
Abstract
Resveratrol has been shown to possess anticancer, anti-aging, anti-inflammatory, antimicrobial, and neuroprotective activities. In this study, we examined the antiproliferative properties of resveratrol and its molecular mechanism(s) of action in Huh-7 cells, a new human hepatoma cell line system for hepatitis C virus. Results showed that resveratrol significantly inhibited Huh-7 cell proliferation (50% inhibitory concentration = 22.4 μg/mL) and effectively induced cell cycle arrest and apoptosis. It up-regulated p21/WAF1 expression in a p53-independent manner, but the expressions of cyclin E, cyclin A, and cyclin-dependent kinase 2 were down-regulated. It also caused an increase in the ratio of pro-apoptotic/anti-apoptotic protein, which was associated with the mitochondrial membrane depolarization and the increase in caspase activity. Resveratrol showed no effect on Fas, Fas ligand, extracellular signal regulated kinase (ERK) 1/2, and p38 expression but down-regulated phospho-ERK and phospho-p38 expression. In addition, resveratrol was noted to trigger autophagic cell death through the increased expression of autophagy-related Atg5, Atg7, Atg9, and Atg12 proteins. These results suggest that resveratrol could be an important chemoprevention agent for hepatoma of hepatitis C virus infection.
Introduction
H
Increasing evidence suggests that diets rich in fruits and vegetables can decrease the risk of malignancies. Studies have shown that antioxidant vitamins and phytochemicals of fruits and vegetables are promising chemopreventive agents. 3 –6 Some of these dietary chemopreventive phytochemicals have also been reported to possess excellent chemotherapeutic potentials.
Apoptosis, or programmed cell death, plays an essential role in all multicellular organisms. It is characterized by chromatin condensation, nuclear breakdown, DNA fragmentation, and activation of the caspases. 7 Caspase activation occurs via cascade signaling events, in which the activation of initiator caspases would lead to the activation of downstream executive caspases, 8 –10 and ultimately triggers the apoptotic cell death program. 11
Autophagy (or macroautophagy) is characterized by the presence of cytoplasmic engulfing vesicles (autophagosomes) and has been known to play both pro-survival and pro-death roles. 12 It is regulated by the autophagy-related (Atg) genes that control the formation of autophagic vesicles. 13 Recent investigations have demonstrated that the co-regulation of both apoptosis and autophagy can participate in mammalian cell death. Some reports have further indicated that apoptosis and autophagy may be interconnected and even simultaneously regulated by the same signal. 14 –16 Although the mechanism linking apoptotic cell death and autophagic cell death under physiological conditions remains unclear, the modulation of autophagy has been suggested to be a novel strategy for cancer treatment.
Resveratrol (trans-3,4',5-trihydroxystilbene) (Fig. 1), one of the plant antibiotics and phytoalexins, is richly found in many plants such as grapes, berries, nuts, eucalyptus, and rheum. Its biological activities have been extensively studied, including antibacterial, 17 antifungal, 18 anti-aging, anti-inflammatory, and antiviral, 19 and neuroprotective 20,21 activities. Studies have also indicated that resveratrol possesses activities against human hepatoma Hep G2 and SK-HEP-1 22 –25 and cancers of breast, 26 lung, 27 leukemia, 22 bladder, 28 ovary, 29 and prostate. 30 It prevented the proliferation of tumor cells by inhibiting DNA synthesis 31 and cell cycle progression 32 and by modulating a series of signaling molecules. 28 However, no study has been reported on the effects of resveratrol on hepatoma of hepatitis C virus infection. 33 In this study, our aim was to examine the antiproliferative effect of resveratrol and its molecular mechanism(s) of apoptotic induction and autophagy in HCC Huh-7 cells, a commonly used cell culture system for the study of hepatitis C virus infection. 34,35
Structure of resveratrol.
Materials and Methods
Chemicals
Resveratrol, dimethyl sulfoxide (DMSO), sodium 3,3-[(phenylamino)carbonyl]-3,4-tetrazolium-bis(4-methoxy-6-nitro)benzenesulfonic acid hydrate, RNase, and propidium iodide were purchased from Sigma Chemical Co. (St. Louis, MO, USA). The p53, p21/WAF1, caspase inhibitors, and JC-1 mitochondrial membrane potential assay kits were obtained from Calbiochem (Cambridge, MA, USA). Caspase activity assay kits were obtained from Invitrogen (Carlsbad, CA, USA). Fas and Fas ligand (Fas L) activity assay kits were obtained from Bender (Vienna, Austria). Dulbecco's modified Eagle's medium and fetal bovine serum were purchased from Gibco BRL (Gaithersburg, MD, USA).
Antibodies
The antibodies for Bax (diluted 1:1,000), Bcl-2 (diluted 1:1,000), cyclin A (diluted 1:2,000), cyclin E (diluted 1:1,000), cyclin-dependent kinase (CDK) 2 (diluted 1:1,000), extracellular signal-regulated kinase (ERK) 1/2 (diluted 1:1,000), p38 (diluted 1:1,000), phospho-ERK1/2 (p-ERK1/2) (diluted 1:1,000), and phospho-p38 (p-p38) (diluted 1:1,000) were purchased from Cell Signaling (Beverly, MA, USA). The antibody for Bcl-xL (diluted 1:1,000) was purchased from Epitomics (Burlingame, CA, USA). The autophagy sampler kit was obtained from Abgent (San Diego, CA, USA). β-Actin (diluted 1:5,000) and anti-mouse and anti-rabbit secondary horseradish peroxidase conjugates were obtained from Chemicon (Billerica, MA, USA).
Cell culture
Human HCC Huh-7 cells were provided by C.D.R. and were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum. The cells were maintained under standard cell culture conditions at 37°C and 5% CO2 in a humidified incubator.
Preparation of test solution
Resveratrol was dissolved in DMSO. To ensure cell viability was not affected by DMSO, its final concentration used for all treatments was controlled at 0.1% (vol/vol).
Cell proliferation assay
Cells (1 × 105 cells per well) were seeded in 96-well plates. After 24 hours of incubation, seeded cells were treated with resveratrol (10, 20, 40, and 60 μg/mL) for 48 hours followed by 3,3-[(phenylamino)carbonyl]-3,4-tetrazolium-bis(4-methoxy-6-nitro)benzenesulfonic acid hydrate assay. The 50% inhibitory concentration (IC50) of resveratrol was calculated based on the values of absorbance measured at 48 hours after treatment.
Cell cycle analysis
Cell cycle distribution was determined by flow cytometry. A total of 5 × 105 cells per well were seeded in six-well plates and then treated with resveratrol (10, 20, and 40 μg/mL) or vehicle alone (0.1% DMSO) for 48 hours. After treatment, the cells were collected by trypsinization, washed twice with phosphate-buffered saline, and then fixed in methanol/phosphate-buffered saline (2:1 vol/vol) at 4°C overnight. After two washes with phosphate-buffered saline, cells were resuspended in 1 mL of phosphate-buffered saline containing 25 μg/mL RNase and 50 μg/mL propidium iodide, followed by incubating in the dark for 30 minutes. The measurement was performed by a FACSCalibur™ system from Becton-Dickinson (San Jose, CA, USA), and the data were analyzed by ModFit version 3.0.
Apoptosis assay
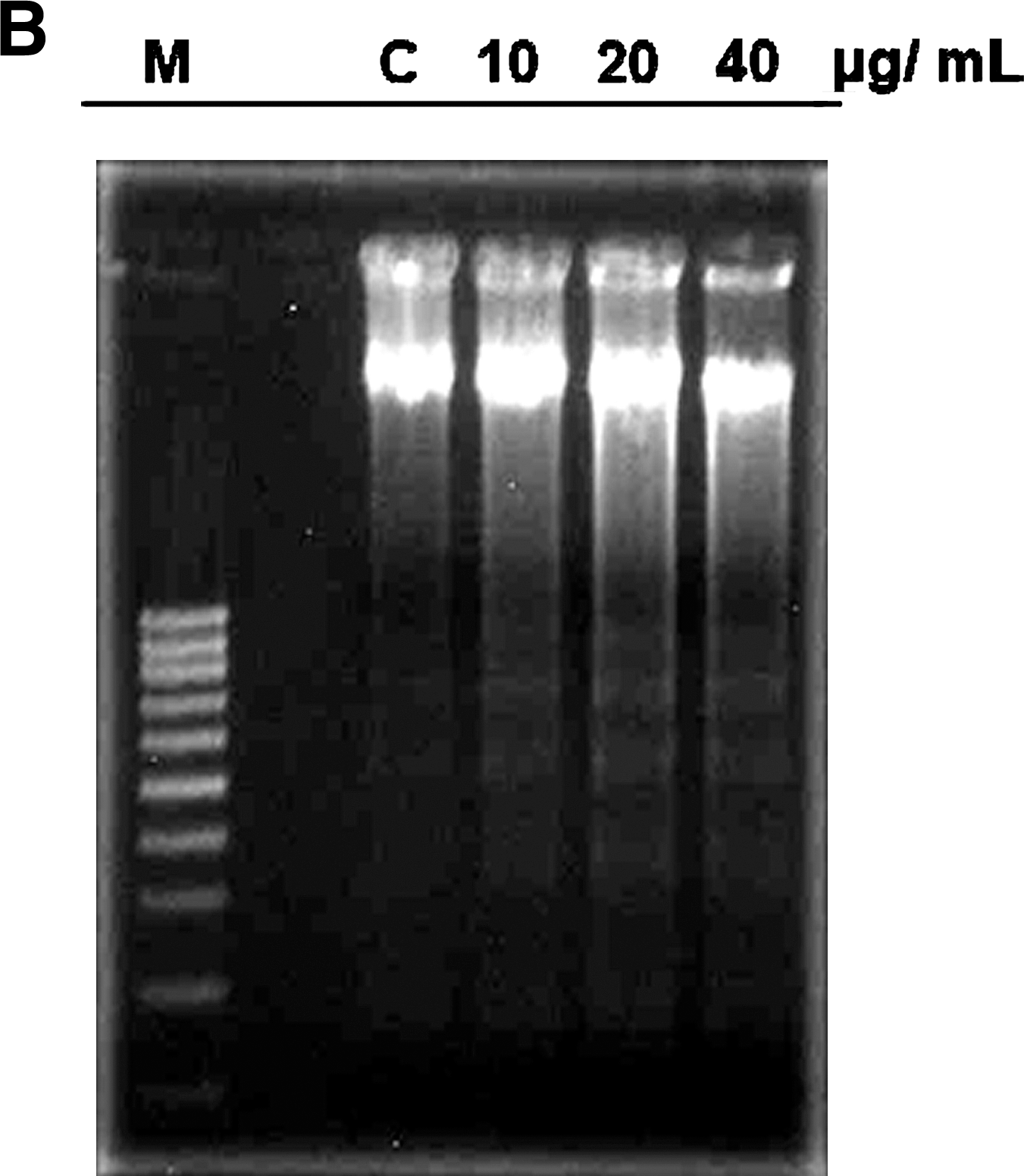
In brief, 1 × 106 cells were seeded in 100-mm-diameter dishes and treated with resveratrol (10, 20, and 40 μg/mL) or vehicle alone (1% DMSO) for 48 hours. After centrifugation, the cell pellets collected were lysed by nuclei lysis buffer, treated with RNase and protein precipitation solution, and further subjected to centrifugation. DNA was extracted with an equal volume of isopropanol and then separated in 1% agarose gel. After staining with ethidium bromide, the sample was visualized by ultraviolet light.
p53, p21/WAF1, Fas, and Fas L expression assay
Activities of p53, p21/WAF1, Fas, and Fas L were measured by enzyme-linked immunosorbent assay kits. In brief, cells were treated with 10 μg/mL resveratrol for various time periods (0, 6, 12, 24, and 48 hours). The cell lysates collected were placed in 96-well plates coated with the specific monoclonal antibodies and then incubated according to the manufacturer's instructions. After the unbound sample was removed, the sample was washed three times with washing buffer, and then streptavidin (horseradish peroxidase conjugated) was added to bind the respective antibodies. The absorbance was measured at 450 nm. The concentrations of p53, p21/WAF1, Fas, and Fas L were determined by their respective standard curves.
Mitochondrial membrane potential assay
For measurement of the mitochondrial membrane potential, the mitochondria-specific cationic dye JC-1 was used. In brief, cells were seeded in 96-well plates (1 × 104 cells per well) and treated with 10 μg/mL resveratrol for 0, 6, 12, 24, and 48 hours. Cells were stained with 10 μg/mL JC-1 for 30 minutes at 37°C. Fluorescence was monitored with a fluorescent plate reader at a wavelength of 490 nm (excitation)/540 nm (emission) and 490 nm (excitation)/590 nm (emission). The ratio of values at 590 nm to 540 nm indicates the changes in the mitochondrial membrane potential, whereby the mitochondrial depolarization is indicated by a decrease in the red/green fluorescent intensity ratio.
Caspase activity assay
Caspase activities were determined by the colorimetric assay using caspase 8 and caspase 9 activation kits and performed according to the manufacturer's protocols. Forty-eight hours after resveratrol treatment, cells were lysed by lysis buffer. The supernatants collected were incubated with the reaction buffer containing dithiothreitol and substrates at 37°C. The caspase activity was determined at a wavelength of 405 nm.
Western blotting analysis
To measure the expression of cyclins, CDK2, Bcl-2 family, ERK, p38, and Atg proteins, cells were treated with resveratrol at various concentrations and time periods and then lysed in lysis buffer, followed by centrifuging at 15,000 g (model 5417R centrifuge, Eppendorf AG, Hamburg, Germany) for 10 minutes to obtain the supernatants. Equal amount of proteins were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred to nitrocellulose membranes. The membranes were blocked in 5% nonfat milk for 1 hour at 37°C and then incubated with the specific primary antibody overnight at 4°C. Subsequently, the membranes were incubated with an appropriate amount of horseradish peroxidase-conjugated secondary antibody. The specific protein was detected by an ECL kit (Amersham Biosciences, Piscataway, NJ, USA).
Statistical analysis
Data were expressed as mean ± SD values from three independent experiments. Statistical comparisons of the results were done using analysis of variance. Significant differences (P < .05) between the means of control and resveratrol-treated cells were analyzed by Scheffé's test.
Results and Discussion
Resveratrol causes Huh-7 cell death
Figure 2 shows that treatment of Huh-7 cells with 10, 20, 40, and 60 μg/mL resveratrol resulted in a dose-dependent increase in cell death (32.7%, 49.9%, 72.9%, and 88.7%, respectively), and the IC50 value was estimated to be 22.4 μg/mL. Resveratrol significantly inhibited cell cycle progression (Fig. 3A) and caused a persistent accumulation of cells in S phase. It also caused the degradation of DNA with the formation of DNA fragments in a dose-dependent manner (Fig. 3B). Compared with the vehicle treatment, resveratrol showed a dose-dependent induction of Huh-7 cell apoptosis (Fig. 3C). These results suggest that resveratrol was effective in suppressing Huh-7 cell proliferation and inducing cell death through cell cycle arrest and apoptosis.
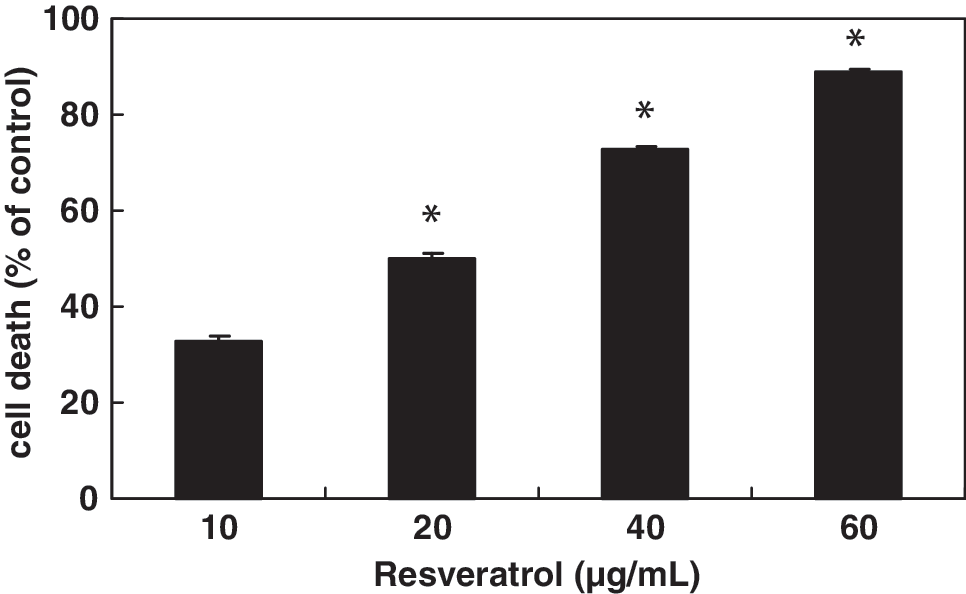
Effect of resveratrol on Huh-7 cell viability. Values indicating the cell number are expressed as percentages of the control cells. Data are mean ± SD values of triplicate experiments. *P < .05, significant difference between control and resveratrol-treated cells as analyzed by Scheffé's test.

Effect of resveratrol on cell cycle arrest and apoptosis in Huh-7 cells.

Resveratrol increases p21/WAF1 expression through a p53-independent pathway
Because resveratrol treatment was shown to arrest the cell cycle at S phase, we further assessed the effect of resveratrol on cell cycle-regulatory proteins, including p53, p21/WAF1, CDK2, cyclin A, and cyclin E. Results showed that treatment with 10 μg/mL resveratrol did not affect the expression of p53 at any time point (Fig. 4A). However, treatment with the same concentration of resveratrol was noted to trigger a time-dependent reduction in cyclin A, cyclin E, and CDK2 expression (Fig. 4B) but caused an increase in the expression of p21/WAF1 at 6, 12, 24, and 48 hours after treatment (Fig. 4C). Furthermore, resveratrol showed inhibitory effects on CDK2/cyclin A and CDK2/cyclin E complexes and induced the p21/WAF1 expression in a p53-independent manner. These observations suggest that resveratrol-induced apoptosis was not dependent on p53 but was mediated through an increase in p21/WAF1 expression to inhibit the cell cycle progression.

Effects of resveratrol on cell cycle-related proteins in Huh-7 cells:
Resveratrol-mediated apoptosis involves caspase 8 but not Fas/Fas L
In clarifying the mechanism of resveratrol-induced apoptosis in Huh-7 cells, we further examined the activity of the death receptor apoptotic pathway, specifically the Fas receptor and the Fas L. Results showed that resveratrol did not affect the expression of either Fas receptor or Fas L in Huh-7 cells (Fig. 5A and B). However, it did up-regulate the activity of caspase 8, the downstream effector of the mitochondria-independent apoptotic pathway (Fig. 5C). Pretreatment with the caspase 8 inhibitor IETD-fmk was shown to decrease the percentage of Huh-7 cell death induced by resveratrol (Fig. 6), suggesting that caspase 8 is involved in apoptotic cell death.
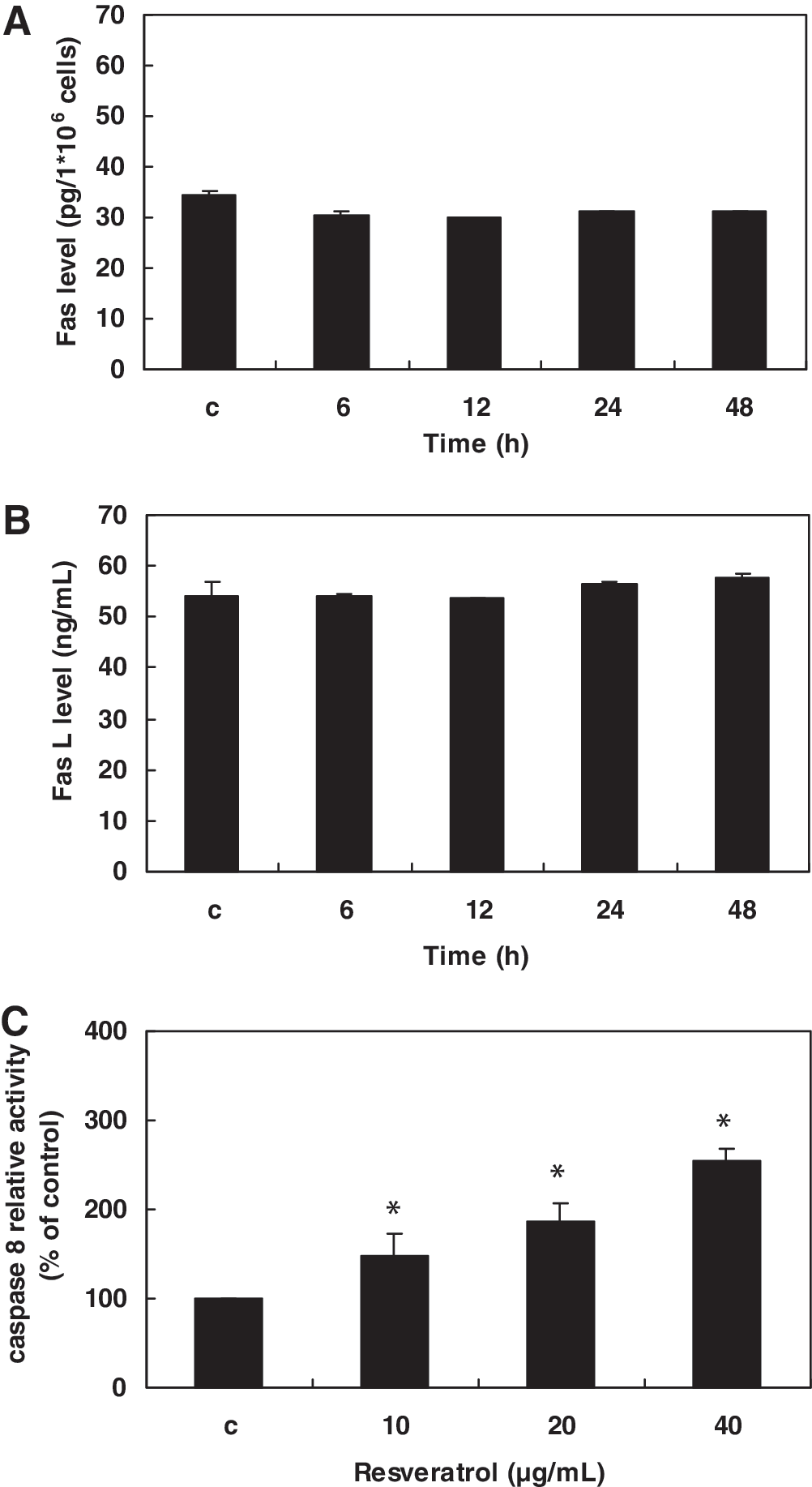
Effect of resveratrol on Fas, Fas ligand (Fas L), and caspase 8 expression in Huh-7 cells:

Effects of caspase 8 and caspase 9 inhibitors on resveratrol (Resv)-treated Huh-7 cells. Cells were treated with 40 μg/mL Resv for 48 hours. Data are mean ± SD values of triplicate experiments. *P < .05, significant difference between control and Resv-treated cells as analyzed by Scheffé's test.
Resveratrol activates the mitochondrial apoptotic pathway
To investigate whether the mitochondrial apoptotic pathway was involved in resveratrol-induced cell death, we determined the levels of pro-apoptotic protein (Bax) and anti-apoptotic proteins (Bcl-2 and Bcl-xL). Results showed that resveratrol treatment decreased expression of the anti-apoptotic proteins Bcl-2 and Bcl-xL but has no effect on the pro-apoptotic protein Bax (Fig. 7A). The ratio of Bax/Bcl-2 or Bax/Bcl-xL was increased by resveratrol treatment, which indicates an important role in triggering the mitochondrial apoptotic pathway.

Resveratrol-induced apoptosis through the mitochondrial apoptotic pathway.
Mitochondrial dysfunctions have been proposed to be the early events during apoptotic cell death. We therefore assessed the change in mitochondrial membrane potential using the mitochondria-specific dye JC-1. Time-dependent mitochondrial dysfunction was induced by resveratrol as indicated by the ratio of red/green fluorescence, which was significantly decreased compared with the control group (Fig. 7B).
The mitochondrial apoptotic process eventually leads to the activation of the caspases, which represent both initiators and effectors of the apoptotic programmed cell death. Similar to the above observations on caspase 8, treatment with resveratrol significantly increased the activity of caspase 9 in Huh-7 cells (Fig. 7C). Furthermore, pretreatment with the caspase 9 inhibitor LEHD-fmk protected Huh-7 cells against resveratrol-induced cell death (Fig. 6).
Two major distinct apoptotic pathways have been described for mammalian cells: the mitochondria-dependent pathway and the mitochondria-independent pathway. Resveratrol treatment resulted in a significant decrease in the levels of Bcl-2 and Bcl-xL, suggesting that the change in the ratio of pro-apoptotic/anti-apoptotic Bcl-2 family proteins might contribute to the apoptosis-promoting activity of resveratrol. These regulatory effects of resveratrol on the Bcl-2 family are associated with the release of cytochrome c from the mitochondria into the cytoplasm and the activation of caspase 9. 36 The importance of this pathway was further confirmed in our subsequent experiment with caspase 9 inhibition. Caspase 8, which is regulated by the death receptor and growth factor pathway, was observed to be stimulated by resveratrol. Inhibition of caspase 8 protected the cells against resveratrol-induced cell death, suggesting that the death receptor-caspase 8 signaling cascade may be important in the resveratrol-induced apoptotic process.
Resveratrol negatively affects the activity of ERK1/2 and p38
As shown in Figure 8A, treatment with 10 μg/mL resveratrol resulted in a rapid inhibition of p-ERK1/2 in the Huh-7 cells and an inhibitory effect on the expression of p-p38 at 24 and 48 hours. However, resveratrol showed no effect on the expression of total ERK1/2 and total p38. To clarify the possible role of ERK1/2 and p38 in resveratrol-mediated cell death, the Huh-7 cells were pretreated with U0126, the chemical inhibitor for mitogen-activated protein kinase 1/2 (an upstream kinase in the ERK1/2 signaling pathway), and a potent specific inhibitor for p38, SB203580. Results showed that pretreatment with either U0126 or SB203580 increased the resveratrol-mediated cell death (Fig. 8B). Furthermore, we also observed that pretreatment with SB203580 strongly increased the activity of caspase 3. Taken together, these observations indicate that ERK1/2 and p38 have important roles in regulating the Huh-7 cell death and are involved in resveratrol-induced apoptosis.

The role of extracellular signal-regulated kinase (ERK) 1/2 and p38 in resveratrol-induced Huh-7 cell apoptosis.
The mitogen-activated protein kinase family of proteins comprises a large number of serine/threonine kinases involved in regulating a wide array of cellular processes, including proliferation, differentiation, stress adaptation, and apoptosis. In general, the p38 kinase pathway is often associated with an increased apoptosis, whereas the ERK1/2 pathway has been shown to suppress apoptosis. 37,38 Interestingly, our results showed that the inhibition of both ERK1/2 and p38 is involved in resveratrol-mediated apoptosis in Huh-7 cells.
The p38 mitogen-activated protein kinase pathway is known to be active in many cancer cells and may result in enhanced resistance to apoptosis. In some types of cancer cells, the inhibition of p38 can reduce cell survival and enhance the effects of chemotherapeutic drugs. Previous studies have documented such effects in cell death induced by delphinidin, 39 docosahexaenoic acid, 40 pomegranate fruit extract, 41 and genistein. 42 Similar to these findings, we observed that inhibition of p38 resulted in an increase in resveratrol-mediated cell death in Huh-7 cells. Furthermore, whereas ERK1/2 inhibition did not significantly increase the activity of caspase 3 following resveratrol treatment, inhibition of p38 by SB203580 led to a strong caspase 3 activation under the same treatment (Fig. 8C). This finding suggests that p38 plays an important role in mediating protection against resveratrol-triggered apoptosis.
Resveratrol causes autophagic cell death
To examine whether autophagy was involved in the resveratrol-mediated Huh-7 cell death, we measured the major proteins of autophagic cell death process, namely, Atg5, Atg7, Atg9, and Atg12. Results demonstrated that resveratrol at 10 μg/mL caused a minimal effect on the expression of autophagic proteins (data not shown), but a time-dependent trend of induction on Atg5, Atg7, Atg9 and Atg12 expression was noted with 20 μg/mL (Fig. 9), suggesting that resveratrol may trigger autophagy in Huh-7 cells.
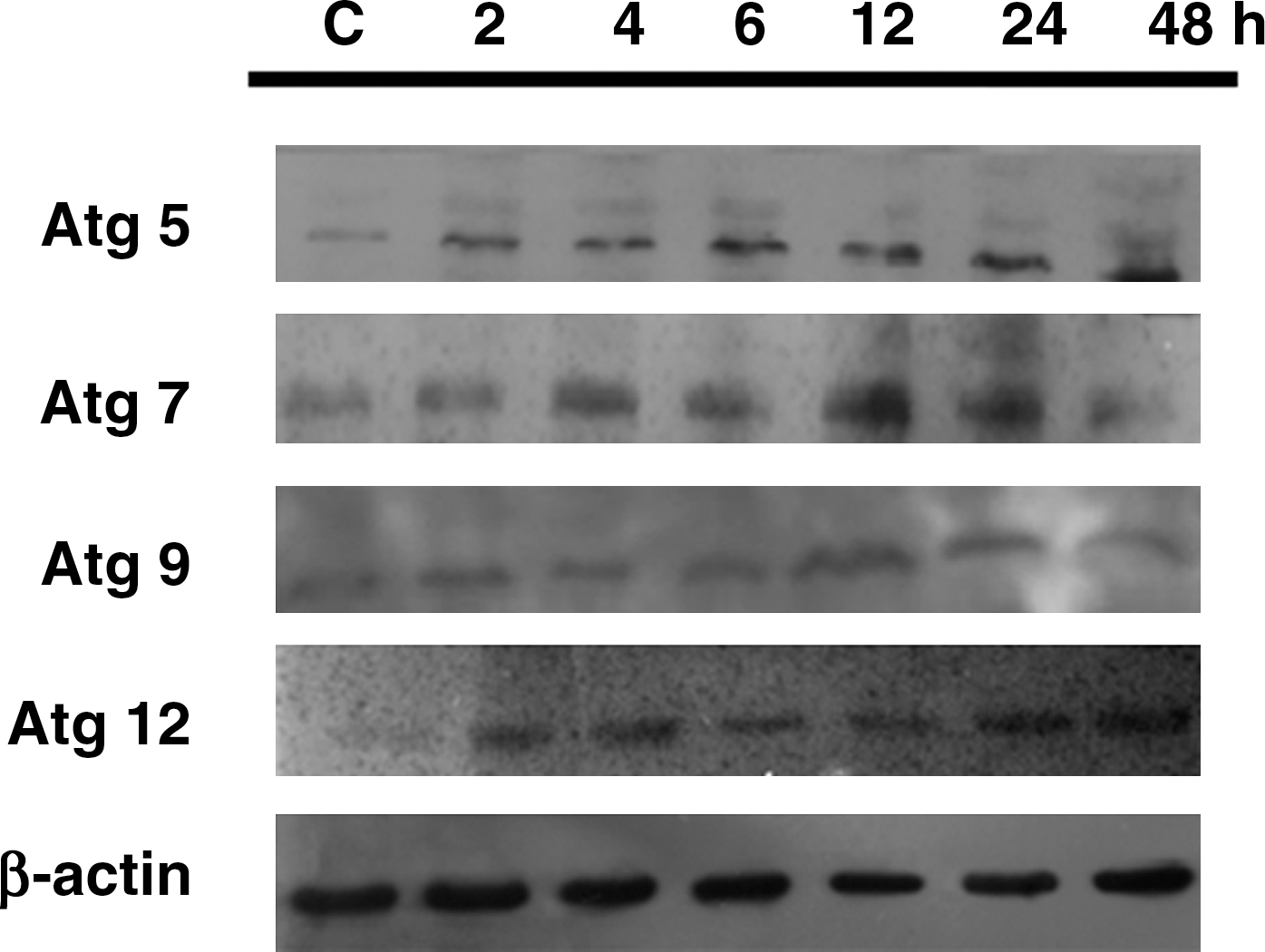
The role of autophagy in resveratrol-mediated Huh-7 cell death. The expressions of autophagy-related Atg5, Atg7, Atg9 and Atg12 proteins were determined by western blotting after treating the Huh-7 cells with 20 μg/mL resveratrol for 0, 2, 4, 6, 12, 24, and 48 hours. C, control (0.1% dimethyl sulfoxide).
Autophagy is an evolutionarily and highly conserved catabolic pathway that degrades cellular macromolecules and organelles. Resveratrol has previously been reported to induce autophagy in cervical, 43 ovarian, 29 gastric, 44 and breast 45 cancer cells. In this study, we also noted that resveratrol can induce autophagic cell death in Huh-7 cells. Autophagy and apoptosis can be triggered by the same upstream signals and thus may result in combined autophagy and apoptosis. This indicates that the apoptotic and autophagic machineries share common pathways that either link or polarize each cellular death responses. It has been reported that the depolarized mitochondria would move into autophagosomes, and thus the mitochondrial dysfunction may be a point of overlap between apoptotic and autophagocytic processes. 46
Several signal transduction pathways that are triggered by common cellular stress can induce both autophagy and apoptosis, such as the general stress mediators, reactive oxygen species, p53, and BH3-only proteins. The p53 protein is an essential tumor suppressor and apoptosis inducer. However, many reports indicate that p53 plays a positive role in cell survival in response to physiological stress, such as by stimulating antioxidant pathways 47 and autophagy. 48 Beclin-1 is a novel BH3-only protein. Like other BH3-only proteins, beclin-1 interacts with members of the anti-apoptotic Bcl-2 family via the BH3 domain. 49,50 The other BH3-only proteins, such as BAD and ABT737 (a BH3-mimetic compound), disrupt the interaction between beclin-1 and Bcl-2 or Bcl-xL, 49,51 thus liberating beclin-1 from an inhibitory complex and allowing it to activate VPS34, a lipid kinase that stimulates autophagy. It is unknown if the regulation of autophagic-initiating proteins is also involved in the mechanism(s) of action of resveratrol in Huh-7 cells. Nevertheless, to our knowledge, the present study was the first to demonstrate that resveratrol possesses potent antiproliferative activity against Huh-7 cells, a new human hepatoma cell line system for studying hepatitis C virus. 34,35 Its mode of actions was mediated through S-phase cell cycle arrest and apoptotic induction, as well as triggering autophagy in the Huh-7 cell system.
Conclusions
The present study concludes the following: (1) resveratrol possesses potent antiproliferative effects on human hepatoma Huh-7 cells; (2) the reduced survival of Huh-7 cells after exposure to resveratrol is associated with S-phase cell cycle arrest and apoptotic induction; (3) resveratrol inhibited cell cycle progression at the S phase by increasing p21/WAF1 and decreasing the expression of CDK2, cyclin A, and cyclin E; (4) resveratrol-induced cell death in Huh-7 cells is mediated by the inhibition of p38 and ERK1/2; and (5) resveratrol induced autophagic cell death in Huh-7 cells. These data have provided a basic mechanism for the chemopreventive properties of resveratrol in Huh-7 cells and provide evidence for potential therapeutic efficacy.
Footnotes
Acknowledgment
C.D.R. provided the Huh-7 cells.
Author Disclosure Statement
No competing financial interests exist.