
Abstract
This study investigated the anxiolytic-like effects of 4-O-methylhonokiol, a neolignan compound of Magnolia officinalis, by using the experimental paradigms of anxiety and compared the results with those of a known anxiolytic, diazepam. A single treatment with 4-O-methylhonokiol (0.1, 0.2, and 0.5 mg/kg, p.o.) or treatment for 7 days (0.5 mg/kg in drinking water) increased the percentage of time spent in the open arms and the number of open arms entries in the elevated plus-maze test. However, the 4-O-methylhonokiol-increased percentage of time spent in the open arm was abolished by treatment with flumazenil, a benzodiazepine receptor antagonist (10 mg/kg). 4-O-Methylhonokiol also increased the number of head dips in the hole-board test, but decreased locomotor activity. Molecular experiments revealed that the α1-subunit of γ-aminobutyric acid (GABA) type A receptors was overexpressed in the cortex of brains of mice after treatment with 4-O-methylhonokiol for 7 days. In addition, 4-O-methylhonokiol also increased chloride influx in cultured cortical cells. It is concluded that 4-O-methylhonokiol may have anxiolytic-like effects and that these effects may be mediated by GABAergic transmission with the increase of Cl– channel opening.
Introduction
A
The γ-aminobutyric acid (GABA)inergic system is the most common target of anxiolytic drugs. GABA is the main inhibitory neurotransmitter in the brain, 11 and it is known to interact with three receptor types: GABAA, GABAB, and GABAC. The GABAA receptors are probably the most complicated heteromeric ligand-gated Cl– channels that cause the negative Cl– to flow into the cell, leading to hyperpolarization and thus inhibition of cellular activity. 12 Thus, GABAA receptors are important drug targets for a variety of medications used to treat anxiety and insomnia.
Magnolia officinalis is known to contain many pharmacologically active components, with magnolol, obovatol, and honokiol being the main constituents. They have been used as traditional medicines in China and Japan and have a wide spectrum of pharmacological activities. 13 These compounds exhibit central depressant, muscle-relaxing, and anticonvulsant effects, as well as neuroprotective effects against chemical and heat damages. 14 –18 It was found that honokiol and magnolol have anxiolytic activities. 19 We also found that obovatol isolated from Magnolia obovata has an anxiolytic effect. 20 In our previous study, we found that 4-O-methylhonokiol, a novel compound from M. officinalis, promotes neurite outgrowth in a concentration-dependent manner in rat embryonic neuronal cells 21 as well as having memory-improving activities. 21,22 In the present study, we investigated the anxiolytic-like effects of single or repeated treatments of 4-O-methylhonokiol using the experimental paradigms of anxiety such as the elevated plus-maze and hole-board tests and compared the results with those of a known anxiolytic compound, diazepam. In addition, chloride influx in cortical neurons and expressions of GABAA receptor subunits in cortex and hippocampus of brains of mice after the mice were administered 4-O-methylhonokiol were measured to elucidate the possible anxiolytic mechanisms of 4-O-methylhonokiol. We found that 4-O-methylhonokiol showed anxiolytic-like effects that may be mediated by GABAergic transmission accompanied by an increase in Cl– channel opening.
Materials and Methods
Animals
Male ICR mice (Samtako, Osan, Republic of Korea) weighing 20–25 g, in groups of 10–12, were used for behavioral experiments. Animals were housed in acrylic cages (45×60×25 cm) with water and food available ad libitum, a 12-hour light/dark cycle (light on at 7:00 hours), and a constant temperature (22±2°C) according to the guidelines for care and use of laboratory animals of the animal ethics committee of Chungbuk National University (Cheongju, Republic of Korea). Mice were housed acclimated for 1 week before testing to ensure adaptation to the new environment. All of the behavior experiments were performed between 10:00 and 17:00 hours. Mice were randomly allocated to the following groups: control nontreated group, diazepam (2.0 mg/kg p.o.)-treated positive control group, and 4-O-methylhonokiol (0.1, 0.25 and 0.5 mg/kg, p.o)-treated groups.
Experimental compounds and drugs
4-O-Methylhonokiol (Fig. 1) was isolated and identified from bark of M. officinalis as described elsewhere; 21,23 pure (99.1%) 4-O-methylhonokiol was obtained from Bioland Co. (Chungnam, Republic of Korea). 4-O-Methylhonokiol was dissolved in 0.05% ethanol and then added to drinking water, which was provided ad libitum, and levels were measured for 7 days prior to the plus-maze test. Single treatment was done by oral administration at a volume of 0.1 mL/10 g of body weight with free access to diet. Diazepam was obtained from Myung In Pharr. Co., Ltd. (Kyunggi-Do, Republic of Korea) and was dissolved in 0.9% NaCl (physiological saline) before experiments. All other chemicals including flumazenil were obtained from Sigma Chemical Co. (St. Louis, MO, USA), and the solutions were prepared freshly before the experiments. Single treatment was done with 4-O-methylhonokiol at 0.1, 0.2 and 0.5 mg/kg, p.o., and repeat treatment was done with 4-O-methylhonokiol at 0.5 mg/kg of body weight added to drinking water by calculation of the amount of consumption per day for 7 days.

Chemical structure of 4-O-methylhonokiol.
Elevated plus-maze test
The elevated plus-maze apparatus consists of four arms (30×5 cm) elevated 45 cm above the floor. Each arm was positioned at 90° relative to the adjacent arms. The two enclosed arms had 30-cm-high walls, and to facilitate grip on the open arms these included a raised edge of 0.25 cm. Open and closed arms were connected via a central area (5×5 cm) to form a plus sign. The maze floor was constructed of black plexiglass, and the walls of the enclosed arms were constructed of clear plexiglass. 24 Four 25-W red fluorescent lights arranged as a cross at 100 cm above the maze were used as the source of illumination, and the video camera was suspended above the maze to record movements for analysis. Mice were randomly assigned (with a slight adjustment for matched body weight) to experimental groups. The test commenced by placing a mouse on the central platform of the maze facing an open arm. The number of entries into and the time spent in each of the two types of arms were recorded during the 5-minute trial. 25,26 All experimental sessions were recorded with a video camera mounted vertically above the maze. The activity on the open arm was evaluated as (1) time spent in the open arms relative to the total time spent in the plus maze, expressed as a percentage (100×time spent in the open arm/total time in the plus maze), and (2) the number of entries into both the open and closed arms, expressed as a percentage (100×open arm/total entries). An arm entry was defined as all four paws having crossed the dividing line between an arm and the central area.
Hole-board test
The hole-board apparatus (Ugo Basile, Comerio, Italy) consisted of gray Perspex® (Lucite International, Southampton, United Kingdom) panels (40×40 cm, 2.2 cm thick) with 16 3-cm holes spaced equidistantly on the floor. Photocells below the surface of the hole measured the number of head dips. The board was positioned 15 cm above a table. Mice were randomly allocated to the following groups: control, diazepam-treated, and 4-O-methylhonokiol-treated groups, as described above. Each mouse was individually placed on the center of the board facing away from the observer and allowed to freely roam about the apparatus prior to the testing. Diazepam and 4-O-methylhonokiol were administered 30 and 60 minutes before the test, respectively. The number of head dips on the hole-board was counted for 5 minutes. 27 –29 After each trial, the floor of the apparatus was wiped with absolute methanol to remove traces of previous paths. This test session also was recorded with a camera mounted vertically above the hole-board test.
Spontaneous locomotor activity
Spontaneous locomotor activity was measured automatically with a tilting-type ambulometer (model AMB-10, O'Hara & Co., Ltd., Tokyo, Japan). Each mouse was placed in the activity cage (20 cm in diameter, 18 cm in height) after an adaptation period of 10 minutes. Diazepam was administered orally 30 minutes prior to the experiment as described elsewhere. 20 4-O-Methylhonokiol was administered orally 60 minutes prior to the experiment. Ambulation activity was measured for 30 minutes after oral administration of the agents.
Cell culture
Primary cultured brain cells were prepared from cortex of 18-day-old Sprague–Dawley rat embryos as previously described.
30
In brief, cells were plated at 1×106 cells per 0.2 mL in 96-well microplates or 2×106 cells per 2.0 mL in 60-mm-diameter dishes precoated with poly-
The intracellular Cl– concentration in primary cultured brain cells was estimated using the Cl–-sensitive fluorescence probe N-(ethoxycarbonylmethyl)-6-methoxyquinolinium bromide (MQAE) according to the method of West and Molloy
31
with a slight modification. The buffer (pH 7.4) used contained the following: 2.4 mM HPO4
2–, 0.6 mM H2PO4
–, 10 mM HEPES, 10 mM
Expression of GABAA receptor subunits
After the behavior experiments, the mice were decapitated, brains were removed, and cortex and hippocampus were dissected on ice. Cortex and hippocampus tissue were homogenized with lysis buffer, and the extracts were centrifuged at 20,000 g for 20 minutes. Equal amount of proteins were separated on a sodium dodecyl sulfate/12% polyacrylamide gel and transferred to a nitrocellulose membrane (Hybond ECL, Amersham Pharmacia Biotech Inc., Piscataway, NJ, USA). The blots were blocked overnight at 4°C with 5% (wt/vol) nonfat dried milk in a Tris-buffered saline solution (10 mM Tris [pH 8.0] and 150 mM NaCl) containing 0.05% Tween-20 (TBST). The membrane was incubated with the specific antibodies, rabbit polyclonal antibodies against GABAA receptor subunits (diluted 1:500) (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), overnight at 4°C. The blot was washed in TBST three times and then incubated for 2 hours with the corresponding conjugated anti-rabbit immunoglobulin G–horseradish peroxidase (diluted 1:2,000) (Santa Cruz Biotechnology Inc.). The membrane was washed in TBST three times, and the immunoreactive proteins were detected using the ECL western blotting detection system (Amersham Pharmacia Biotech Amersham, United Kingdom).
Statistical analysis
Data were analyzed using GraphPad Prism 4 software (version 4.03, GraphPad Software, Inc., San Diego, CA, USA). The results are presented as mean±SEM values, and the significance of the effects of the compounds was assessed using analysis of variance. In the case of significant variation (P≤.05), the individual values were compared with Dunnett's test.
Results
Effects of 4-O-methylhonokiol on the elevated plus-maze model
As the positive control, diazepam at 2 mg/kg significantly increased open arm entries and time spent in open arms compared with the vehicle group (determined at 7 days after treatment). The mice treated with 4-O-methylhonokiol had a longer percentage of time spent in open arms compared with that of the saline-treated control animals in a dose-dependent manner, with 0.5 mg/kg treatment being the longest (Fig. 2A). However, the percentage of open arm entries was not significantly changed. Thus, we compared the percentage of number of open arm entries and time spent in open arms at 1, 3, 5, and 7 days after treatment with 4-O-methylhonokiol. Diazepam significantly increased both the percentages of number of entries and time spent throughout the assay period; 4-O-methylhonokiol significantly increased percentage of number of entries in open arms only on days 1 and 5, whereas the time spent increased significantly at days 1, 5, and 7 (Fig. 2C and D).
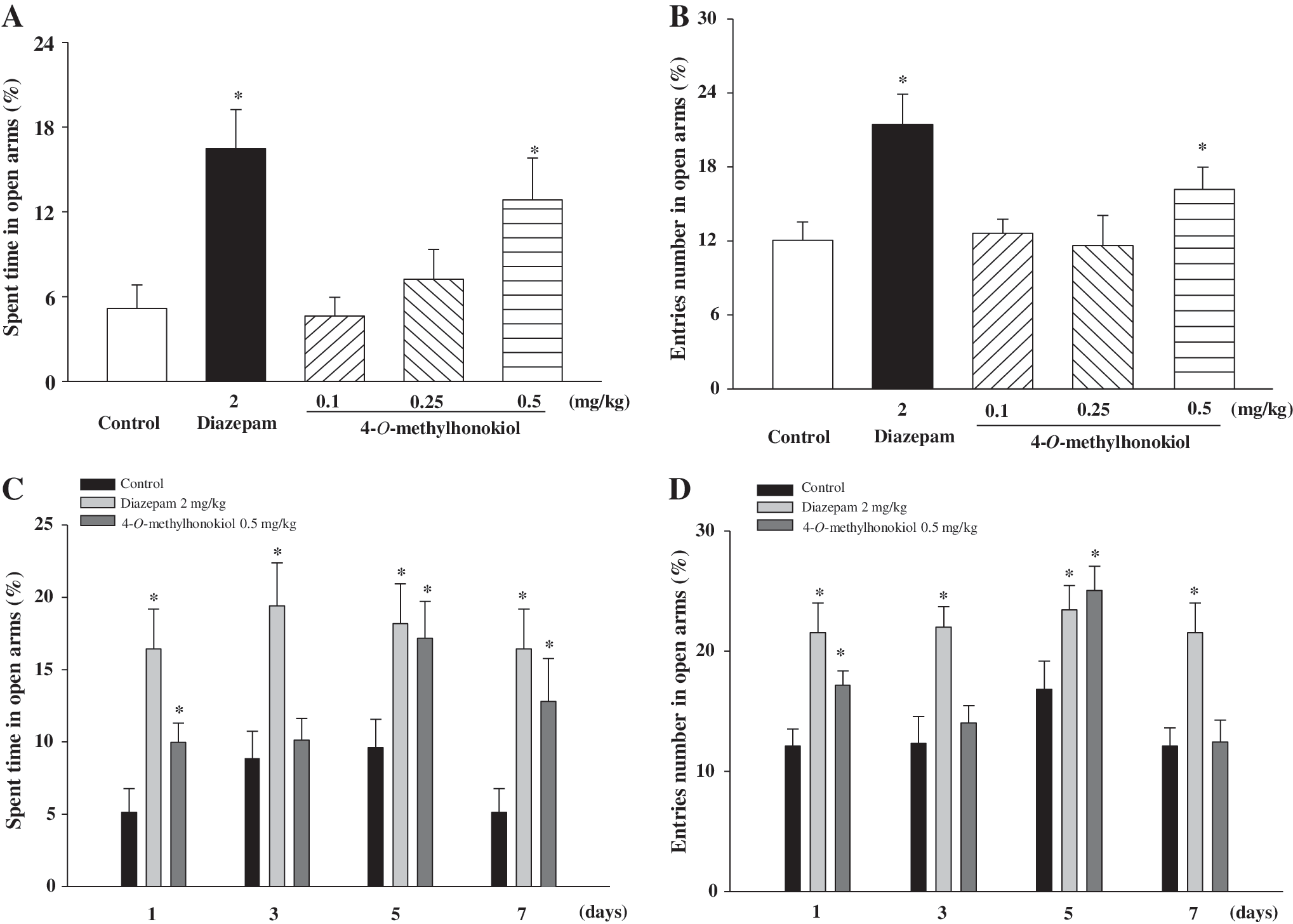
Effects of 4-O-methylhonokiol on the percentage of open arm entries and time spent in open arms of the elevated plus-maze by mice (n=9–11). Open arm entries and time spent in open arms of the elevated plus-maze were measured for 5 minutes, 30 minutes, or 1 hour after oral administration of diazepam or 4-O-methylhonokiol: (
In order to determine whether the anxiolytic-like effects of 4-O-methylhonokiol are exerted via GABAergic systems, 4-O-methylhonokiol-treated mice were subjected to a co-treatment with flumazenil, a benzodiazepine receptor antagonist. The anxiolytic-like effects of 4-O-methylhonokiol were abolished by flumazenil (10 mg/kg) as determined by the elevated plus maze test (Fig. 3).
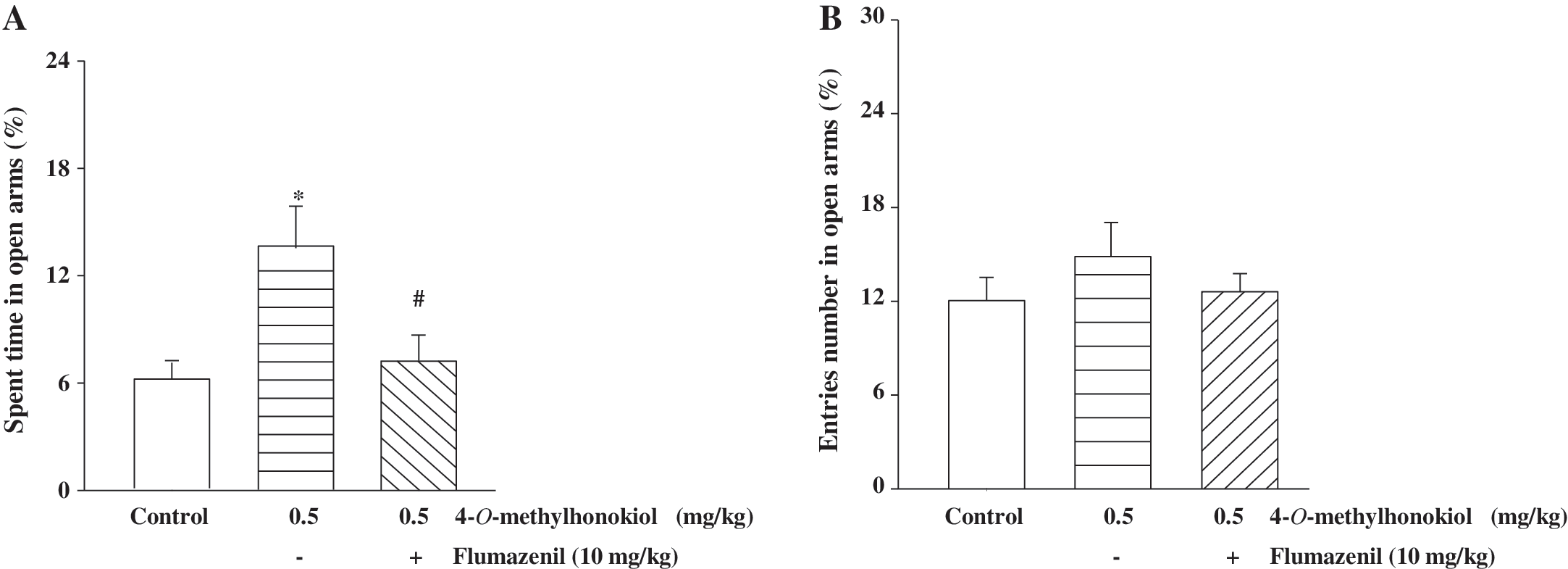
Reversals of 4-O-methylhonokiol-induced anxiolytic-like effects by flumazenil. Flumazenil, a benzodiazepine receptor antagonist, was intraperitoneally pretreated 30 minutes or 1 hour prior to the oral administration of diazepam or 4-O-methylhonokiol, respectively. Data are mean±SEM values from 10 mice. *P<.05 compared with the saline group. # P<.05 compared with the 4-O-methylhonokiol-treated group.
Effects of 4-O-methylhonokiol on the hole-board test
In order to investigate the exploratory and anxiolytic effects, 32 the hole-board test was used to evaluate the animals following treatment with diazepam or 4-O-methylhonokiol. Because the elevated plus-maze test showed that chronic treatment did not show any time-dependent difference, the hole-board test was done after single dose treatment. As shown in Figure 4, both diazepam (2 mg/kg) and 4-O-methylhonokiol (dose dependently and significantly) increased the number of head dips.
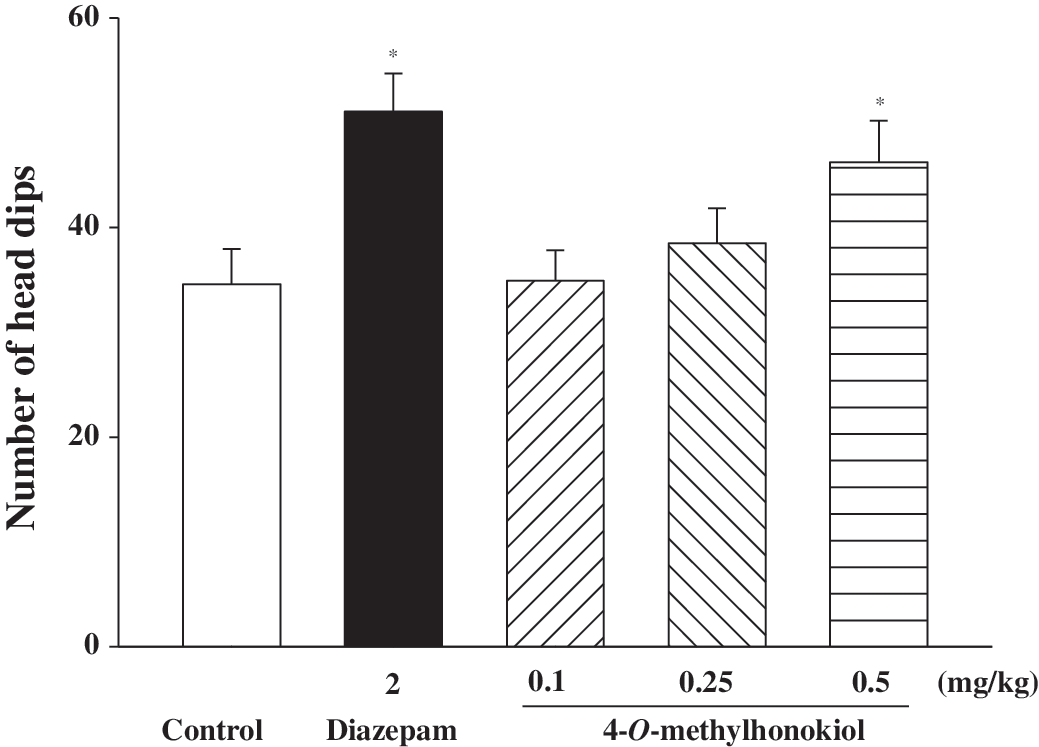
Effects of 4-O-methylhonokiol on head dips in the hole-board test in mice (n=10–12). Head dips in the hole-board test were measured for 5 minutes, 30 minutes, or 1 hour after oral administration of diazepam or 4-O-methylhonokiol. Data are mean±SEM values. *P<.05 compared with the control group.
Effects of 4-O-methylhonokiol on locomotor activity
Because the plus-maze experiment was affected by changes in spontaneous locomotor activity, spontaneous locomotor activity was measured. Spontaneous locomotor activity was significantly decreased by diazepam (1.0 mg/kg). Spontaneous locomotor activity was also dose-dependently decreased in animals pretreated with 4-O-methylhonokiol. The inhibitory effect was similar to that of diazepam (Fig. 5).
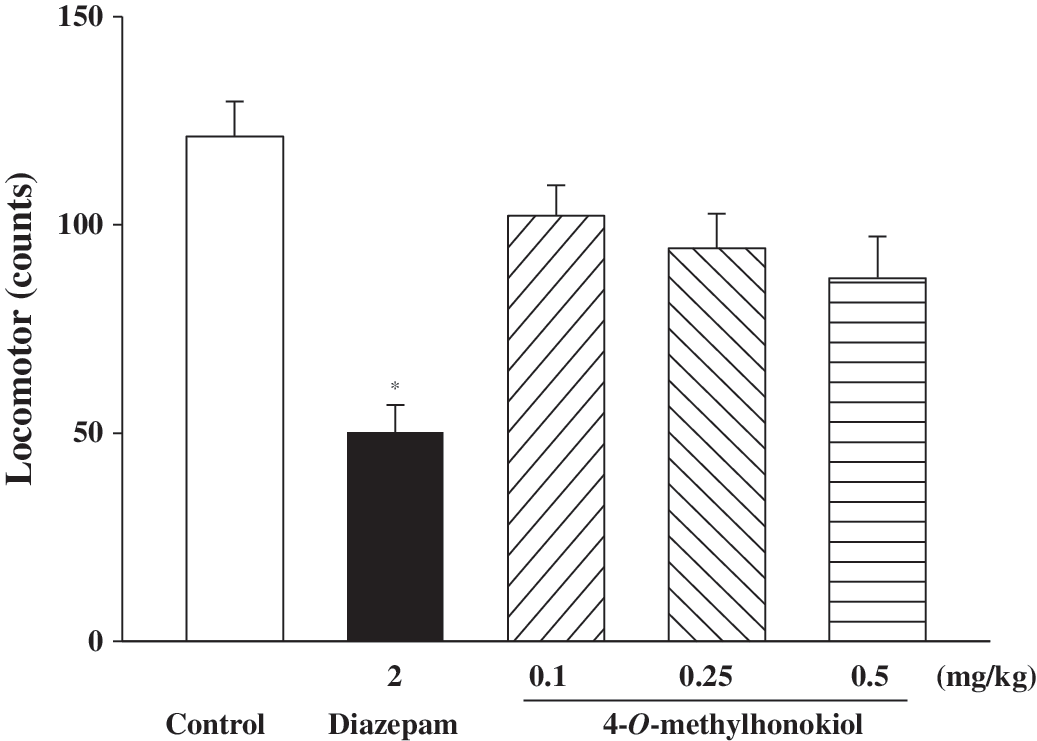
Effects of diazepam and 4-O-methylhonokiol on spontaneous locomotor activity. Diazepam or 4-O-methylhonokiol was orally administered 30 or 60 minutes, respectively, prior to the experiment. Ambulation activity was measured for 30 minutes. Data are mean±SEM values from 10 mice. *P<.05 compared with the control group.
Effects of 4-O-methylhonokiol on intracellular chloride influx
Becausee one of the major mechanisms of the anxiolytic effects of drugs could be related to the increase in Cl– influx, we investigated whether the anxiolytic-like effects of 4-O-methylhonokiol are due to the increase of Cl– influx in primary cultured brain cells. The resting intracellular Cl– concentration in cultured neurons was 27.3 mM, and treatment of primary cultured brain cells with 4-O-methylhonokiol at 2, 5, and 10 μM increased chloride influx to 29.6, 33.4, and 34.0 mM, respectively. Diazepam at 5 μM increased the chloride influx to 39.3 mM, compared with the control group (Fig. 6).

Effects of 4-O-methylhonokiol on chloride influx in primary cultured brain cells. Data are mean±SEM values from three separate experiments. *P<.05 compared with the control group.
Effect of 4-O-methylhonokiol on expression of GABAA receptor subunits
GABAA receptor subunit expressions were examined in cortex and hippocampus of mice brain after the mice were administered 4-O-methylhonokiol at the dosage of 0.5 mg/kg or diazepam at 2 mg/kg. 4-O-Methylhonokiol and diazepam increased the α-subunit expression significantly in both cortex and hippocampus areas (Fig. 7).
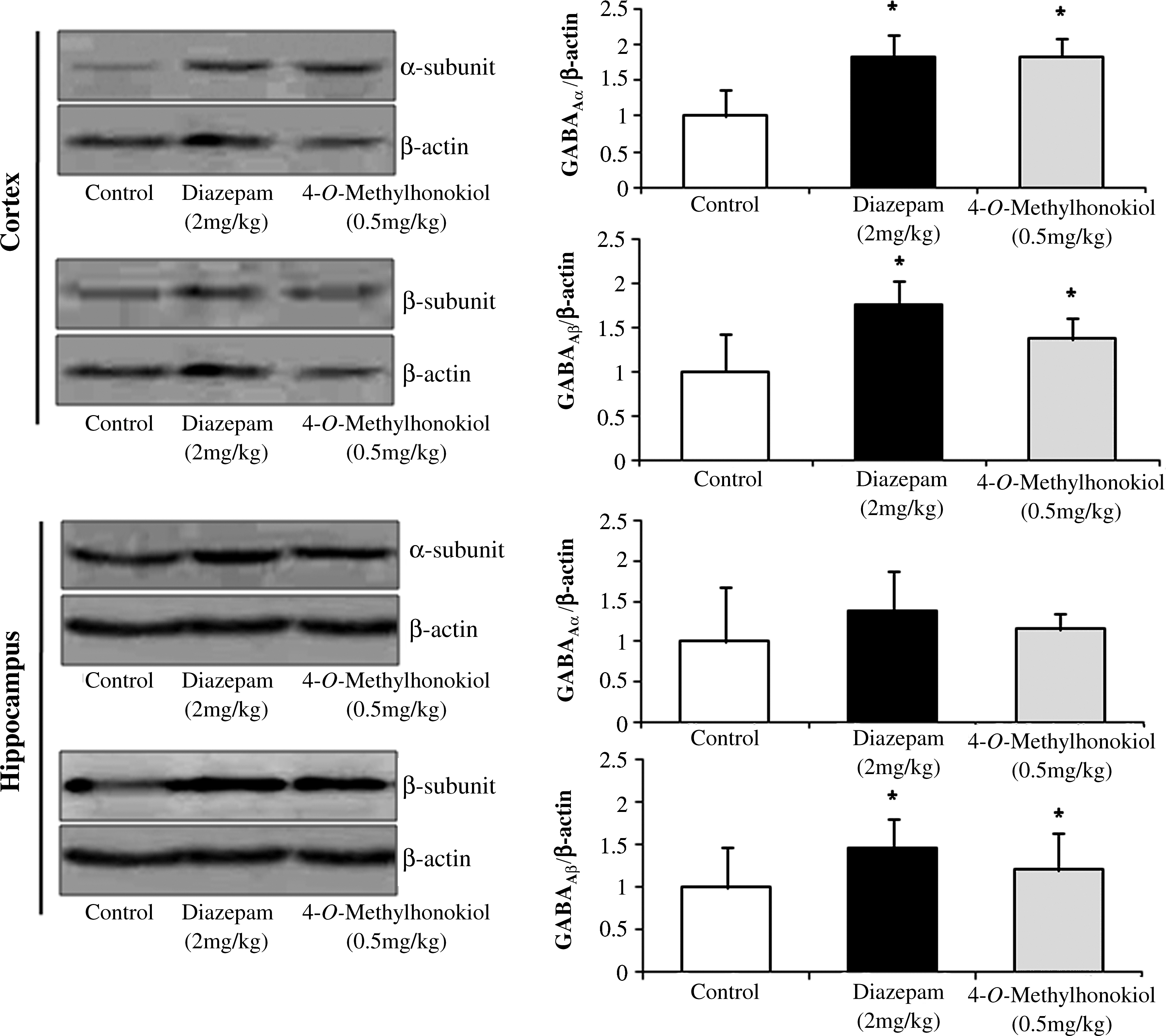
Effects of 4-O-methylhonokiol on γ-aminobutyric acid type A (GABAA) receptor expression. β-Actin was used as an expression control. Data are mean±SEM values from brains of three mice. *P<.05 compared with the control group.
Discussion
Natural remedies and herbal products are commonly used as putative treatments for anxiety and depression. Plants of the Magnoliaceae family have been empirically used for the treatment of various disorders, including anxiety and depression. 20,33 Recently, neolignan compounds such as magnolol and honokinol from M. obovata were isolated, and these compounds have been found to have anxiolytic effects. 33 We also reported that obovatol has anxiolytic effects. 20 This experiment was performed to investigate whether 4-O-methylhonokiol, another major neolignan compound isolated from the bark of M. officinalis, has anxiolytic-like effects and whether it affects expression of GABA/benzodiazepine receptors and Cl– channel activation as possible mechanisms. Diazepam has been used as a standard anxiolytic and also has been frequently used in behavioral pharmacology as a reference compound of potentially anxiolytic-acting substances. 34 –36
In this study, 4-O-methylhonokiol showed anxiolytic effects in animal models of anxiety, such as the elevated plus-maze and hole-board test. 27,37 One of the most widely used animal models for screening putative anxiolytics is the elevated plus-maze, 25,28 in which rodents show an avoidance of exposed open areas of the maze. 38 The anxiolytic effectiveness of a drug is illustrated by a significant exposure to open arms in the elevated plus-maze. The hole-board test provides another simple method for measuring the response of an animal to an unfamiliar environment and is widely used to assess emotionality, anxiety, and/or responses to stress in animals. 39 Head-dipping behavior is sensitive to changes in the emotional state of the animal, suggesting that the expression of an anxiolytic state in animals may be reflected by an increase in head-dipping behavior. 27 In the present study, oral administration of 4-O-methylhonokiol increased the percentages of entries into open arms and time spent in open arms, as well as increased head dips in the hole-board test, which indicated anxiolytic-like effects. The effects of 4-O-methylhonokiol seem to be similar or slightly less potent than those of diazepam, when the same doses of drugs are simply compared. However, 4-O-methylhonokiol may not cause side effects such as sedation, muscle relaxation, amnesia, and dependence as seen with the general benzodiazepines. 5 In other studies, we also observed that >100 mg/kg 4-O-methylhonokiol did not induce general toxicity by oral administration, but can cross the blood–brain barrier (data not shown).
It is well known that benzodiazepines facilitate the ability of GABA to activate the GABA receptor intrinsic Cl– channels and in turn facilitate inhibitory neurotransmission. This is manifest as an increase in the frequency of the ion channel opening in response to GABA. 4-O-Methylhonokiol increased the chloride influx significantly, compared with that of the control group. It is generally accepted that GABA receptor agonists decrease anxiety by facilitating the neuronal influx of chloride. Therefore, it is possible that anxiolytic-like effects of 4-O-methylhonokiol might be mediated by GABA/benzodiazepine receptor-activated Cl– channel opening. This effect is similar to that of obovatol 18 as well as diazepam. GABAA receptors are heteromeric ligand-gated chloride ion channels that are activated by brief releases of GABA into the synaptic cleft. GABA released from presynaptic interneuronal terminals binds to postsynaptic GABA receptors that gate a Cl– flux into the postsynaptic cell membrane. This ionic flux typically hyperpolarizes the postsynaptic neuronal membrane, making it less likely that arrival of excitatory neurotransmitters will be able to depolarize the postsynaptic cell sufficiently above the threshold for action potential initiation. The traditional hypnotics and anxiolytic drugs, such as barbiturates and benzodiazepines, enhance GABA transmission via the GABAA receptor. The α and γ subunits provide the binding site for the GABA molecule, while α and γ subunits are sensitive to benzodiazepine binding. 12,40 We found that 4-O-methylhonokiol increased the abundance of GABAA receptor α-subunit in the brain, indicating that 4-O-methylhonokiol increased GABA receptor to benzodiazepine binding, which activates the chloride channel by influencing the expression of GABA receptor subunit compositions.
To summarize, all the data presented here indicate that 4-O-methylhonokiol induced anxiolytic-like effects in the plus-maze test and hole-board test, and these pharmacological effects may be mediated by GABA/benzodiazepine receptor-activated Cl– channel opening and increasing availability of GABA receptors to benzodiazepine or GABA by influencing GABA receptor subunit compositions. In conclusion, 4-O-methylhonokiol might be a safe and effective treatment for anxiety.
Footnotes
Acknowledgment
This work was supported by the Priority Research Centers Program through the National Research Foundation of Korea funded by the Ministry of Education, Science and Technology (2009-0094035).
Author Disclosure Statement
No competing financial interests exist.