
Abstract
The fruit of Emblica officinalis has been used in the Ayurvedic system of medicine for the treatment of different ailments and is also an ingredient of various traditional medicinal herbal formulations in India and other countries. To investigate the protective effect of Emblica officinalis fruit extract (EFE) against alcohol-induced brain mitochondrial dysfunction, male Wistar rats were orally administered 20% alcohol (5 g/kg of body weight/day) and EFE (250 mg/kg of body weight/day) for 60 days. Alcohol-treated rats showed significantly lowered activities of mitochondrial antioxidant enzymes (superoxide dismutase and glutathione peroxidase) and reduced glutathione compared with those of experimental control rats. Furthermore, alcohol feeding lowered the activities of NADH dehydrogenase, succinate dehydrogenase (SDH), and cytochrome c oxidase and the content of cytochromes followed by increased levels of nitric oxide (NO), thiobarbituric acid-reactive substances, and protein carbonyls. No significant change was observed in membrane potential. Administration of EFE to alcohol-treated rats, lowered the levels of NO, protein carbonyls, and lipid peroxidation and elevated the activities of the antioxidant enzymes SDH, NADH dehydrogenase, and cytochrome c oxidase and the content of cytochromes. The active tannoid principles present in EFE with its antioxidant as well as NO scavenging properties might have contributed to the observed protection against alcohol-induced brain mitochondrial dysfunction.
Introduction
There is worldwide interest in the use of native plant extracts or natural products rather than synthetic drugs. 11 The phytochemicals from traditional medicinal plants have been presenting an exciting opportunity for the development of new therapeutics with novel beneficiary effects in combating alcoholism and related complications without causing side effects. 12,13 Earlier studies found that compounds in their natural formulations are more active than in their isolated form. 14
Emblica officinalis Gaertn. (commonly known in India as Amla, syn. Phyllanthus emblica L.; Family Euphorbiaceae) is regarded as “one of the best rejuvenating herbs” in Ayurveda, an Indian traditional science. 15 The fruits of Emblica are reported to contain hydrolyzable tannins, emblicanin A (β-glucogallin) and B (mucic acid 1,4-lactone 5-O-gallate), punigluconin, pedunculagin, gallic acid, ellagic acid, and rutin. 16,17 The fruit extract shows many pharmacological activities for the treatment of a number of diseases. Previous studies reported that Emblica fruit extract possesses hepatoprotective, 18 antidiabetic, 19 anticancer, 20 anti-atherogenic, 21,22 immunomodulatory, 23 and antioxidant 24,25 properties. Because inhibition of NO synthase activity and NO formation can be used as a potential therapeutic target, NO radical scavenging principles present in Emblica may be used for the treatment of various disease conditions. 26 Despite its extensive medicinal use, no information related to its use on alcohol toxicity is available. Hence, the present work was planned to study the protective effect of Emblica fruit extract against alcohol-induced brain mitochondrial dysfunction.
Materials and Methods
Chemicals
1-Anilino-8-naphthalene sulfonate, reduced glutathione (GSH), nitro blue tetrazolium, and NADH were purchased from Sigma Chemical Co. (St. Louis, MO, USA). All other chemicals were analytical grade and obtained from SISCO Research Laboratories (Mumbai, India).
Preparation of aqueous extract
Aqueous E. officinalis fruit extract (EFE), a dark-brown dry powder, was obtained from Chemiloids Ltd. (Vijayawada, India), a manufacturer and exporter of herbal extracts. Phytochemical analysis of extract showed the presence of flavonoids and phenols (90.8% water-soluble extractives, including 49.5% tannins). A 5% solution was prepared in water prior to administration.
Animals and experimental design
Two-month-old male albino Wistar rats, weighing about 120–140 g, were maintained in our animal house. They were fed with commercial pellet diet (Hindustan Lever Ltd., Bangalore, India) and tap water ad libitum. The animals were divided into four groups of eight rats each: group I control rats, which received glucose (i.e., caloric equivalent to 5 g of alcohol), group II alcohol-treated rats, which received 20% (vol/vol) alcohol in water (5 g/kg of body weight/day), group III rats, which received aqueous EFE (250 mg/kg of body weight/day) and after 8 hours 20% (vol/vol) alcohol in water (5 g/kg of body weight/day); and group IV rats, which received aqueous EFE (250 mg/kg of body weight/day) alone. The dose of the EFE in the present study was fixed at 250 mg/kg of body weight/day, based on our earlier studies. 27 All the groups received the treatment for 60 days by oral administration with the help of a stomach tube. The research was approved by the Sri Krishnadevaraya University Institutional Animal Ethics Committee. At the end of the experimental period, the rats in each group were fasted overnight and then sacrificed by cervical dislocation. Brain tissues were collected and processed immediately for further analysis.
Isolation of mitochondria
Mitochondria were isolated from brain as described previously with slight modifications. 28 In brief, brain was removed quickly and placed in ice-cold medium A (75 mM sucrose, 225 mM mannitol, 10 mM HEPES, 0.1 mM EDTA, and 0.1% bovine serum albumin, pH-adjusted to 7.4). A 10% homogenate was prepared using a Potter-Elevehjem homogenizer with a Teflon® (Dupont, Wilmington, DE, USA) pestle, by three or four up-and-down strokes for complete homogenization. The homogenate was centrifuged at 700 g for 10 minutes. The pellet was discarded, and the supernatant was centrifuged at 7,000 g for 10 minutes. The supernatant was discarded, and the pellet containing mitochondria was suspended in medium B (75 mM sucrose, 225 mM mannitol, 10 mM HEPES, and 0.1 mM EDTA, pH-adjusted to 7.4) by gentle homogenization by hand using a pestle and centrifuged at 7,000 g. This step was repeated to wash the mitochondria. The final pellet was suspended in medium B to a protein concentration of 15–20 mg of protein/mL and immediately used for studies.
Determination of thiobarbituric acid-reactive substances and protein carbonyls
Thiobarbituric acid-reactive substances (TBARS) levels were measured as described previously using malondialdehyde as the standard and expressed as nmol of malondialdehyde/mg of protein. 29 The concentration of protein carbonyls was determined using 2,4-dinitrophenylhydrazine. Protein carbonyl content was calculated using the molar extinction coefficient of 22,000 and expressed as nmol of carbonyls/mg of protein. 30
Determination of antioxidant status
Manganese superoxide dismutase (Mn-SOD) was assayed; a single unit of enzyme was expressed as 50% inhibition of nitro blue tetrazolium reduction/minute/mg of protein. 31 Glutathione peroxidase (GPx) activity 32 and total GSH content were measured. 33
Nitrite/nitrate analyses
Nitrite/nitrate levels were determined as described previously. 34 In brief, samples were treated with 30% zinc sulfate to deproteinize before centrifugation at 4,000 g for 5 minutes. Nitrite was determined from 0.5-mL aliquots of supernatant by diazotization using Griess reagent (1% sulfanilamide, 2.5% phosphoric acid, and 0.1% 1-naphthyl-ethylenediamine). For nitrate estimation, sample was swirled for 90 minutes separately with activated cadmium filings for the conversion of nitrate to nitrite, and then Griess reagent was added. Nitrite concentrations were estimated using a standard curve developed with sodium nitrite.
Determination of respiratory enzyme activities and membrane potential
NADH dehydrogenase activity was measured spectrophotometrically, using potassium ferricyanide as the electron acceptor. 35 The assay system contained 20 μg of mitochondrial protein, 250 mM sucrose, 30 mM Tris-HCl, 10 mM potassium phosphate, 5 mM magnesium chloride, 1 mM potassium cyanide, 1 mM potassium ferricyanide, and 1 mM NADH, pH 7.4. Activity was expressed as nmol of potassium ferricyanide reduced/minute/mg of protein. The activity of succinate dehydrogenase (SDH) was assayed using 2,6-dichlorophenolindophenol as the electron acceptor. 36 The reaction system was the same as used for NADH dehydrogenase except that potassium ferricyanide was replaced with 1 mM phenazine methosulfate and 70 μM 2,6-dichlorophenolindophenol. The rate of the reduction of 2,6-dichlorophenolindophenol was followed at 600 nm. Ten micrograms of mitochondrial protein was incubated with 10 μL of 0.5 M sodium succinate (pH 7.4) at room temperature for 10 minutes before assaying SDH activity. The SDH activity was calculated based on the millimolar extinction coefficient (E = 16.9 mM). Cytochrome c oxidase activity was measured spectrophotometrically by following the decrease in the absorbance of ferrocytochrome c at 550 nm. 37 Activity was expressed as nmol of cytochrome c oxidized/minute/mg of protein. Membrane potential (ΔΨ) was determined by the distribution of the fluorescent probe 1-anilino-8-naphthalene sulfonate across the membrane. Succinate was used to polarize and carbonyl cyanide m-chlorophenyl hydrazone to depolarize the membrane. The changes of 1-anilino-8-naphthalene sulfonate fluorescence in the energized and deenergized state were used for the calculation of ΔΨ. Membrane potential was calculated using Nernst's equation and expressed in mV. 38
Estimation of cytochromes
Cytochromes b, c, and a a 3 concentrations were determined as described previously. 39 In brief, mitochondrial suspension (1.0 mL containing 4 mg of protein) was mixed with 0.4 mL of potassium phosphate buffer (100 mM), 0.2 mL of potassium ferricyanide (15 mM), and 0.2 mL of deoxycholate (0.5 mg/mg of protein), pH 7.4. After thorough mixing, the suspension (containing the cytochromes in oxidized form) was divided into two equal parts and transferred to reference and sample cuvettes. The difference spectrum was recorded, which serves as baseline. Then the contents of the sample cuvette were reduced with a few grains (∼50 mg) of sodium dithionite. Reduced minus oxidized difference spectrum was recorded from 500 to 650 nm. The concentration of cytochrome b was determined by reducing the contents of the reference cuvette by adding 0.1 mL of sodium ascorbate (3 mM) and 200 mM N,N,N′,N′-tetramethyl-p-phenylenediamine. The difference spectrum of ascorbate plus N,N,N′,N′-tetramethyl-p-phenylenediamine reduced versus sodium dithionite reduced was recorded from 500 to 600 nm. Millimolar extinction coefficients used were 19.1, 20, and 16 for cytochrome c (540–550 nm), cytochrome b (560–575 nm), and cytochrome a a3 (605–630 nm), respectively. Protein content in mitochondria was estimated as previously described. 40
Statistical analysis
All results were expressed as mean ± SD values of eight rats in each group. The data were subjected to analysis of variance followed by Duncan's multiple range test. A value of P < .05 was considered statistically significant.
Results
Figure 1 shows the extent of lipid peroxidation in brain mitochondria of control and experimental animals. The concentration of TBARS was significantly (P < .05) higher in alcohol-administered rats than in the control rats. Administration of EFE and alcohol together reduced the concentration of TBARS significantly (P < .05) compared with those of the alcohol-administered rats. EFE administration to control rats did not produce any significant change in the TBARS concentration.
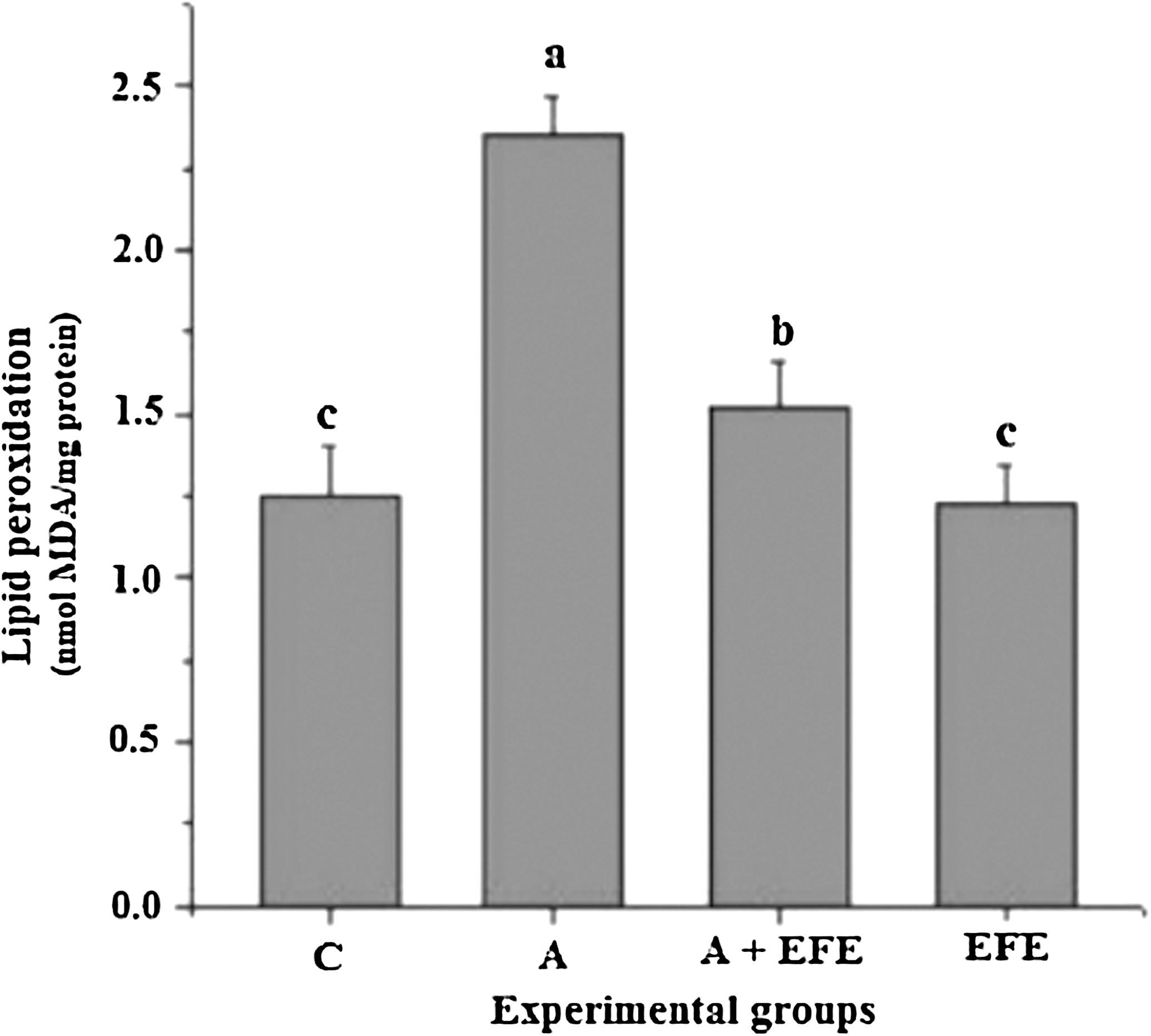
Effect of E. officinalis fruit extract (EFE) administration on brain mitochondrial thiobarbituric acid-reactive substances in alcohol-treated rats. Data are mean ± SD values of eight rats in each group. abcMeans sharing a common superscript are not significantly different (P < .05) among groups. A, alcohol alone-treated rats; A + EFE, alcohol- and EFE-treated rats; C, control (glucose-treated) rats; MDA, malondialdehyde.
The carbonyl concentration was significantly (P < .05) higher in brain mitochondria of alcohol-administered rats than in experimental control rats (Fig. 2). Treatment with EFE alone did not alter the brain mitochondrial carbonyl content. Administration of EFE along with alcohol resulted in significantly lower carbonyl content (P < .05) compared with alcohol alone-administered rats.
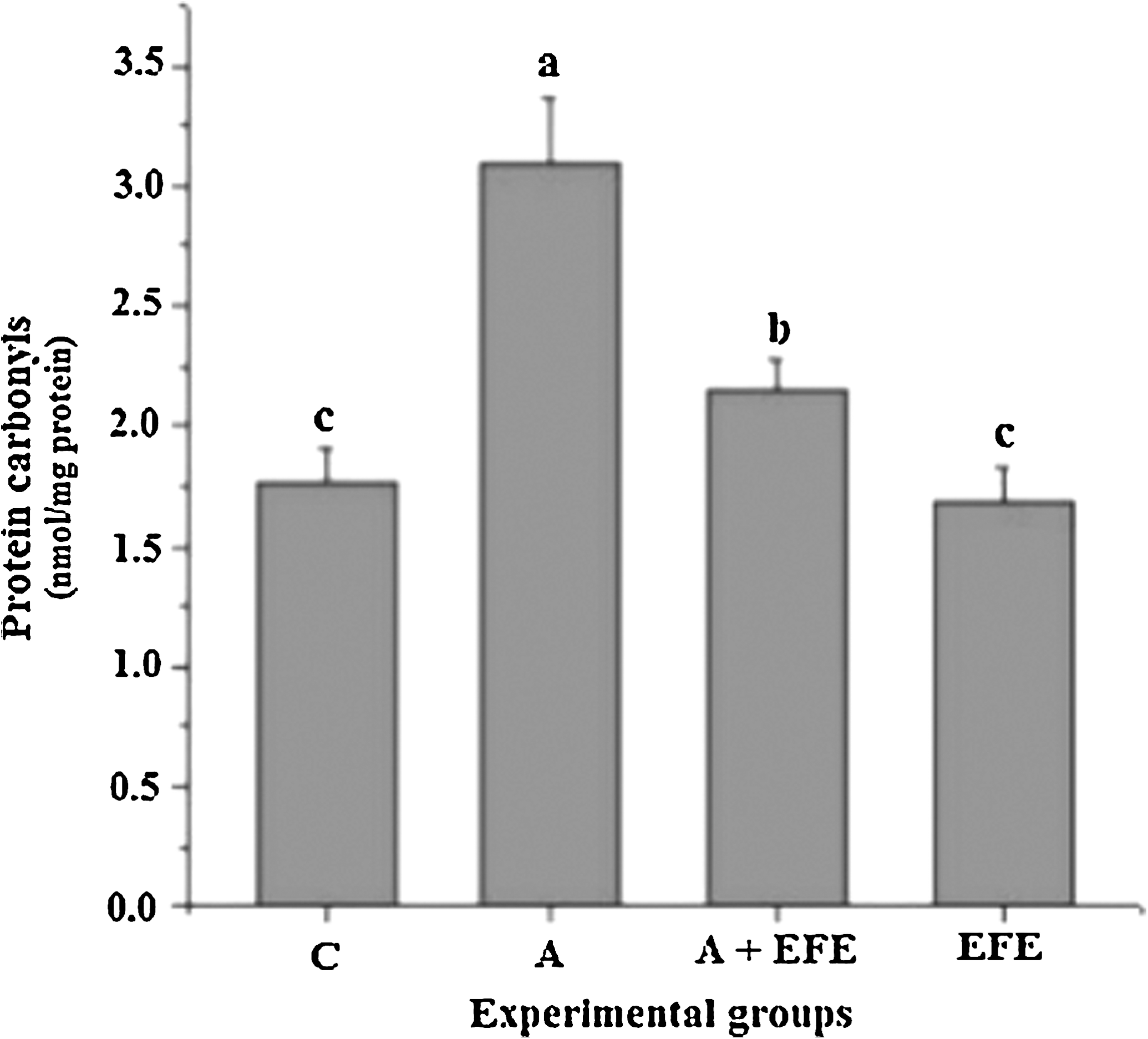
Effect of EFE administration on brain mitochondrial protein carbonyl content in alcohol-treated rats. Data are mean ± SD values of eight rats in each group. abcMeans sharing a common superscript are not significantly different (P < .05) among groups.
Nitrite/nitrate levels are reliable indicators of NO production. The concentrations of brain mitochondria nitrite/nitrate levels of control and experimental animals are shown in Figure 3. Administration of EFE to control rats did not significantly alter the nitrite/nitrate levels, whereas nitrite/nitrate levels were significantly (P < .05) elevated in the alcohol-treated rats, suggesting overproduction of NO compared to the other groups. EFE administration significantly decreased nitrite/nitrate levels in rats that received alcohol.
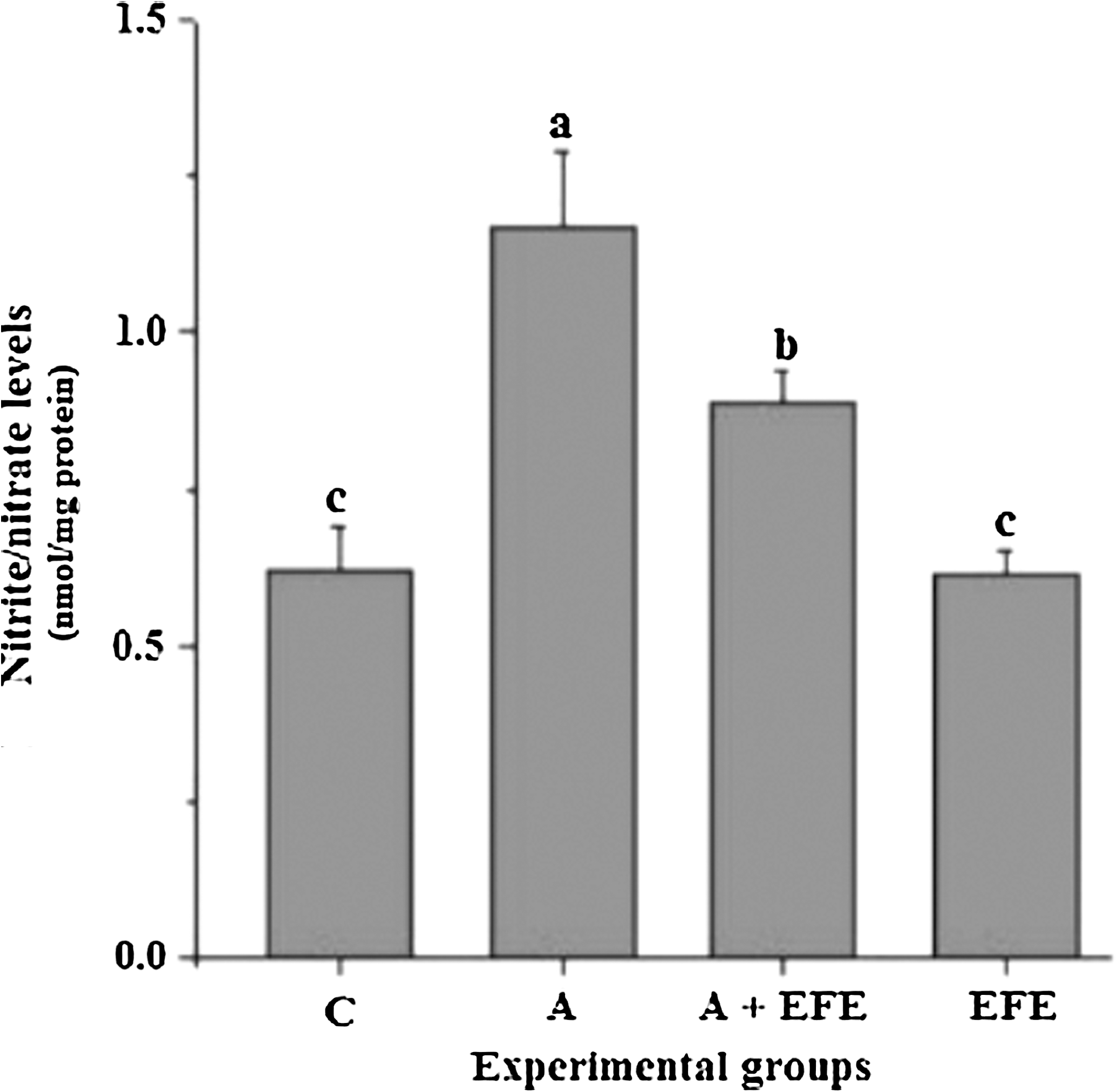
Effect of EFE administration on brain mitochondrial nitric oxide production in alcohol treated rats. Data are mean ± SD values of eight rats in each group. abcMeans sharing a common superscript are not significantly different (P < .05) among groups.
The activities of brain mitochondrial antioxidant enzymes are shown in Table 1. Alcohol administration significantly (P < .05) decreased the activities of SOD and GPx with a fall in GSH content compared to controls. Administration of EFE to alcohol-administered rats significantly (P < .05) increased the activities of these enzymes and GSH content to normal levels. No significant (P < .05) changes were observed in EFE alone-administered rats.
Data are mean ± SD values of eight rats in each group.
Means in the same column not sharing a common superscript are significantly different (P < .05) between groups.
GPx, glutathione peroxidase; GSH, reduced glutathione; Mn-SOD, manganese-superoxide dismutase.
There was a significant (P < .05) decrease in the activities of SDH, NADH dehydrogenase, and cytochrome c oxidase with no significant change in ΔΨ due to alcohol administration (Table 2). Treatment with EFE along with alcohol significantly (P < .05) prevented the alcohol-induced inhibition of SDH, NADH dehydrogenase, and cytochrome oxidase activities. EFE relieved the inhibition caused by ethanol almost completely for NADH dehydrogenase and to some extent for cytochrome oxidase. The administration of EFE alone did not have any significant effect on these enzymes.
Data are mean ± SD values of eight rats in each group.
Means in the same row not sharing a common superscript are significantly different (P < .05) between groups.
SDH, succinate dehydrogenase; ΔΨ, membrane potential.
The amounts of cytochromes a a 3, b, and c were decreased significantly (P < .05) in alcohol-administered rats compared to controls (Table 3). Simultaneous administration of EFE and alcohol augmented the cytochrome contents of the mitochondria, but not back to the control levels. The maximum relief was seen for cytochrome a a 3 and then for cytochrome b. However, administration of the EFE alone did not result in any significant (P < .05) changes in the cytochrome contents compared with control rats.
Data are mean ± SD values of eight rats per group. Cytochrome a a3 , b, and c values are expressed as μmol/minute/mg of protein.
Means in the same column not sharing a common superscript are significantly different (P < .05).
Discussion
E. officinalis is known for its therapeutic properties. In the present study we tested the therapeutic efficacy of Emblica extract against alcohol-induced brain mitochondrial dysfunction. Chronic alcohol treatment affects mitochondrial function, including oxygen uptake at high blood alcohol levels. Recent studies have suggested that ethanol impairs mitochondrial function by increasing the production of ROS and causes cellular toxicity. 41 ROS-mediated toxicity has been considered a key underlying mechanism responsible for ethanol-induced brain injury. 42 We observed increased mitochondrial protein carbonyls and lipid peroxidation in alcohol-treated rats. Protein and lipid oxidation by ROS has to be considered one of several ethanol-related modifications especially within the mitochondria because ethanol increases ROS within this organelle. 43 In the present study, alcohol consumption increased NO levels, which may further lead to toxicity by formation of peroxynitrite (ONOO−), a potent oxidant that produces nitration of tyrosine and inactivation of biologically important proteins and enzymes. 44 Basal levels of NO produced by constitutively active NO synthase are protective, whereas high NO concentrations generated by inducible NO synthase may be harmful. The low concentration of NO diminishes metal-catalyzed lipid peroxidation, whereas the high concentration of NO potentiates oxidative stress and causes mitochondrial dysfunction. The protection is due to up-regulation of protective proteins (heme oxygenase and ferritin). 45 Inhibition of NO synthase and selective scavengers of peroxynitrite exert beneficial effects in various animal models. 46 Earlier reports revealed that Emblica inhibits inducible NO synthase enzyme activity in aging. 47 Administration of EFE to rats receiving alcohol significantly decreased lipid peroxidation and protein carbonyls to normal levels and NO to moderate levels. The protective effect of EFE against alcohol-induced oxidative damage is evident from decreased lipid and protein oxidation.
Brain tissue is particularly vulnerable to oxidative damage, possibly because of its high consumption of oxygen, low levels of antioxidant enzymes, high levels of free iron, and the consequent generation of high quantities of ROS during oxidative phosphorylation. 48 Generation of oxygen metabolites such as superoxide, hydrogen peroxide, and hydroxyl radicals is believed to be important in the pathogenesis of alcoholic brain injury. 49 To counteract these oxidants mitochondria have several antioxidant enzymes including Mn-SOD and GPx. Hydrogen peroxide can be readily converted to water by mitochondrial GPx, which oxidizes GSH to oxidized glutathione. Glutathione reductase then converts oxidized glutathione back to GSH. However, electron leak to oxygen through complexes I and III can generate superoxide anion (O2 •−), which can be converted to hydrogen peroxide by the mitochondrial matrix enzyme Mn-SOD. However, in the presence of reduced transitional metals, hydrogen peroxide can be converted to the highly reactive hydroxyl radical (OH•), which may further react with NO radical (NO•) to generate the highly reactive peroxynitrite. 6 Mitochondria undergo oxidative damage when ROS production exceeds the antioxidant capacity of mitochondria. In the present study, decreased mitochondrial SOD and GPx activities followed by a drop in GSH content were observed in alcohol-treated rats compared to control rats. Many antioxidants are unable to penetrate the blood–brain barrier and may not reach the relevant sites of free radical generation, making the brain more vulnerable to oxidative damage. Large proteins like Mn-SOD and catalase do not penetrate cell membranes and are therefore ineffective against intracellular ROS. 50 Administration of EFE to alcohol-treated rats significantly increased antioxidant enzyme activities. The efficacy of EFE essentially depends on the active constituents such as phyllembic acid, ellagic acid, and gallic acid, reported to be powerful antioxidants. 24,51 Moreover, the hydrolyzable low-molecular-weight tannoid principles β-glucogallin, mucic acid 1,4-lactone 5-O-gallate, punigluconin, pedunculagin, gallic acid, ellagic acid, and rutin of EFE may have the ability to penetrate the blood–brain barrier, enabling them to contribute to the enhanced antioxidant status against ROS production in mitochondria.
Oxidation and nitration of proteins alter function of many metabolic enzymes in the mitochondrial electron transport chain, including NADH dehydrogenase, SDH, and cytochrome c oxidase. 52 In the present study NADH dehydrogenase, SDH, and cytochrome c oxidase activities were decreased significantly with no significant change in ΔΨ in alcohol-administered rats. NO through its interactions with components of the mitochondrial respiratory chain may function as a physiological regulator of cell respiration and a modulator of the generation of ROS by mitochondria, thereby affecting mechanisms of cell survival or death. 53,54 In addition, NO irreversibly inhibits cytochrome c oxidase activity. 55 Ethanol-induced overproduction of NO leads to secondary production of reactive nitrogen species, such as peroxynitrite, leading to irreversible modification of respiratory chain proteins and an increased sensitivity to NO. 4 Our results showed that administration of EFE to alcohol-treated rats restored the activities of all respiratory enzymes to some extent compared to alcohol alone-administered rats. Cytochromes play a crucial role as electron carriers. When there is a significant decrease in the concentration of these cytochromes (b, c, and a a 3), the rate of electron transfer also decreases significantly. 38 In the alcohol-treated group, brain mitochondrial cytochrome a a 3, b, and c contents were significantly decreased compared to controls. Furthermore, the cytochrome levels were significantly restored in EFE-administered rats receiving alcohol.
Six compounds of Emblica were reported to have NO scavenging effects: geraniin, corilagin, furosin, gallic acid, methyl gallate, and curcumin. 26 In the present study, ethanol-induced overproduction of NO was decreased by the administration of EFE. Modulation of this process may prevent or at least attenuate alcoholism-induced changes in the brain. These findings suggested EEE could alter the activities of mitochondrial respiratory enzymes. In addition to NO scavenging compounds, free hydroxyl groups of phenolic compounds present in EFE are also responsible for the antioxidant and ROS/reactive nitrogen species scavenging activity. In conclusion, this study clearly showed that EFE offers protection against alcohol-induced brain mitochondrial dysfunction, perhaps by inhibiting inducible NO synthase enzyme activity and/or by scavenging NO. Furthermore, low-molecular-weight hydrolyzable tannoid compounds may cross the cell membranes to improve antioxidant status by scavenging free radicals. The molecular interactions between tannoid compounds and free radicals need further in-depth study.
Footnotes
Acknowledgment
This study was supported in part by the University Grants Commission (grant F-3-11/97), New Delhi, India.
Author Disclosure Statement
No competing financial interests exist.