB activity, and NF-κB-related inflammatory genes.
Introduction
S
According to a recent report, 2 soy saponins affect the production of pro-inflammatory mediators in lipopolysaccharide-stimulated peritoneal macrophages, significantly inhibiting prostaglandin E2, nitric oxide (NO), tumor necrosis factor-α (TNF-α), and monocyte chemoattractant protein-1. Moreover, soy saponins down-regulated the expression of cyclooxygenase-2 (COX-2) and inducible NO synthase (iNOS) and suppressed nuclear factor κB (NF-κB) activation by blocking IκBα degradation in the NF-κB signaling pathway. 2
Soy anthocyanins isolated from black soybean seed coat are known to inhibit levels of vascular cell adhesion molecule-1 (VCAM-1), intracellular adhesion molecule-1 (ICAM-1), and COX-2 induced by TNF-α, which is mediated by the NF-κB-dependent pathway. 3
Soy isoflavones have been shown to inhibit TNF-α-induced NF-κB activation in cultured human lymphocytes, and its supplementation in healthy men was shown to prevent NF-κB activation by TNF-α in blood lymphocytes. 4 Soy isoflavones also suppress cell adhesion and migration by inhibiting active NF-κB and activated protein-1 in breast cancer cells. 5 Soybean is known to suppress mitogen-activated protein kinase activation: the extracellular signal-regulated kinase 1/2 and the p38 signaling pathway-mediated NF-κB activation. 6
Deonjang (DNJ) and cheonggukjang (CGJ) are two traditional Korean fermented soybean foods. However, processing methods, fermentation conditions, and taste of the final product are quite different between DNJ and CGJ. For instance, to produce DNJ, dried soybeans are boiled and stone-ground into coarse bits. The resulting paste is then formed into blocks, which are called meju. Then, the meju is exposed to sunlight or warmth, allowing the fermentative growth of Bacillus subtilis. After 1–3 months, the meju blocks are put into large opaque pottery jars with brine and left to ferment further. After fermentation, the liquids and solids are separated into Korean soy sauce, called Joseon ganjang, and DNJ.
CGJ is also a fermented soybean paste, but one that is fermented by steaming soybeans at around 40°C for 2 or 3 days. CGJ is fermented mainly by B. subtilis. CGJ contains whole as well as ground soybeans, and its fermentation time is much shorter than that of DNJ. 7,8
In a recent study, DNJ was shown to have more antimutagenic effects than raw soybean. 9 CGJ, on the other hand, was shown to have a strong inhibitory effect on cancer cell growth in vitro and a stronger preventive effect against chemically induced mammary tumorigenesis in vivo, compared with steamed soybeans. 10 Fermented CGJ also suppresses the stimulation of tumor growth more than steamed soybean. 11 Therefore, soybean fermentation has stronger anti-inflammatory and tumor suppression effects than unfermented soybean.
To date, there has been little study of the anti-inflammatory effects of DNJ and CGJ in the high-fat diet-fed rat model. Many inflammatory peptides are linked to adiposity, which can be ultimately characterized as a state of low-grade systemic inflammation. 12 Increased oxidative stress, mainly from excessive macronutrient intake or increased metabolic rate, might be partly responsible for the inflammatory response. 13 Furthermore, reactive species (RS) are considered an essential contributor to obesity, 14,15 as RS production increases and accumulates, leading to oxidative stress. Thus, in a vicious cycle, obesity produces significantly higher RS and potentiates the pro-inflammatory state. RS affect many intracellular signaling pathways. 16 For example, NF-κB transcriptional activity is increased in overweight and obese humans. 17,18 Obesity also increases the activity of genes for inflammation-related proteins, including TNF-α, interleukin-6, monocyte chemoattractant protein-1, and soluble ICAM-1. 17 –20
Thus, we investigated the effects of the fermented soybean products DNJ and CGJ on oxidative stress, NF-κB activation, and pro-inflammatory gene expression using a high-fat diet-induced obese rat model.
Materials and Methods
Materials
We obtained steamed soybean and DNJ from Jinmi Foods (Daejeon, Republic of Korea) and CGJ from Moonokrae Food (Sunchang, Republic of Korea). Steamed soybean, DNJ, and CGJ were lyophilized before experimentation.
Animals
Specific pathogen-free, 4-week-old male Sprague–Dawley rats were obtained from Samtako (Osan, Republic of Korea). The animal protocol used in this study was reviewed and approved by the Committee of Institutional Animal Care and Use at Pusan National University (Busan, Republic of Korea). The rats were divided into five groups of six rats each. The first group was fed a normal diet (AIN-93M, Oriental Yeast, Tokyo, Japan). The second group was fed a high-fat diet (20% fat). The third, fourth, and fifth groups were fed a high-fat diet with 5% soybean, DNG, or CGJ, respectively. Soybean, DNJ, and CGJ were added at the level of 5 g/100 g of diet instead of casein. Compositions of diets are shown in Table 1. After 6 weeks of feeding, the rats were sacrificed by decapitation, and the kidneys were quickly removed and rinsed in ice-cold buffer (100 mM Tris, 1 mM EDTA, 0.2 M phenylmethylsulfonyl fluoride, 1 μM pepstatin, 2 μM leupeptin, 80 mg/L trypsin inhibitor, 20 mM β-glycerophosphate, 20 mM NaF, and 2 mM sodium orthovanadate, pH 7.4). The tissues were immediately frozen in liquid nitrogen and stored at −80°C.
ND, normal diet (based on the AIN-93M diet); HFD, high-fat diet (16% lard added to ND); HFD + soybean, HFD + 5% soybean (Jinmi Foods); HFD + DNJ, HFD + 5% doenjang soybean preparation (Jinmi Foods); HFD + CGJ, HFD + 5% cheonggukjang soybean preparation (Moonokrae Food); TBHQ, tert-butylhydroquinone.
Tissue preparation
All solutions, tubes, and centrifuges were maintained at 0–4°C. The preparation of nuclear extract was based on previous methods. 21 The kidney was used for the study because of its vulnerability to oxidative stress and inflammation stimuli. 22 Three hundred milligrams of kidney was homogenized with 2 mL of homogenate buffer A (10 mM HEPES [pH 7.8], 10 mM KCl, 2 mM MgCl2, 1 mM dithiothreitol, 0.1 mM EDTA, 0.1 mM phenylmethylsulfonyl fluoride, 1 μM pepstatin, and 1 mM p-aminobenzamidine) with a tissue homogenizer for 20 seconds. Homogenates were kept on ice for 15 minutes, 125 μL of 10% Nonidet P-40 was added, and the mixture centrifuged for 2 minutes at 14,320 g. The supernatant containing cytosol proteins was then centrifuged again, and the pelleted nuclei were washed once with 400 μL of buffer A plus 25 μL of 10% Nonidet P-40, centrifuged, suspended in 50 μL of buffer C (50 mM HEPES [pH 7.8], 50 mM KCl, 300 mM NaCl, 0.1 mM EDTA, 1 mM dithiothreitol, 0.1 mM phenylmethylsulfonyl fluoride, and 10% [vol/vol] glycerol), mixed for 20 minutes, and centrifuged for 5 minutes at 14,320 g. The supernatant containing nuclear proteins was stored at −80°C.
Measurement of RS
The basic reaction of this method is conversion of nonfluorescent 2,7-dihydrofluorescin diacetate (DCF-DA) into 2,7-dichlorodihydrofluorescein (DCFH) esterase and subsequently oxidation to highly fluorescent 2,7-dichlorofluorescein by RS. In brief, DCF-DA was dissolved in absolute ethanol at the concentration of 12.5 mM and kept at −70°C in the dark. Phosphate buffer (50 mM) at pH 7.4 was used to dilute DCF-DA stock to a working solution. DCF-DA (25 μM) was added to homogenate for a concentration of 250 μM in the final volume, and then changes in fluorescence intensity were measured for 30 minutes. The fluorescence was determined using a GENios microplate fluorescence reader (TECAN, Schweiz AG, Mannedorf, Switzerland) at excitation and emission wavelengths of 485 nm and 535 nm, respectively. DCFH diacetate (DCFH-DA) was obtained from Molecular Probes, Inc. (Eugene, OR, USA).
Nitrate/nitrite assay
Nitrate/nitrite levels were measured with a commercial kit according to the manufacturer's protocol (Cayman Chemical Co., Ann Arbor, MI, USA). In brief, a serum sample was ultrafiltered and loaded (20 μL) with assay buffer (260 μL) and enzyme cofactor mixture (10 μL), and then the nitrate reductase mixture (10 μL) was added. After a 1-hour incubation at room temperature, Griess Reagent R1 (50 μL) and Griess Reagent R2 (50 μL) were added. After 10 minutes at room temperature, absorbance was determined at 540 nm.
Protein analysis and western blotting
Protein concentrations were determined by the Folin phenol reagent method (Sigma, St. Louis, MO, USA) using bovine serum albumin as a standard. Homogenized samples were boiled for 5 minutes with a gel-loading buffer (pH 6.8) (composed of 0.125 M Tris-Cl, 4% sodium dodecyl sulfate, 10% 2-mercaptoethanol, and 0.2% bromphenol blue) at a ratio of 1:1. Equal amounts of protein were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis using 6–17% acrylamide gels. The gels were subsequently transferred onto a nitrocellulose membrane (Hybond C, Amersham Corp., Arlington Heights, IL, USA). The membrane was immediately placed in a blocking solution (5% nonfat dry milk in Tris-buffered saline with Tween [TBS-T] buffer containing 10 mM Tris, 100 mM NaCl, and 0.1% Tween 20, pH 7.5) at room temperature for 1 hour. The membrane washed in TBS-T buffer for 30 minutes and then incubated first with the antibody at room temperature for 2 hours. After three 10-minute washings in TBS-T buffer, the membrane was incubated with a second antibody at room temperature for 1 hour. After four 10-minute washing in TBS-T buffer, antibody labeling was detected using enhanced chemiluminescence following the manufacturer's instructions and exposed to radiographic film. Prestained blue protein markers were used for molecular weight determination. Immobilon-P transfer membrane was obtained from Millipore Corp. (Bedford, MA, USA). Antibodies to NF-κB p65, NF-κB p50, IκBα, COX-2, iNOS, and VCAM-1 were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Enhanced chemiluminescence western blotting detection reagents were from Amersham Life Science, Inc. (Arlington Heights).
Electrophoretic mobility shift assay
The electrophoretic mobility shift assay method was used to characterize the binding activities of NF-κB transcription factors in nuclear extracts. The NF-κB oligonucleotide sequence was 5′-GAGAGGCAAGGGGATTCCCTTAGTTAGGA-3′. Protein–DNA binding assays were performed with 20 μg of nuclear protein. Nonspecific binding was blocked by using 1 μg of poly(dI-dC)•poly(dI-dC). The binding medium contained 5% glycerol, 1% Nonidet P-40, 1 mM MgCl2, 50 mM NaCl, 0.5 mM EDTA, 2 mM dithiothreitol, and 10 mM Tris-HCl, pH 7.5. In each reaction, 20,000 cpm of a radiolabeled probe was included. Samples were incubated at room temperature for 20 minutes, and the nuclear protein with 32 P-labeled oligonucleotide complex was separated from free 32 P-labeled oligonucleotide by electrophoresis through a 5% native polyacrylamide gel in a running buffer containing 50 mM Tris (pH 8.0), 45 mM borate, and 0.5 mM EDTA. After separation was achieved, the gel was vacuum-dried for autoradiography. 23
Measurement of total phenolic contents
The total phenolic content was determined with Folin–Denis reagent (Fluka, St. Louis) using gallic acid as a standard. 24 Five grams of each lyophilized sample was defatted with 15 mL of ethyl ether during 2 hours of shaking. After shaking, the samples were separated from ethyl ether. Two hundred milligrams of the separated sample was re-extracted for 2 hours with 2 mL of 80% ethyl alcohol containing 1% hydrochloric acid at room temperature on an orbital shaker set at 200 rpm. The mixture was filtered through No. 2 filter paper (Toyo Roshi Kaisha, Tokyo). Filtrates were used for the total phenolic content determination. Two hundred microliters of the filtrate was mixed with 5.5 mL of 10-fold diluted Folin–Denis reagent, the mixture was kept at 22°C for 5 minutes, and then 1 mL of 2% Na2CO3 was added to the mixture. Absorbance was read after 90 minutes at 725 nm.
Statistical analysis
The results are presented as mean ± SE values of three individual experiments, and each measurement was performed in triplicate. Statistical significance was tested using one-way analysis of variance followed by Newman–Keuls Multiple Comparison Test. Statistical significance was set at P < .05.
Results
Effects of DNJ and CGJ feeding on food intake and body weight
Changes in body weight, food intake, and food efficiency ratio are shown in Table 2. The initial weights of the rats were almost the same among the groups, but 6 weeks later, the weights were quite different. The final body weight of the high-fat diet group was 500.7 g, whereas that of the high-fat diet with CGJ group was 426.5 g. Food intakes among the five groups ranged from 19.9 g/day to 21.3 g/day during the 6 weeks. However, body weight gains ranged from 2.8 g/day to 5.7 g/day. Therefore, CGJ may inhibit body weight gain. (Some of the data presented in Table 2 came from a published doctoral dissertation. 25 )
Means with different letters in the same row are significantly different (P < .05) by Duncan's multiple range test.
ND, normal diet (based on the AIN-93M diet); HFD, high-fat diet (16% lard added to ND); HFD + soybean, HFD + 5% soybean (Jinmi Foods); HFD + DNJ, HFD + 5% doenjang soybean preparation (Jinmi Foods); HFD + CGJ, HFD + 5% cheonggukjang soybean preparation (Moonokrae Food).
Suppression of RS level by soybean and fermented soybean products
First, high-fat-induced RS generation in rats fed a high-fat diet was studied using the DCFH-DA method in kidney homogenate. The results showed increased RS levels with the high-fat diet. However, RS production was suppressed by soybean and soybean products (Fig. 1), indicating that fermented soybean, especially CGJ, had a strong antioxidant effect on the rats fed the high-fat diet. The group of rats fed high-fat diet with CGJ had a 48% reduction in RS generation compared with the group fed the high-fat diet alone.
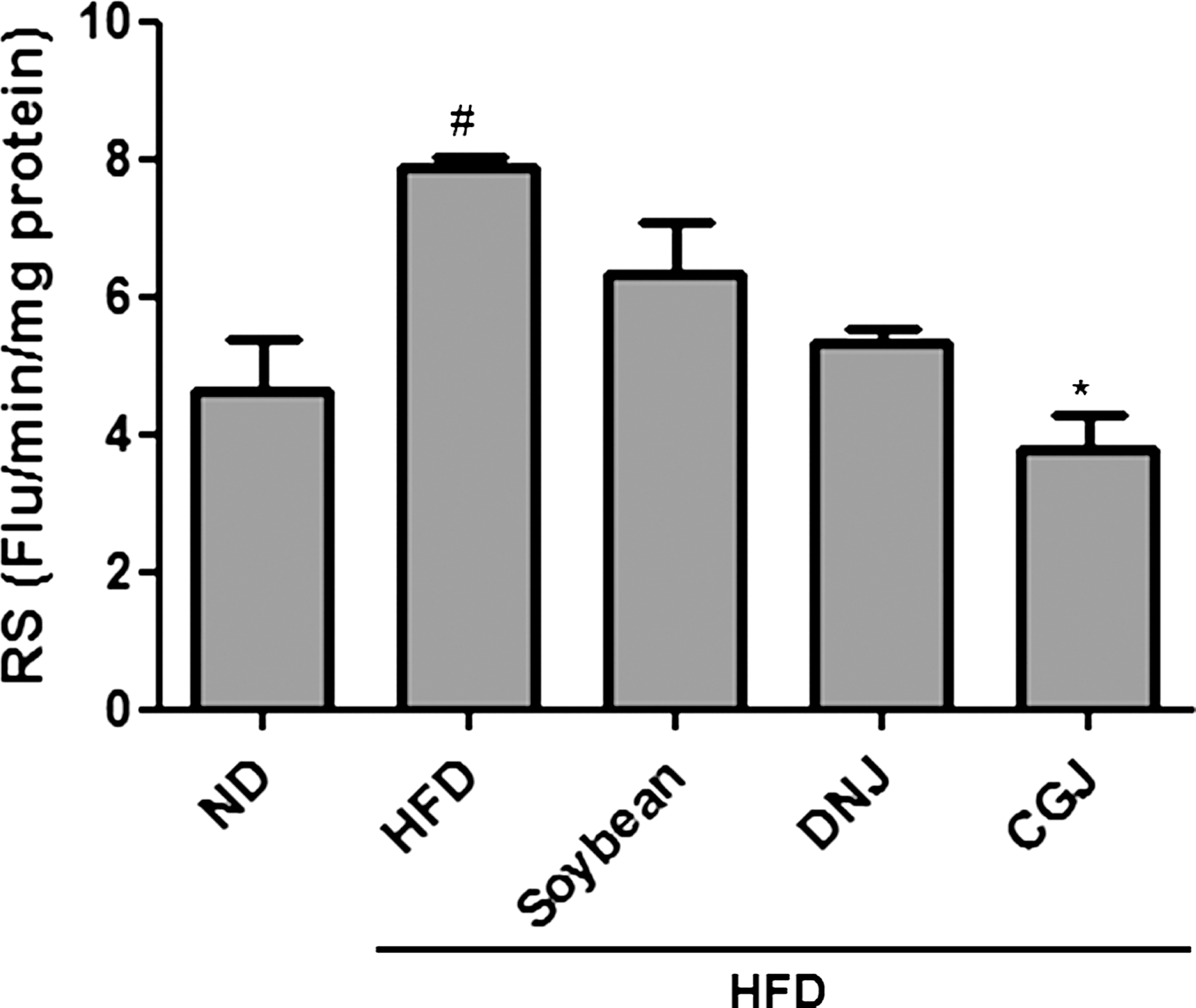
Suppression of reactive species (RS) levels by soybean products. RS generation in Sprague–Dawley rats was determined by using the 2,7-dihydrofluorescin diacetate method in kidney homogenate. Data are mean ± SE values from six rats. Statistical significance: *P < .05, **P < .01 compared with HFD. ND, normal diet; HFD, high-fat diet; Soybean, HFD with 5% cooked soybean; DNJ, HFD with 5% doenjang; CGJ, HFD with 5% cheonggukjang; Flu, fluorescence.
Suppression of NO level by soybean and fermented soybean products
NO derived from iNOS is mainly a mediator of inflammation. The NO level in serum was determined using the Griess assay. The result showed that the high-fat diet group had increased NO levels, but NO levels were suppressed by soybean and soybean products (Fig. 2). This result suggests that soybean and fermented soybean products modulated the inflammation mediator NO in the rats fed the high-fat diet.

Suppression of nitric oxide levels in serum by soybean products. The nitric oxide level in Sprague–Dawley rats was determined by using a nitrate/nitrite colorimetric assay kit (Cayman Chemical Co.) in serum. Data are mean ± SE values from six rats. Statistical significance: ### P < .001 compared with ND, ***P < .001 compared with HFD. ND, normal diet; HFD, high-fat diet; soybean, HFD with 5% cooked soybean; DNJ, HFD with 5% doenjang; CGJ, HFD with 5% cheonggukjang.
Suppression of NF-κB by soybean and fermented soybean
Activation of NF-κB signaling by oxidative stress leads to phosphorylation and proteolysis of IκB. Following stimulation, NF-κB–inducing kinase activates IκB kinase α/β and initiates IκBα phosphorylation. Phosphorylated IκBα is then ubiquitinated, thereby releasing NF-κB dimers (p65 and p50) into the cytoplasm and allowing them to translocate into the nucleus. Western blot analysis was probed with antibodies specific for cytosol IκBα and nuclear p65 and p50 in kidney homogenate.
The resulting data demonstrate that the NF-κB level significantly increased with feeding of the high-fat diet. In contrast, soybean and fermented soybean products, especially CGJ, decreased p65 and p50 protein levels in nuclear extracts and increased IκBα protein levels in cytosolic extracts (Fig. 3). The NF-κB binding activity was determined with nuclear protein by electrophoretic mobility shift assay (Fig. 4). The high-fat diet was shown to increase NF-κB binding activity compared with the normal diet. However, the soybean and fermented soybean diets significantly inhibited NF-κB binding activity compared with the high-fat diet. The CGJ-supplemented diet showed the strongest effect on the suppression of NF-κB activation. This finding demonstrated that soybean and fermented soybean products, especially CGJ, suppressed NF-κB activation through the IκB pathway.
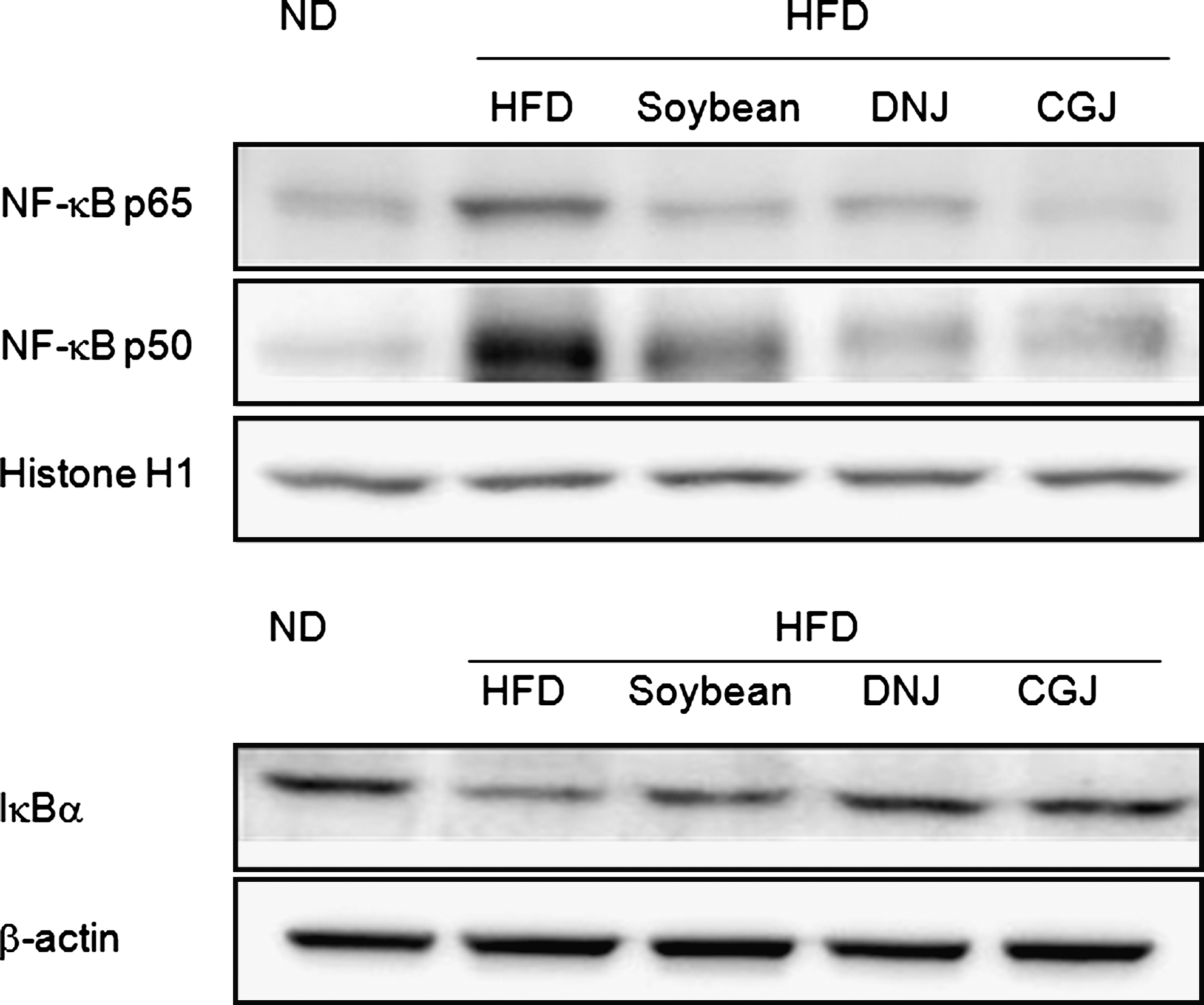
Suppression of nuclear factor κB (NF-κB) translocation and activation by soybean products. Western blot analysis was performed to detect NF-κB (p65, p50) and histone H-1 protein levels in nuclear extracts (40 μg of protein) and to detect IκBα and β-actin protein levels in cytosol extracts (50 μg of protein). Western blot analysis was probed with antibodies specific for p65, p50, histone H-1, IκBα, and β-actin. ND, normal diet; HFD, high-fat diet; soybean, HFD with 5% cooked soybean; DNJ, HFD with 5% doenjang; CGJ, HFD with 5% cheonggukjang.

Inhibition of NF-κB DNA binding activity by soybean products. The electrophoretic mobility shift assay method was used to determine nuclear NF-κB binding activities using 10 μg of nuclear extract. BL, blank as absence of nuclear extract; ND, normal diet; HFD, high-fat diet; Soy, HFD with 5% cooked soybean; DNJ, HFD with 5% doenjang; CGJ, HFD with 5% cheonggukjang; Cold, competition assay using a 100-fold excess of an unlabeled NF-κB DNA oligonucleotide.
Inhibition of NF-κB-dependent genes by soybean and fermented soybean products
To further elucidate the regulatory role of soybean and fermented soybean through NF-κB activation, we determined the effects of soybean and fermented soybean on several NF-κB targeting genes. COX-2 and iNOS are known to have an NF-κB binding site in their promoter region and are controlled by NF-κB regulation. NF-κB activation increased the expression of the adhesion molecule, VCAM-1. The high-fat diet remarkably increased expressions of these pro-inflammatory genes, but soybean and fermented soybean products, especially CGJ, reduced their expression (Fig. 5). These results suggest that soybean and fermented soybean inhibit the expression of pro-inflammatory genes by suppressing NF-κB activation.
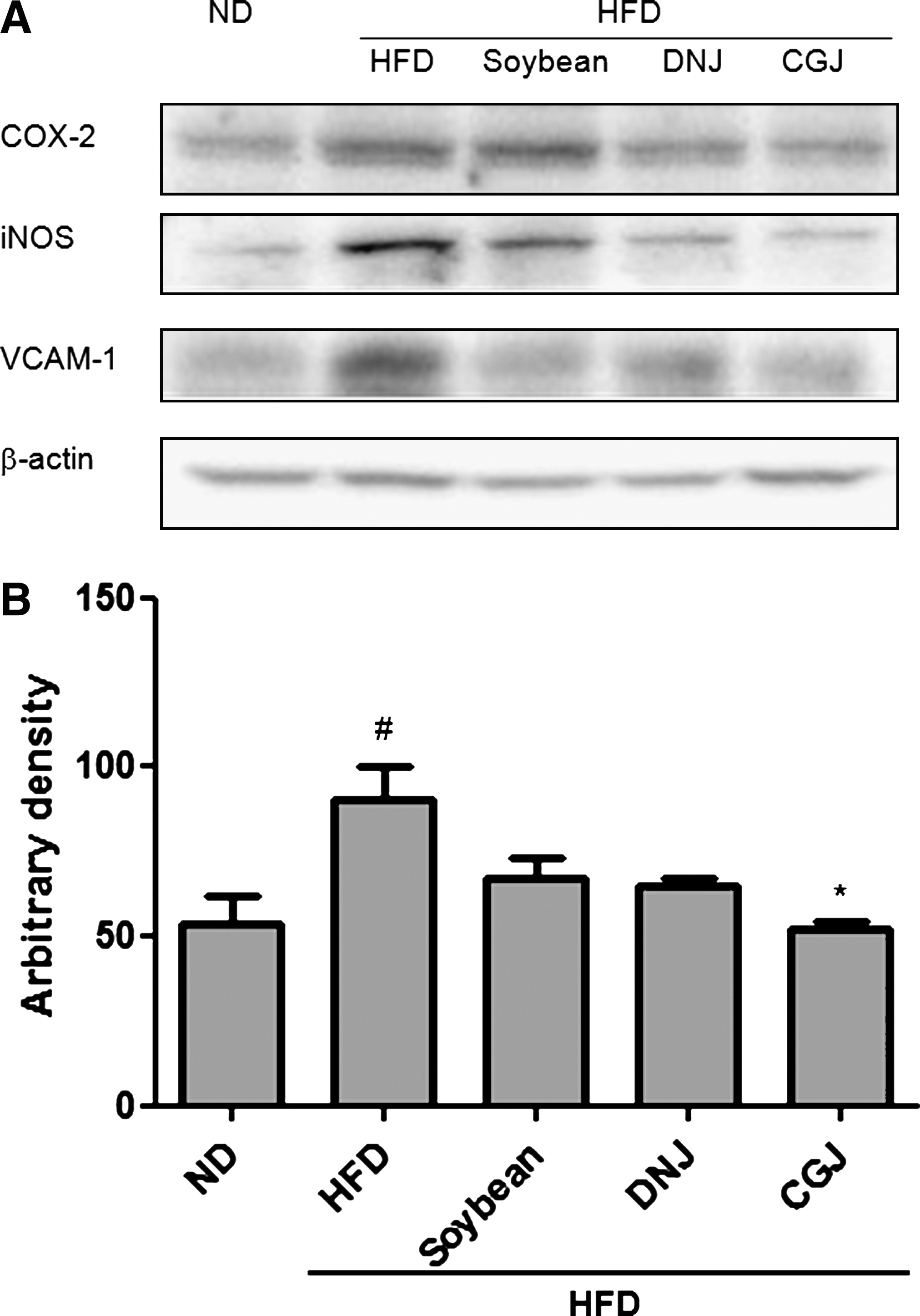
Inhibition of NF-κB-dependent genes by soybean products. ND, normal diet; HFD, high-fat diet; soybean, HFD with 5% cooked soybean; DNJ, HFD with 5% doenjang; CGJ, HFD with 5% cheonggukjang. (
Discussion
In the current study, we obtained evidence that soybean and the fermented soybean products DNJ and CGJ modulate inflammation-related NF-κB activation in Sprague–Dawley rats fed a high-fat diet. Our results indicate that CGJ more efficiently suppressed both high-fat diet–related RS generation and activation of NF-κB than DNJ.
Soybeans are used in many products. Nonfermented soyfoods include fresh green soybeans, whole dry soybeans, soy nuts, soy sprouts, whole-fat soy flour, soy milk and soymilk products, tofu, okara, and yuba. Fermented soy foods include DNJ, CGJ, tempeh, miso, soy sauces, natto, fermented tofu, and fermented soymilk products. Recent research into the physiological effects of soybean products revealed that fermented soybean foods have increased antihypertensive, antimutagenic, antioxidative, and antitumor bioactivity. These beneficial effects were based on their release of isoflavone, saponins, phenolic acids, and phytic acid. 26
Oxidative stress is considered a major cause of molecular inflammation and of many inflammation-related diseases such as atherosclerosis, arthritis, dementia, osteoporosis, cardiovascular diseases, and aging. Soy products regulate the inflammatory process by modulating the redox environment through their antioxidative properties. Among the key cellular components exquisitely sensitive to redox status are transcriptions factors such as NF-κB, COX-2, iNOS, and the adhesion molecule, VCAM-1, all of which are associated with molecular inflammation. 27,28
In our study, soybean and soybean fermented products were shown to scavenge RS and inhibit the inflammatory response by regulating the redox environment through its antioxidative properties. Also, soybean was shown to suppress molecular inflammation.
RS production is an established, essential contributor to oxidative stress. 14,15 As RS production increases, oxidatively damaged molecules accumulate, resulting in oxidative stress. Protein, lipid, and DNA are all directly modified by RS; RS also affects the intracellular signaling pathway. 16 Oxidative stress increases with obesity, mainly from excessive macronutrient intake, increased visceral adiposity or increased metabolic rate, which might be partly responsible for the inflammatory response. 13,29 However, Swain et al. 30 reported that dietary soy protein protects against oxidative stress in perimenopausal women. Also, Rossi et al. 31 reported that soy beverage consumption increases the total antioxidant status in young men. Our current data support the findings of these investigators.
We found that soybean and fermented soybean products suppressed NF-κB p65 and p50 protein levels in nuclear extracts and increased IκBα protein levels in cytosolic extracts. These findings demonstrated that soybean and fermented soybean products, especially CGJ, suppressed NF-κB activation through the IκB pathway. Yang et al. 32 and Fan et al. 33 reported that soy milk fermented by Bifidobacterium and Streptococcus strains enhanced free radical scavenging activities. Moreover, Kobayashi et al. 6 reported that soybean suppressed mitogen-activated protein kinase activation: the extracellular signal-regulated kinase 1/2 and p38 signaling pathway mediated NF-κB activation. Also, Kang et al. 2 reported that soybean saponins suppressed NF-κB activation in lipopolysaccharide-stimulated peritoneal macrophages by blocking IκBα degradation in the NF-κB signaling pathway. Our data also show that expressions of NF-κB-related pro-inflammatory genes, notably COX-2, iNOS, and that of the adhesion molecule VCAM-1, increased with the feeding of a high-fat diet, but that soybean and fermented soybean products modulated these gene expressions.
Possible explanations for our current data on the increased expression of NF-κB-related pro-inflammatory genes such as COX-2, iNOS, and that of the adhesion molecule VCAM-1 may be found in the work of other studies. For instance, Kang et al. 2 reported that soy saponins suppressed the production of pro-inflammatory mediators in lipopolysaccharide-stimulated peritoneal macrophages; significantly, soy saponins inhibited prostaglandin E2, NO, TNF-α, and monocyte chemoattractant protein-1 and the expression of COX-2 and iNOS. Davis et al. 4 reported that soy isoflavone inhibited TNF-α-induced NF-κB activation in cultured human lymphocytes; soy isoflavone supplementation was also reported to prevent NF-κB activation by TNF-α in blood lymphocytes of healthy men. Kim et al. 3 reported that anthocyanins isolated from black soybean seed coat inhibited VCAM-1, ICAM-1, and COX-2 levels via TNF-α, through its NF-κB-dependent pathway.
In the current study, CGJ consistently outperformed soybean and DNJ in modulating RS generation and the NF-κB system, presumably because of active compounds such as isoflavone. Although the isoflavone content was not measured in the samples, one could speculate that this finding was due to the high level of isoflavone in CGJ. Isoflavones found in unprocessed food have a glycoside structure, not in a free form, therefore making intestinal absorption very poor. However, during the fermentation process, such as that of CGJ, glycoside is hydrolyzed by β-glucosidase secreted by microorganisms, resulting in active aglycone. 34 –36 Park et al. 37 reported the quantitative conversion of isoflavone to aglycone during CGJ fermentation. Lee et al. 38 reported a higher isoflavone content of CGJ compared with soybean or DNJ; the isoflavone aglycone level of 809 μg/g found in soybean was increased to 873 μg/g in DNJ and 1,045 μg/g in CGJ following fermentation. Although we did not measure the amount of isoflavone aglycone in our study, the total phenolic content of CGJ was higher than DNG as shown in Figure 6.
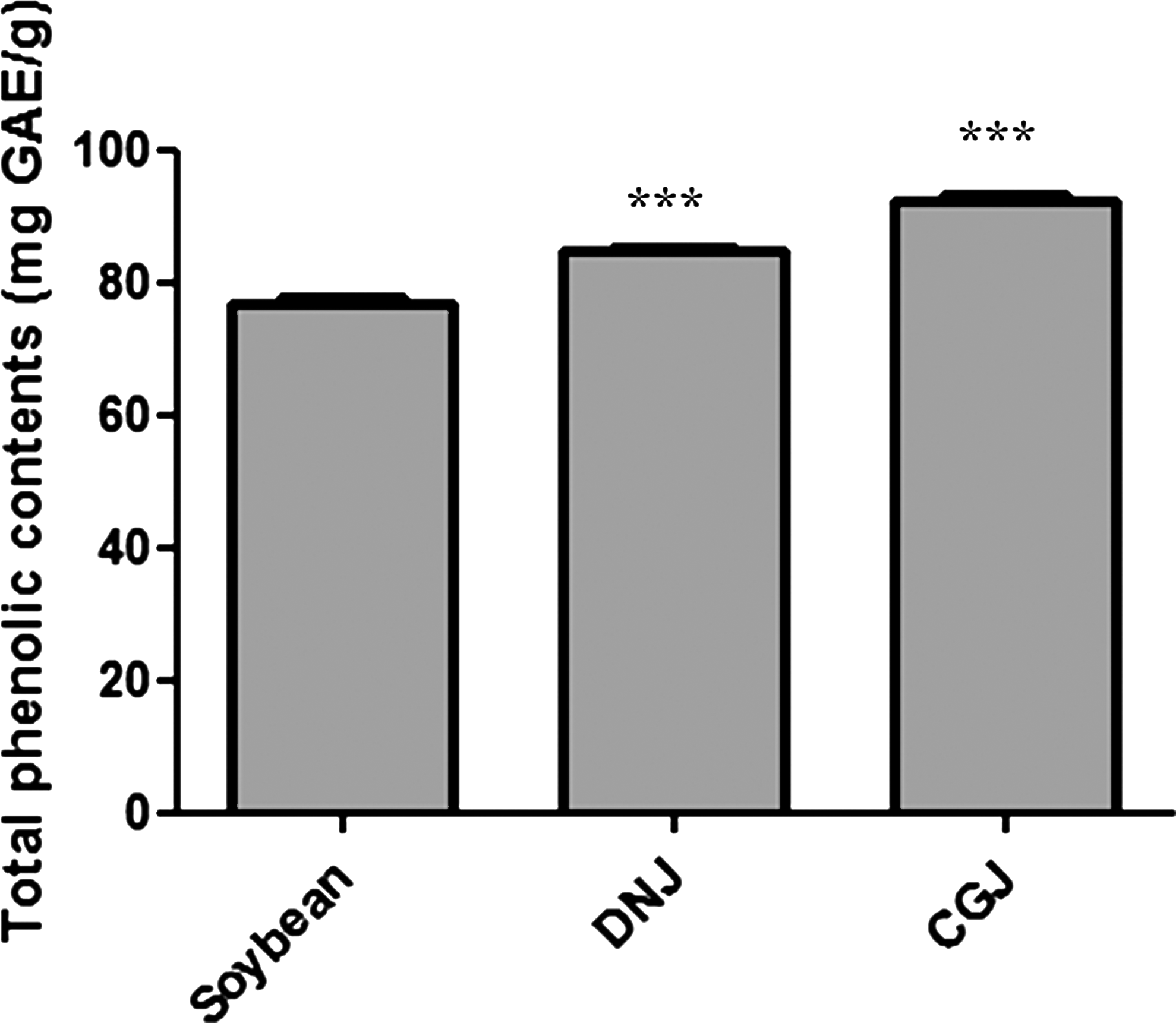
Total phenolic contents in soybean, doenjang (DNJ), and cheonggukjang (CGJ). Total phenolic contents were determined by using the Folin–Denis reagent. Statistical significance: ***P < .001 compared with soybean. GAE, gallic acid equivalents.
Similar results are seen with tempeh, when soybean cake is fermented for 24 hours—the aglycone content doubled. 35 In addition, Marazza et al. 39 reported soymilk fermentation by Lactobacillus rhamnosus CRL981 increased isoflavone aglycone after 12-hour fermentation, producing different aglycone-rich soybean products. Together, these studies suggest that fermenting soy bean enhances bioactivity.
In summary, our data show that CGJ and DNJ preparations significantly decreased high-fat diet–related RS accumulation and NF-κB activation. In particular, CGJ decreased the activation of NF-κB and expressions of NF-κB-dependent pro-inflammatory genes and adhesion molecules. These data support the beneficial role of CGJ in the maintenance of health of the general public who regularly consume fermented soybean foods by decreasing the pro-inflammatory activity.
Footnotes
Acknowledgments
This study was supported by the Technology Development Program (grant 105075-3) for Agriculture and Forestry, Ministry for Agriculture, Forestry and Fisheries, Republic of Korea and National Research Foundation of Korea grant 2009-0083538 funded by the Korean Government. We thank the Aging Tissue Bank for supplying research resources.
Author Disclosure Statement
No competing financial interests exist.