
Abstract
Boussingaultia gracilis Miers var. pseudobaselloides Bailey is used as a Chinese folk medicine for treatment of diabetes, inflammation, and liver disease. The present study is to investigate the anti-obesity and hypolipidemic effects of B. gracilis Miers var. pseudobaselloides Bailey ethanol extract (BGE). Six-week-old Sprague–Dawley male rats were separately fed for 6 weeks with two kinds of diets—a normal diet (ND) and a high-calorie high-fat diet (HD). Then the animals were treated with tea catechin (100 mg/kg) or BGE (300, 600, or 900 mg/kg) for another 6 weeks. BGE significantly lowered body weight gain, fat-pad weights, and serum and hepatic lipid levels in HD-induced obese rats. The lipid droplets in hepatic tissue of BGE-treated groups were also markedly lessened compared with HD group rats via oil red O staining. Significant increases were observed in the expressions of genes for peroxisome proliferator-activated receptor (PPAR) α and for fatty acid oxidation and thermogenesis-related proteins—acyl-coenzyme A oxidase, carnitine palmitoyl transferase-1, and uncoupling protein-2—in the liver of the BGE-treated groups. Moreover, BGE was found to suppress the expression of sterol response element binding protein-1, a lipogenic gene, as well as those of fatty acid synthase and PPARγ in adipose tissue and liver of HD group rats. These results indicate that B. gracilis Miers var. pseudobaselloides Bailey may have an anti-obesity and hypolipidemic effect through regulation of expression of genes involved in lipolysis and lipogenesis.
Introduction
To treat and control obesity, other than diet therapy and exercise, many different approaches have been suggested, such as drugs for weight loss, appetite suppressants, and food supplements. However, there have been no drugs with long-term efficacy, but without adverse side effects, for the treatment of obesity. Medicinal plants have been used as dietary supplements for body weight management and control in many countries. 5 Some herbal products and plant extracts, such as Semen Cassiaem and Panax ginseng berry extract, have been shown to exert anti-obesity effects in rodents with high fat diet-induced obesity. 6 –9 Boussingaultia gracilis Miers var. pseudobaselloides Bailey, originally distributed in Brazil, has been cultivated in China since 1976, where it is known as Tenq-san-qi, and used as a substitute for Yun-nan-bai-yao (a styptic drug). 10 It is widely used as a hypoglycemic agent, as an anti-inflammatory analgesic, and as a stomachic in folk medicine. 11 Now it is consumed as a new leafy vegetable in China and has proven moderate antimutagenic activity. 12
In the present study, we investigated the anti-obesity and hypolipidemic effects of B. gracilis Miers var. pseudobaselloides Bailey ethanol extract (BGE) in high-calorie high-fat diet (HD)-induced obese rats. Body weight gain, fat-pad weights, and serum and hepatic lipid levels were examined. Green tea catechin, which showed anti-obesity, antidiabetic, and cardioprotective effects in animal and human studies, 13 was used as a positive control. To gain insight into the molecular mechanisms underlying the effects described above, we investigated genes likely to be involved in BGE-regulated anti-obesity action and lipid metabolism, such as the expression of genes for the lipogenesis-related proteins peroxisome proliferator-activated receptor (PPAR), fatty acid synthase (FAS), and sterol regulatory element binding protein-1 (SREBP-1), and the expression of genes for fatty acid oxidation-related proteins acyl-coenzyme A oxidase (ACO), carnitine palmitoyl transferase-1 (CPT-1), and the thermogenic protein, uncoupling protein (UCP)-2.
Materials And Methods
BGE
The dried leaves of B. gracilis Miers var. pseudobaselloides Bailey were obtained from Shenyang Pharmaceutical University, Shenyang, China. Chopped dried leaves were extracted three times with a 10-fold volume of 70% fermentation ethanol at 85°C for 4 hours. The resulting extract was filtered and then lyophilized to yield 22.5% (of the initial amount) of powder, and the extracts were stored in a refrigerator (–30°C) for functional studies. The extract powder contained 29.3% crude protein, 2.5% crude fat, and 13.0% fiber, as well as 53.5 mg/g total flavanoid. The compounds were determined by the pharmaceutical analysis laboratory of the School of Pharmacy, Shenyang Pharmaceutical University.
Animals and experimental diets
Thirty-six 6-week-old male Sprague–Dawley rats (Koatech, Pyeongtaek, Gyeonggi, Republic of Korea) were housed in a temperature (21 ± 2.0°C)- and humidity (50 ± 5%)-controlled room with a ratio of 12-hour light/12-hour darkness and were fed a commercial diet (Koatech) for 1 week. After that, the animals were separately fed with two kinds of diets, a normal diet (ND) and HD (Research Diets Inc., New Brunswick, NJ, USA), for 6 weeks, and then the animals were subdivided into six groups (n = 6)—ND group, HD group, HD + BGE (300, 600, or 900 mg/kg) groups, and HD + Tea (tea catechin, 100 mg/kg) group—and treated for another 6 weeks. Tea catechin was kindly provided by Amore Pacific Corp. (Seoul, Republic of Korea).
The compositions of ND and HD are shown in Table 1. The diets were given in the form of pellets. The food intakes were recorded daily, and animals' body weights were monitored every 3 days. At the end of the treatment period, the rats were anesthetized with diethyl ether after an overnight fast. Blood samples were drawn from the abdominal aorta and centrifuged at 890 g for 15 minutes to obtain the serum; the serum was kept at −70°C until analysis. The epididymal, abdominal, visceral, and brown fat-pads and livers were surgically removed, weighed, snap-frozen in liquid nitrogen, and stored at −70°C until use. This study was approved by the Institutional Animal Care and Use Committee of Kyung Hee University, Seoul.
Data are kcal.
HD, high-calorie high-fat diet; ND, normal diet.
Biochemical analysis
Serum concentrations of total cholesterol (TC), high-density lipoprotein-cholesterol (HDL-C), free fatty acid (FFA), and triglyceride (TG) were determined enzymatically using commercial kits (Asan Diagnostics, Seoul). Serum insulin, adiponectin, and tumor necrosis factor (TNF)-α levels were measured by enzyme-linked immunosorbent assay using commercially available kits (Linco Research, St. Charles, MO, USA). To analyze hepatic lipid content, the lipids were extracted by the method of Folch et al. 14 Hepatic TG, HDL-C, and TC were determined using commercial kits (Asan Diagnostics, Seoul), and hepatic FFA was mesured by the ACS-ACOD method using a NEFA-HR kit (Wako Pure Chemical Industries, Osaka, Japan).
Oil red O staining
Staining with oil red O was performed in accordance to Woo et al. 15 and captured at ×200 magnification. The liver was fixed in 10% formalin solution and embedded in paraffin wax. Frozen liver sections from fixed tissues were cut into 10-μm pieces and mounted on a slide. The samples were stained with oil red O in propylene glycol and with hematoxylin/eosin as the counterstain.
RNA isolation and reverse transcription–polymerase chain reaction analysis
The total RNA was isolated from the liver and epididymal fat tissue of each rat using an Easy-spin™ total RNA extraction kit (iNtRON, Seongnam, Republic of Korea). RNA concentrations were quantified by obtaining the absorbance at 260 nm and 280 nm of an appropriate dilution on an Optizen 2120UV spectrophotometer (Mecasys, Daejeon, Republic of Korea). Reverse transcription (RT)–polymerase chain reaction (PCR) assays for target mRNA levels in adipose tissue and liver were performed using a Maxime™ RT-PCR PreMix kit (iNtRON) according to the manufacturer's protocol. The primers for the PCR analysis were synthesized at Bioneer (Daejeon). Using the primers listed in Table 2, the RT-PCR was performed at 45°C for 30 minutes. Then the PCR was carried out as follows: 5 minutes at 94°C, 30 cycles of (94°C for 1 minute, 56°C for 1 minute, and 72°C for 5 minutes), and a 5-minute incubation at 72°C. The β-actin gene was used as an internal control for semiquantitative RT-PCR.
F, forward; R, reverse.
Statistical analysis
Statistical analysis was performed by Dunnett's multiple comparison method using one-way analysis of variance (SPSS version 13.0; SPSS, Inc., Chicago, IL, USA). Differences between treatment and control groups were considered statistically significant at P < .05. Data are presented as mean ± SD values.
Results And Discussion
Body weight, food efficiency ratio, and fat-pad weights
To examine the anti-obesity and hypolipidemic effects of BGE in HD-induced obese rats, we fed rats two different diets for 6 weeks. The rats fed HD-only had significantly higher body weights and fat-pad weights than the rats fed ND. Then they were treated with BGE or tea catechin for another 6 weeks. Effects of BGE on body weight and food efficiency ratio in HD-induced obese rats are shown in Table 3. At the end of the experiment, administration of BGE (900 mg/kg) or tea catechin for 6 weeks remarkably decreased the body weight gain by more than 10% compared with the HD group. BGE significantly and dose-dependently decreased the final body weight and food efficiency ratio. In addition, the epididymal, abdominal, visceral, and brown fat-pad weights of the obese rats given BGE (600 or 900 mg/kg) were reduced significantly compared with the HD-only rats (Table 4).
Data are mean ± SD values (n = 6).
P < .05, ## P < .01, ### P < .001 versus ND group; *P < .05, **P < .01, ***P < .001 versus HD groups.
FER, food efficiency ratio.
Data are mean ± SD values (n = 6).
P < .05, ### P < .001 versus ND group; *P < .05, **P < .01 versus HD groups.
Biochemical analysis in serum and liver
In general, a high-fat diet significantly increases the TC and TG levels in serum and liver compared with normal diets. In our experiment, the serum and hepatic lipid levels were measured and are shown in Figures 1 and 2, respectively. Rats in the HD group exhibited significantly higher TG, TC, and FFA levels as well as lower HDL-C and HDL-C/TC ratio. However, both BGE and tea catechin led to a reversal of the aforementioned parameters to levels similar to those of the ND group. BGE treatment with 900 mg/kg resulted in 31%, 22%, and 61% decreases in serum TG, TC and FFA, respectively, and a 15% and 49% increase in HDL-C and HDL-C/TC ratio, respectively, compared with the HD rats. With the same BGE (900 mg/kg) administration, the hepatic TG, TC, and FFA levels exhibited 43%, 37%, and 52% decreases, respectively. These results indicate that oral administration of BGE dose-dependently suppresses the accumulation of body fat, resulting in improved lipid profiles in serum and liver.
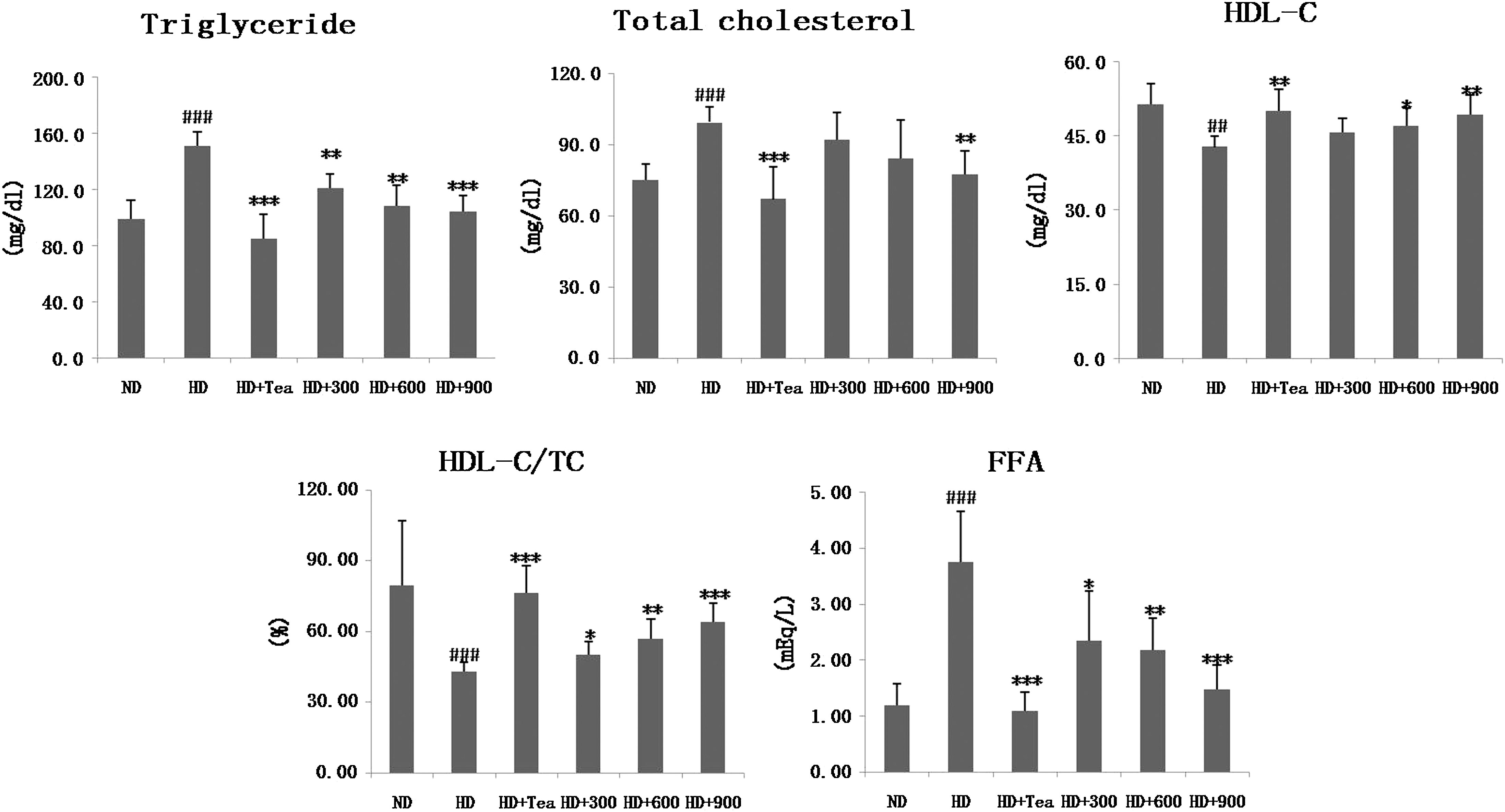
Effects of B. gracilis Miers var. pseudobaselloides Bailey extract on serum lipid levels in HD-induced obese rats. Data are mean ± SD values (n = 6). ## P < .01, ### P < .001 versus the ND group; *P < .05, **P < .01, ***P < .001 versus HD groups. FFA, free fatty acid; HDL-C, high-density lipoprotein-cholesterol; TC, total cholesterol.
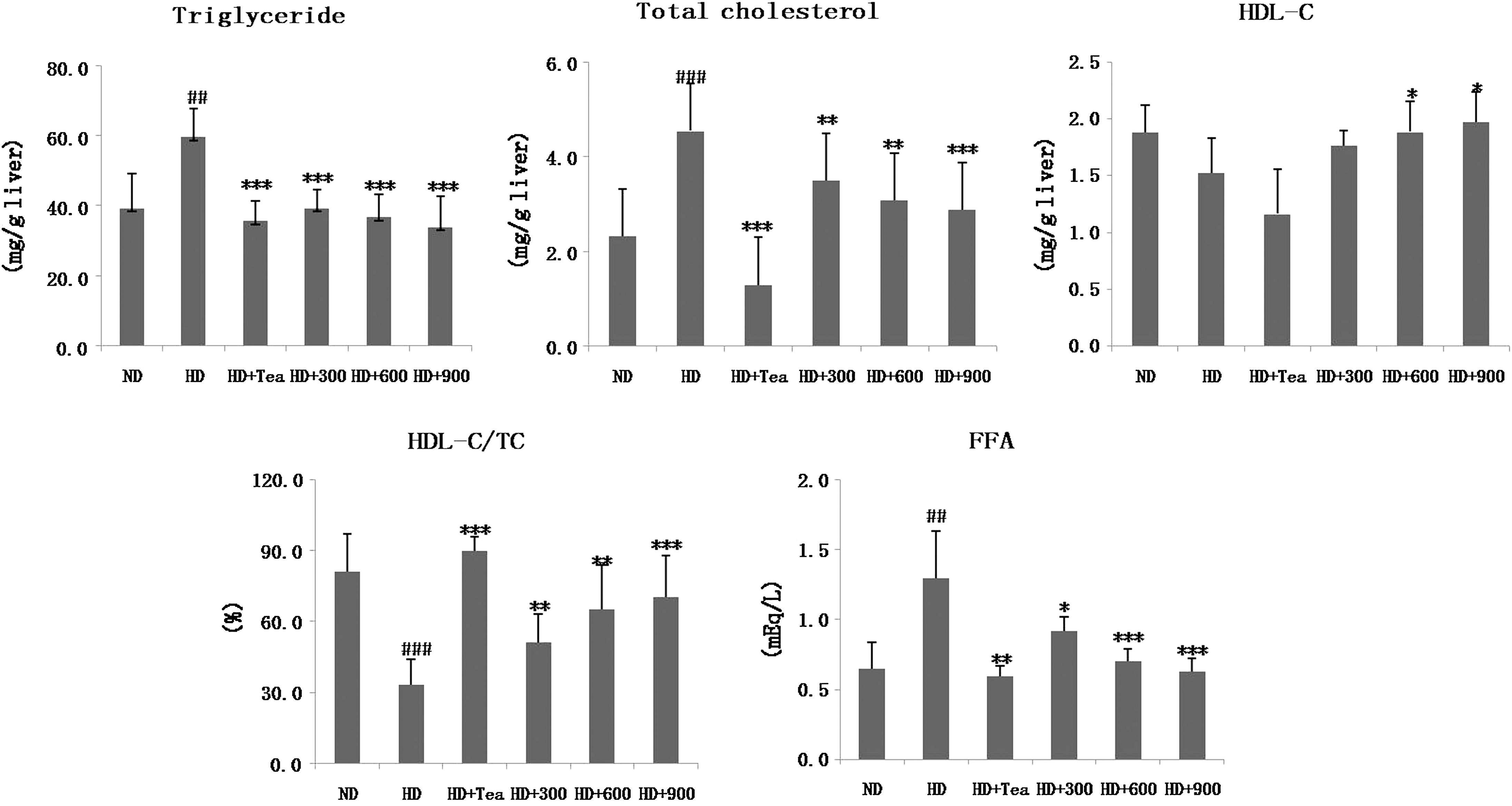
Effects of B. gracilis Miers var. pseudobaselloides Bailey extract on hepatic lipid levels in HD-induced obese rats. Data are mean ± SD values (n = 6). ## P < .01, ### P < .001 versus ND group; *P < .05, **P < .01, ***P < .001 versus HD groups.
Excessive visceral fat accumulation during high-fat feeding causes a disturbance in cytokine secretion from adipose tissue and is involved in the pathogenesis of type 2 diabetes, cardiovascular disease, and hypertension. 16 Therefore, the levels of serum glucose, insulin, TNF-α, and adiponectin were also examined and are presented in Table 5. The glucose, insulin, and TNF-α levels were greater in the HD group than in the ND group. Conversely, the adiponectin level was significantly lower in the HD group. Again, BGE (900 mg/kg) treatment reduced the serum glucose, insulin, and TNF-α levels and elevated the adiponectin level to values similar to those of the ND group. Tea catechin also showed parallel effects to BGE at 900 mg/kg (Table 5). The lower TNF-α and adiponectin levels could be attributed to a decrease in the fat mass of rats that were treated with BGE. Many studies have reported that TNF-α secretions are elevated through the accumulation of fat in adipocytes 17 and that TNF-α induces insulin resistance in obese animal models. 18 Moreover, serum adiponectin levels have been reported to be reduced in obese rodent models and humans and to correlate inversely with insulin resistance. 19 –23 A previous article reported that the butanol-soluble fraction from B. gracilis Miers var. pseudobaselloides Bailey showed hypoglycemic activity, which afforded a complex mixture of tritepenoid glycosides. 24 We presume that the ethanol and butanol extracts may possess the same hypoglycemic active component. Our results indicate insulin levels were consistent with glucose levels and were related to body fat mass, putatively because of changes in insulin sensitivity due to obesity.
Data are mean ± SD values (n = 6).
P < .01, ### P < .001 versus ND group; *P < .05, **P < .01, ***P < .001 versus HD groups.
TNF-α, tumor necrosis factor-α.
Liver histology
Because liver is a major target tissue for lipid and lipoprotein metabolism, we processed the oil red O-stained sections of livers from normal and obese rats treated with BGE or tea catechin. As shown in Figure 3, livers from ND group rats showed minimal or barely visible fat in hepatic cells as demonstrated by the sparse distribution of small microvesicles within hepatocytes. By contrast, livers from obese rats showed widespread deposition of fat globules of different sizes inside parenchymal cells. Livers from rats treated with BGE (900 mg/kg) showed barely visible fat in hepatic cells as demonstrated by the sparse distribution of small microvesicles within hepatocytes. This is consistent with the aforementioned results showing that BGE improved hepatic lipid profiles.
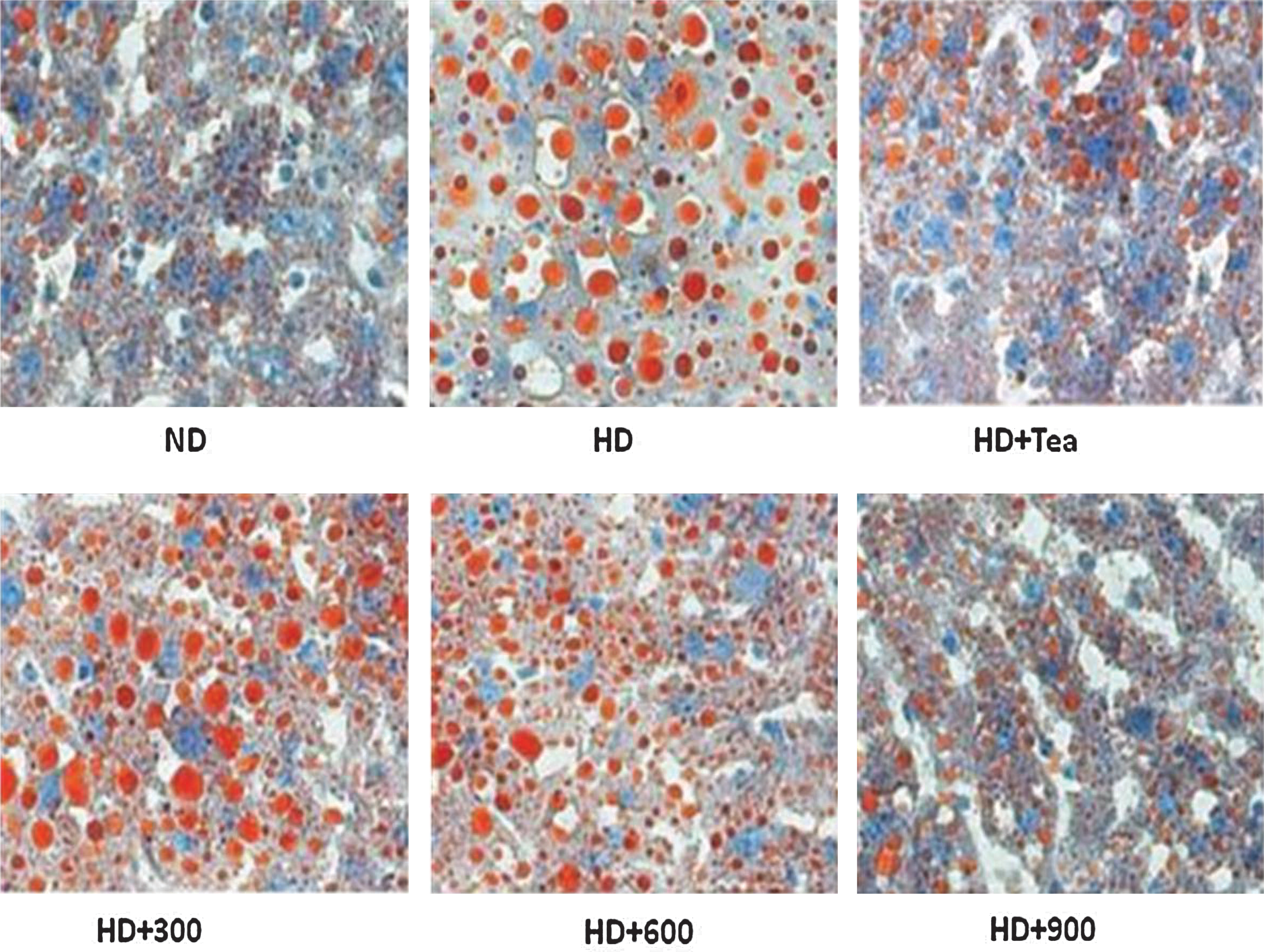
Effects of B. gracilis Miers var. pseudobaselloides Bailey extract on lipid droplet deposition of hepatic tissue by oil red O staining in HD-induced obese rats. Magnification ×200. Color images available online at
Lipolytic and lipogenic gene expressions
To gain insight into the molecular mechanisms underlying the effects described above, we investigated genes likely to be involved in BGE-regulated anti-obesity action and lipid metabolism. Fatty acid oxidation is regulated in part by several molecular markers, including mitochondrial UCP, ACO, and CPT-1, that have been reported to be associated with the AMP-activated protein kinase (AMPK) signaling pathway. 25 AMPK belongs to the metabolic sensor protein kinase family and is activated by intracellular stress conditions such as hypoxia, ischemia, and reactive oxygen species; it inhibits cellular ATP-consuming pathways by inhibiting cell proliferation. 26 AMPK has also been proposed to play a beneficial role in the prevention of metabolic diseases such as type 2 diabetes, obesity, and cancer. 27 Our results support previous studies that reported decreased AMPK mRNA levels in the muscles and adipose tissues of HD-induced obese mice and rats. 28 –30 The results from the semiquantitative RT-PCR analyses of the total RNA prepared from the epididymal adipose tissue and the liver indicate that feeding rats HD apparently down-regulated the expression of the AMPK and UCP2 genes (Figs. 4 and 5). These HD-induced down-regulations in the transcription gene levels were dramatically reversed when the rats were treated with BGE (900 mg/kg) or tea catechin. The adiposity decrease caused by BGE in this study appears to have been due, at least in part, to the enhanced β-oxidation of fatty acid through its function of increasing AMPK protein expression. In this study we also examined changes in the expression of the UCP2 gene, a fatty acid oxidation marker, in HD-induced obese rats. 31 The RT-PCR results clearly indicate that HD feeding decreases the expression of the UCP2 gene in white adipose tissue and the liver of obese rats. UCP2 was suggested to lie downstream of AMPK and PPARα 32,33 and is widely used as a target for the treatment of hyperlipidemia. 34 –36 Based on the observation that BGE-treated rats evidently reversed the HD-induced down-regulation of UCP2 gene expression in the white adipose tissue and the liver, BGE appears to reduce adiposity, at least in part, by raising uncoupled respiration in adipose tissue and liver.
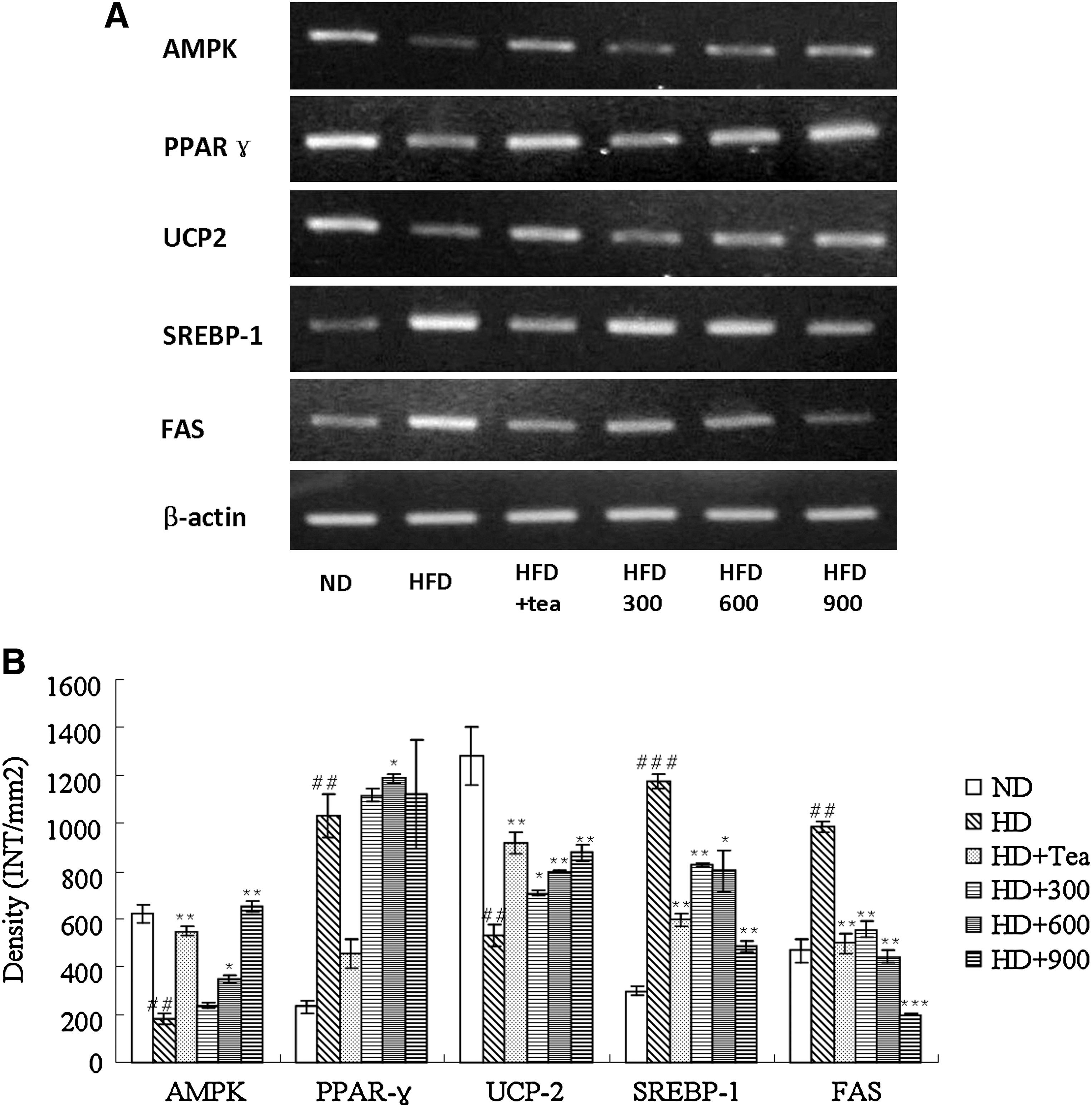
(
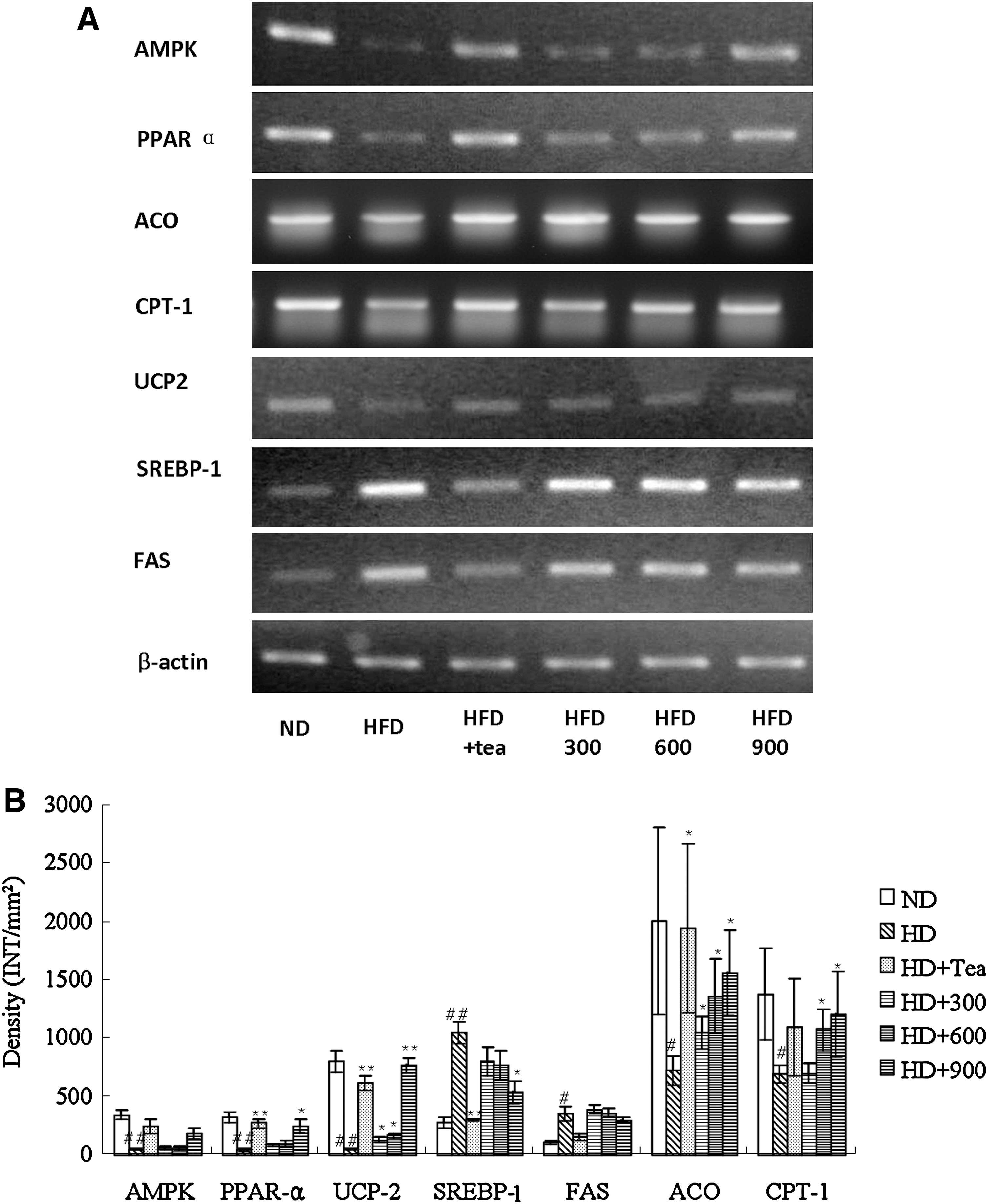
(
The SREBP-1 and FAS genes, which were up-regulated in the adipose tissue and the liver of rats fed with HD, were down-regulated when they were fed with BGE (900 mg/kg) or tea catechin (Figs. 4 and 5). FAS, which plays a central role in de novo lipogenesis in mammals, is sensitive to nutritional status in lipogenic tissues such as liver and adipose tissues. The regulation of FAS expression occurs primarily through changes in FAS gene transcription by SREBP-1, a master regulator of lipogenesis. 37 Recent studies found many of the aforementioned beneficial effects of green tea were attributed to its most abundant catechin by FAS gene suppression. 38,39 Our results indicate that BGE decreases weight gain through down-regulation of SREBP-1, which leads to the inhibition of FAS induction and subsequent inhibition of the lipogenic pathway.
PPARs, a subfamily of the 48-member steroid and nuclear receptor superfamily, are ligand-dependent transcription factors that control energy homeostasis by regulating carbohydrate and lipid metabolism. 40 They exist in three different subtypes—PPARα, PPARβ/δ, and PPARγ—of which PPARα is expressed in liver, kidney, heart, and muscles and is involved in lipoprotein metabolism, and PPARγ is predominantly expressed in adipose tissue and is involved in controlling insulin resistance, adipocyte differentiation, and lipid storage. PPARα was down-regulated in the liver of obese rats, whereas PPARγ was up-regulated in the adipose tissue in the current study (Figs. 4 and 5). Some studies have suggested that fatty acid oxidative PPARα target genes and PPARα expression are suppressed in obese mice, resulting in hypertriglyceridemia and hypercholesterolemia, 41,42 and hepatic PPARα activation may be crucial in energy homeostasis, such as in body weight control and lipid metabolism. 43 –46 On the other hand, PPARγ is activated under conditions of adipocyte differentiation, and the inhibition of PPARγ expression with specific ligands can successfully induce anti-obesity effects. 47,48 PPARα and PPARγ gene expressions in both the BGE-treated groups were significantly up-regulated. Gene expressions of molecules regulated by PPARα (ACO, a rate-controlling enzyme involved in fatty acid β-oxidation, and CPT-1, regulating acyl-coenzyme A inflow and involved in thermogenesis) were also elevated. Increased hepatic PPARα and decreased adipose tissue PPARγ transactivation with decreased fat mass and serum TG and TC in BGE-treated obese rats indicate that BGE regulated obesity and lipid metabolism in part because of activation of PPARs in liver and adipose tissue.
Tea catechin was reported to act as a pro-oxidant in hepatoma cells and increase UCP2 expression in liver, which could be a means of limiting reactive oxygen species production. 49 The increases of UCP2, ACO, and CPT-1 expression in liver are compatible with increased energy metabolism in liver induced by catechin and could possibly be the consequence of an increased fatty acid oxidation. BGE showed anti-obesity and hypolipidemic activity similar to that of tea catechin in our findings. However, further study is needed to investigate which compounds in the BGE are responsible for the effect, as well as the mechanisms responsible for the activities.
In conclusion, BGE reversed the effects of HD on body weight and serum lipids, and these seem to be mediated by regulation of lipolytic and lipogenic gene expression.
Footnotes
Acknowledgment
The samples of B. gracilis Miers var. pseudobaselloides Bailey were generously donated by Professor Bi Kai Shun of Shenyang Pharmaceutical University, Shenyang, China.
Author Disclosure Statement
No competing financial interests exist.