
Abstract
Cynanchum wilfordii is used in traditional Chinese medicine with almost all parts of this plant considered beneficial for various vascular diseases. This study was performed to evaluate the effect of an ethanol extract of C. wilfordii (ECW) on vascular dysfunction in apolipoprotein E (apoE)−/− mice fed with high fat/cholesterol diets (HFCDs). The apoE −/− mice were fed HFCD consisting of 7.5% cocoa butter and 1.25% cholesterol, with or without 100 or 200 mg/day/kg ECW. Chronic ECW treatment significantly lessened the level of low-density lipoprotein (P<.05) and elevated that of high-density lipoprotein-cholesterol (P<.01). Chronic ECW treatment normalized the HFCD-induced increase in systolic blood pressure, maintained smooth and soft intimal endothelial layers, and decreased intima-media thickness in aortic sections of HFCD-fed apoE −/− mice. ECW significantly restored the diet-induced decrease in vasorelaxation response to acetylcholine; however, the response to sodium nitroprusside did not change. ECW clearly restored the HFCD-induced reduction in endothelial nitric oxide synthase expression levels in aortic tissue, leading to decreased vascular inflammation through an inhibition of cellular adhesion molecules such as E-selectin, vascular cell adhesion molecule-1, and intracellular adhesion molecule-1 as well as endothelin-1 (ET-1) expression. In conclusion, ECW ameliorates endothelial dysfunction via improvement of the nitric oxide/cyclic GMP signaling pathway in a diet/genetic model of hyperlipidemia. ECW also substantially inhibited the development of atherosclerosis, possibly by inhibiting ET-1, cell adhesion molecules, and lesion formation, suggesting a vascular protective role for this herb in the treatment and prevention of atherosclerotic vascular disease.
Introduction
L
Hypertension has been identified as an independent risk factor for the development of endothelial dysfunction and inflammation. 7 Endothelin (ET)-1 expression and level are significantly higher in aortic and mesenteric arteries of hypertensive animal models. Hypertensive patients with high plasma ET-1 levels often exhibit elevated cell adhesion molecule levels and increased risks for developing hypertension-induced organ damage. 8 One early phase of atherosclerosis involves the recruitment of inflammatory cells from the circulation and their transendothelial migration. 9 This process is predominantly mediated by cellular adhesion molecules, which are expressed on the vascular endothelium and on circulating leukocytes in response to several inflammatory stimuli. Selectins and their ligands are involved in the rolling and tethering of leukocytes on the vascular wall. Intracellular adhesion molecule-1 (ICAM-1) and vascular adhesion molecule-1 (VCAM-1) induce firm adhesion of inflammatory cells at the vascular surface. 10
Apolipoprotein E (ApoE) protein is synthesized in the liver and macrophages and has several important anti-atherogenic functions. As a constituent of plasma lipoproteins, ApoE serves as a ligand for cell surface lipoprotein receptors such as low-density lipoprotein (LDL) receptor and LDL receptor–related proteins, 11 thereby promoting the uptake of atherogenic particles from the circulation. Consequently, homozygous deletion of the apoE gene in mice results in a pronounced increase in the plasma levels of LDL and very-low-density lipoprotein attributable to the failure of LDL receptor–mediated and LDL receptor–related protein–mediated clearance of these lipoproteins. 12
The white root tuber of Cynanchum wilfordii, which is called Paeksuo or Paekhasuo in Korea and Baishouwu in China, is one of the most famous traditional medicines used as a blood tonic, enriching vitality and enhancing immunity. 13 It has been confirmed to exhibit a beneficial effect on vascular diseases. 14 Recently, it is found that C. wilfordii decreases serum cholesterol in mice 15 and protects cortical neurons from toxicity induced by hydrogen peroxide. 16 Gagaminine isolated from the root of C. wilfordii inhibits aldehyde oxidase activity and lipid peroxidation in vitro. 17 However, little information is available concerning the pharmacological basis of the activity of C wilfordii on vascular function. Here, we investigated the effects of an ethanol extract of C. wilfordii (ECW) on vascular dysfunction in apoE −/− mice fed HFCD.
Materials and Methods
Preparation of ECW
C. wilfordii (Max.) Hemsley tuber was purchased from the Herbal Medicine Cooperative Association, Jeonbuk Province, Korea and authenticated by Prof. Tae-Oh Kwon, College of Life Sciences and Natural Resources, Wonkwang University, Iksan, Jeonbuk, Korea. A voucher specimen (number DH-127) was deposited in the herbarium of the Professional Graduate School of Oriental Medicine, Wonkwang University. Tubers were cleaned and air-dried at room temperature. Dried tuber (1.2 kg) was extracted with 10 L of 95% ethanol at 24°C for 1 week. The extract was filtered through Whatman (Maidstone, United Kingdom) No. 3 filter paper and concentrated using a rotary evaporator. The resulting extract (48.3 g) was lyophilized using a freeze-dryer and retained until required.
Nuclear magnetic resonance spectroscopy
Nuclear magnetic resonance (NMR) spectra were recorded using a JEOL (Tokyo, Japan) JNM ECP-400 spectrometer (400 MHz for 1H), and chemical shifts were referenced relative to relative to the corresponding residual solvent signals.
High-performance liquid chromatography fingerprinting and isolation of compounds
The chromatographic fingerprint of ECW was obtained on a Young Lin high-performance liquid chromatography (HPLC) system (Young Lin Instrument Co., Ltd., Kyounggi-do, Korea) equipped with a Young Lin ultraviolet detector (model UV 730D) and model Zam3000 evaporative light scattering detector (Schambeck SFD GmbH, Bad Honnef, Germany). Chromatographic separation was carried out on an Eclipse XDB-C18 column (Agilent Technologies, Santa Clara, CA; 4.6×150 mm; particle size, 5 μm) at room temperature with an injection volume of 50 μL using a gradient elution of 10% methanol in water (0.1% formic acid) to 100% methanol over a 60-minute period. Peaks were detected simultaneously at both 210 nm and 254 nm of ultraviolet detection as well as evaporative light scattering detection. HPLC analysis of the extract indicated the majority of compounds were eluted in less than 5 minutes under the described elution condition (data not shown). This result suggested carbohydrates are the major component presented in the extract. Therefore, a portion of the extract (201.6 mg) was subjected to octadecyl functionalized silica gel flash column (4×25 cm; particle size, 75 μm) chromatography. The column was eluted with 20% methanol in H2O followed by 500 mL of 100% methanol (300 mL each), affording two fractions (180.7 mg and 29.4 mg, respectively). HPLC analysis of the fraction that eluted with 100% methanol indicated the presence of three major components, and thus further semipreparative reversed-phase HPLC (Agilent [Palo Alto, CA, USA] preparative C18 column [21.2×150 mm; particle size, 5 μm]; elution rate, 5 mL/minute; detection at 254 nm; eluting with a gradient from 25 to 100% methanol in H2O [0.1% formic acid] over a 60-minute period) was conducted to yield compounds
Experimental animals
The animal procedures were in strict accordance with the 1985 (revised 1996) guidelines for the care and use of laboratory animals of the U.S. National Institutes of Health and were approved by the Institutional Animal Care and Utilization Committee for Medical Science of Wonkwang University. Six-week-old male apoE −/− C57BL6J mice (n=40) and normal C57BL6J mice (n=10) obtained from Central Laboratory Animal (Seoul, Korea) and housed in metabolic cages with automatically controlled temperature (22±2°C), relative humidity (50–60%), and light (12-hour light/dark cycle). Throughout the experiments, all animals had unrestricted access to water. After a week of acclimatization, animals were randomly divided into five groups (n=10 per group). In the control group, C57BL6J mice received regular diet (RD) (catalog number D10012M, Research Diets, Inc., New Brunswick, NJ, USA). In the fluvastatin group, apoE −/− mice received HFCD+10 mg/kg/day fluvastatin. In the ECW I group, apoE −/− mice received HFCD+100 mg/kg/day ECW. In the ECW II group, apoE −/− mice received HFCD+200 mg/kg/day ECW. The control and HFCD groups received RD and HFCD, respectively, for 10 weeks. The HFCD containing 7.5% cocoa butter and 1.25% cholesterol was purchased from Research Diets, Inc. (catalog number D12451) (Table 1). In the fluvastatin, ECW I, and ECW II groups, the particular treatments began after 10 weeks and continued for the next 10 weeks.
Typical analysis of cholesterol in lard=0.95 mg/g.
Cholesterol (mg)/kg=196.5.
Measurement of blood pressure
Systolic blood pressure was determined by a tail-cuff plethysmography method and recorded with an automatic sphygmotonograph (Muromachi Kikai, Tokyo). At least eight determinations were made in every session, and the mean of the lowest five values within 5 mm Hg was taken as the systolic blood pressure level.
Analysis of plasma biochemicals
Plasma insulin levels were measured by an enzyme-linked immunosorbent assay method using a commercial mouse insulin enzyme-linked immunosorbent assay kit (Shibayagi Co., Ltd., Shibukawa, Japan). The total cholesterol, high-density lipoprotein (HDL)-cholesterol, LDL-cholesterol, and triglycerides in plasma were enzymatically measured using commercially available kits (ARKRAY, Inc., Kyoto, Japan).
Recording of isometric vascular tone
Vascular tone was determined as previously described by Kang et al. 19 At the end of the experiment, mice were sacrificed by decapitation. The thoracic aorta was rapidly and carefully dissected and placed into ice-cold Krebs solution (118 mM NaCl, 4.7 mM KCl, 1.1 mM MgSO4, 1.2 mM KH2PO4, 1.5 mM CaCl2, 25 mM NaHCO3, and 10 mM glucose, pH 7.4). The aortae were removed free of connective tissue and fat and cut into rings with a width of approximately 3 mm. All dissecting procedures were done with extreme care to protect the endothelium from inadvertent damage. The aortic rings were suspended by means of two L-shaped stainless steel wires inserted into the lumen in a tissue bath containing Krebs solution at 37°C. A gas mixture of 95% O2 and 5% CO2 was continuously bubbled through the bath. The baseline load placed on the aortic rings was 1.0 g. Changes in isometric tension were recorded using a Grass model FT 03 force displacement transducer (Grass Technologies, Quincy, MA, USA) connected to a model 7E polygraph recording system (Grass Technologies). The aortic relaxation by the cumulative addition of ACh was performed in the presence of endothelium.
Protein preparation and western blot analysis
Thoracic aortae were homogenized in a buffer consisting of 250 mM sucrose, 1 mM EDTA, 0.1 mM phenylmethylsulfonyl fluoride, and 20 mM potassium phosphate buffer (pH 7.6). Large tissue debris and nuclear fragments were removed by two successive low-speed centrifugations (1,197 g, 5 minutes; 6,296 g, 10 minutes, 4°C). The recovered protein (40 μg) was separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred electrophoretically to nitrocellulose membranes (Amersham, Piscataway, NJ, USA) using a Mini-Protean II apparatus (Bio-Rad, Hercules, CA, USA). Sodium dodecyl sulfate–polyacrylamide gel electrophoresis protein standard was used to check transfer efficiency and as a molecular weight marker. Membranes were blocked with 5% nonfat milk powder in phosphate-buffered saline containing 0.05% Tween 20 for 1 hour prior to incubation in the presence of primary antibodies to E-selectin, VCAM-1, ICAM-1, or β-actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at a final dilution of 1:1,000 overnight at 4°C. The blot was washed several times with phosphate-buffered saline containing 0.05% Tween 20 and incubated with the appropriate horseradish peroxidase–conjugated secondary antibody for 1 hour. After the membrane was washed several times with phosphate-buffered saline containing 0.05% Tween 20, bound secondary antibody was detected by enhanced chemiluminescence (Amersham, Little Chalfont, United Kingdom). Protein expression levels were determined by analyzing the signals captured on the nitrocellulose membrane using a Chemi-Doc image analyzer (Bio-Rad).
Quantitative histopathology
Aortae isolated from all groups were fixed in 10% (vol/vol) formalin in 50 mM potassium phosphate buffer (pH 7.0) for 48 hours at 4°C. The tissues were subsequently embedded in paraffin, and cross-sections (6 μm thick) of the aortic arch in each group were stained by use of hematoxylin and eosin. 20 For quantitative histopathologic comparisons, the mean of 10 sections was taken. Intima-media ratio and internal elastic lamina were determined by length using Axiovision 4 imaging/archiving software (Carl Zeiss, Jena, Germany).
Immunohistochemistry
Sections were stained before they were incubated with 5% normal goat serum for 10 minutes at room temperature to reduce nonspecific background staining. Endothelial NO synthase (eNOS) and ET-1 antibody (Oncogene, Cambridge, MA, USA) were applied as a 1:500 dilution and incubated in humidified chambers overnight at 4°C. All slides were then sequentially incubated with biotinylated secondary antibody and horseradish peroxidase–conjugated streptavidin, both for 10 minutes at room temperature. Peroxidase activity was visualized by the 3-amino-9-ethylcarbazole substrate-chromogen system (Zymed, San Francisco, CA, USA), which resulted in brownish-red staining. Representative sections were photographed, and quantification of immunohistological staining was conducted by Axiovision 4 imaging/archiving software.
Statistical analyses
Data are shown as mean±SE values. Statistical analyses were performed using analysis of variance followed by Student's t test for unpaired data and one-way analysis of variance followed by Bonferroni's multiple-comparison test. P<.05 was considered statistically significant.
Results
1H NMR analysis and HPLC profile
1H NMR analysis of ECW revealed signals corresponding to typical carbohydrate moieties such as anomeric protons at 5.38 ppm and 4.94 ppm, along with a number of oxygenated methines and methylene groups ranging from 3.44 ppm to 4.18 ppm (Fig. 1). In addition, HPLC analysis indicated the presence of three major components: 4-hydroxyacetophenone, 2,5-dihydroxyacetophenone, and 2,4-dihydroxyacetophenone, respectively (Fig. 2).
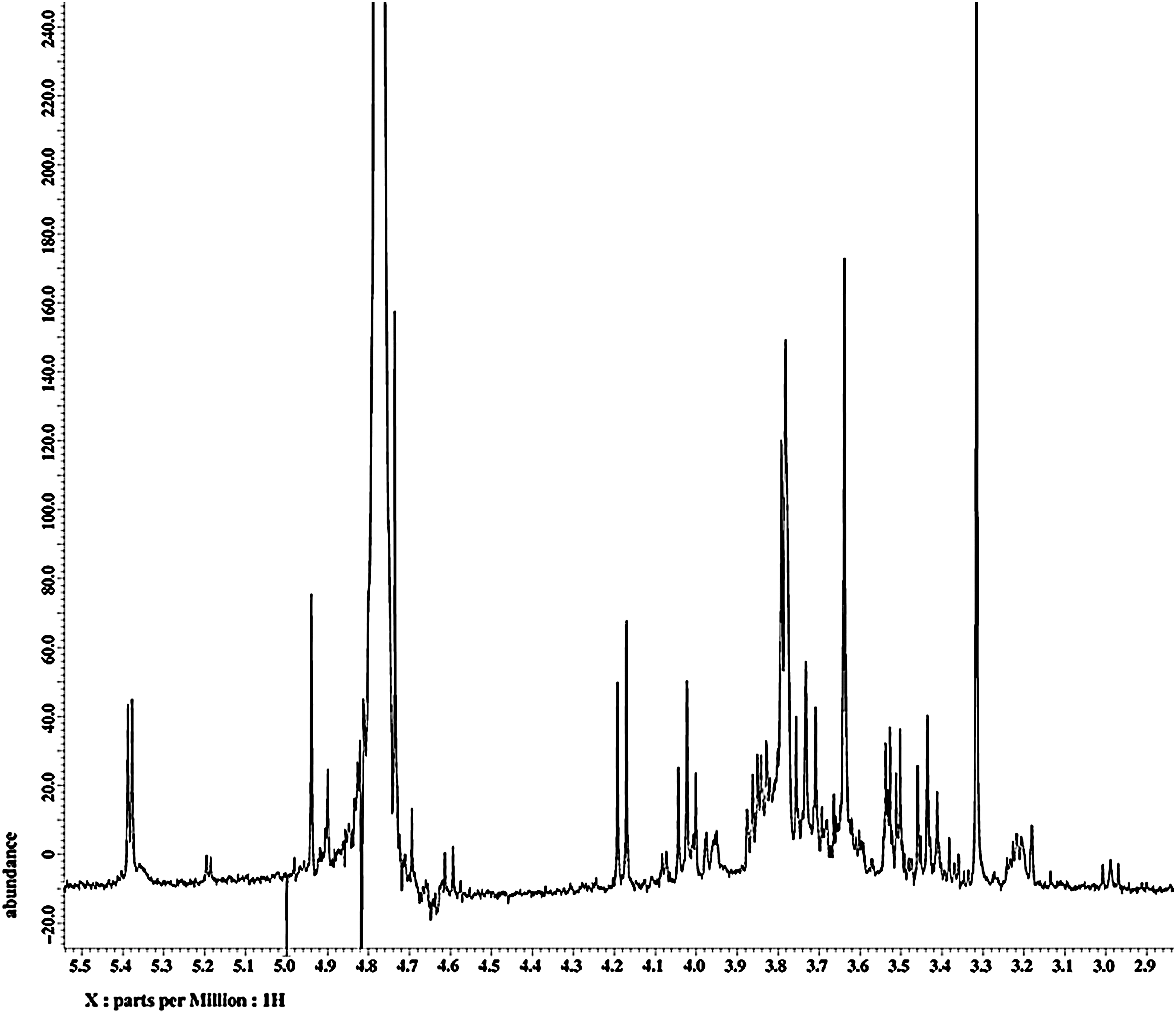
1H nuclear magnetic resonance spectrum of the ethanol extract of C. wilfordii, specifically that of the fraction eluted with 20% methanol in H2O from octadecyl functionalized silica gel flash column chromatography.
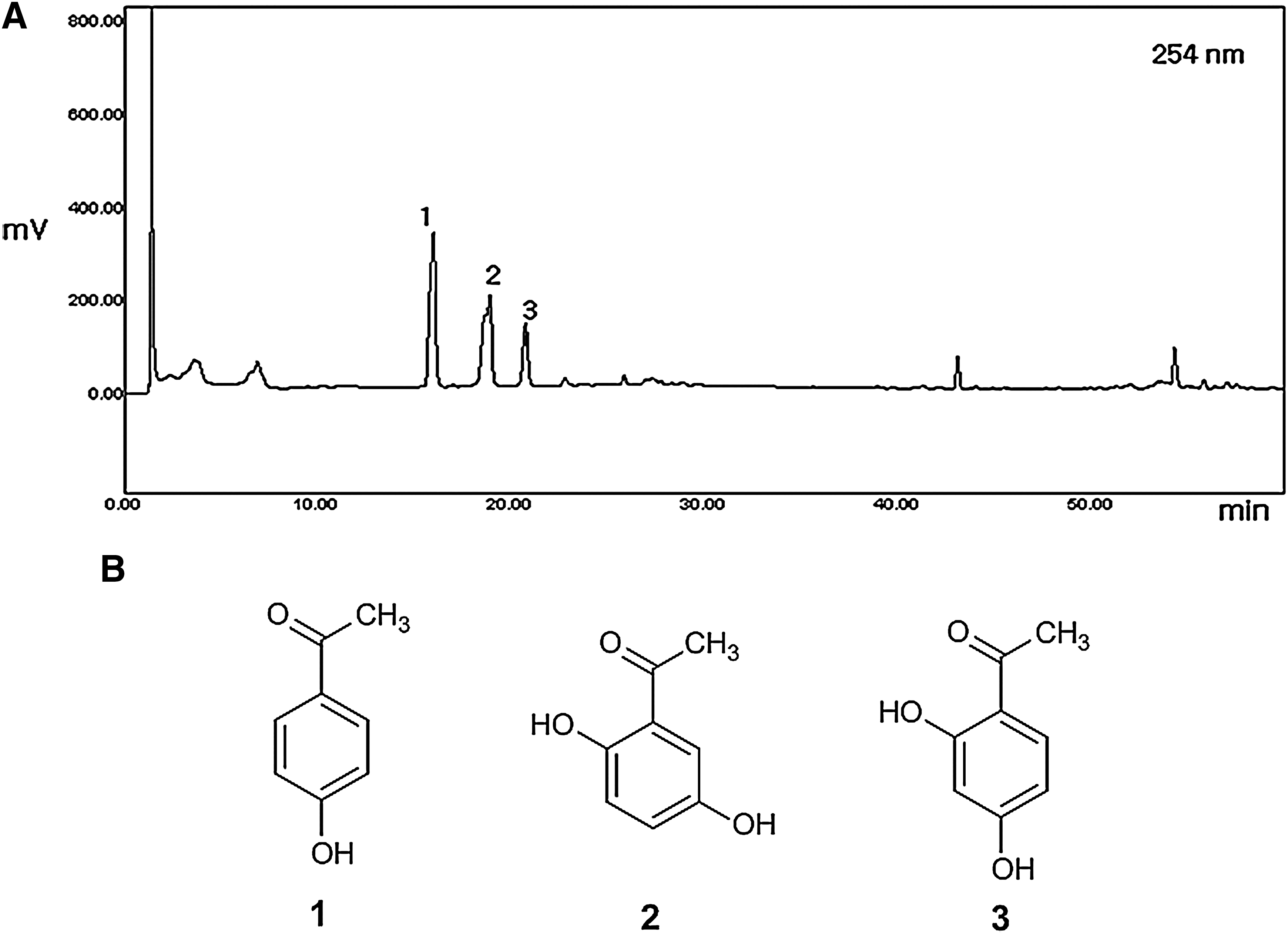
High-performance liquid chromatography profile of the ethanol extract of C. wilfordii. (
Effect of ECW on changes in body weight, lipids, and insulin and glucose levels
As shown in Table 2, HFCD-fed apoE −/− mice showed significantly increased body weight compared with RD-fed control mice, which was significantly decreased by treatment with ECW. HFCD-fed apoE −/− mice also had significantly higher plasma insulin levels compared with RD-fed mice, but treatment with ECW II resulted in significantly lower insulin levels. Plasma blood glucose levels were not statistically different between RD-fed control mice with chronic treatment of ECW.
Data are mean±SE values (n=10).
P<.05, versus control; # P<.05 versus high fat/cholesterol diet (HFCD) alone.
apoE −/−, apolipoprotein E −/−; ECW I, HFCD+100 mg/day/kg ethanol extract of C. wilfordii (ECW); ECW II, HFCD+200 mg/day/kg ECW.
Biochemical analysis of blood samples was performed to find changes in lipid metabolism in HFCD-fed apoE −/− mice (Table 3). HFCD-fed apoE −/− mice had elevated plasma LDL levels compared with their RD-fed counterparts, but treatment with ECW significantly and dose-dependently decreased plasma LDL and total cholesterol. Long-term feeding with HFCD had no effect on HDL levels; however, treatment with ECW significantly elevated HDL levels (P<.01). HFCD did not affect triglyceride levels. However, other lipid markers following non–HDL-cholesterol (defined as total cholesterol minus HDL-cholesterol) and the total cholesterol/HDL-cholesterol ratio showed that 200 mg/kg/day ECW significantly attenuated atherogenic lipoprotein particle accumulation in HFCD-fed apoE −/− mice.
Data are mean±SE values (n=10).
P<.05, ** P<.01, versus control; # P<.05, ## P<.01, versus HFCD alone.
HDL, high-density lipoprotein; HDL-c, HDL cholesterol; LDL, low-density lipoprotein; T-Cho, total cholesterol; TG, triglyceride.
Effect of ECW on changes in blood pressure
Tail cuff measurements revealed that mean systolic blood pressure was significantly increased by feeding with HFCD but was significantly decreased by chronic ECW treatment (Fig. 3).
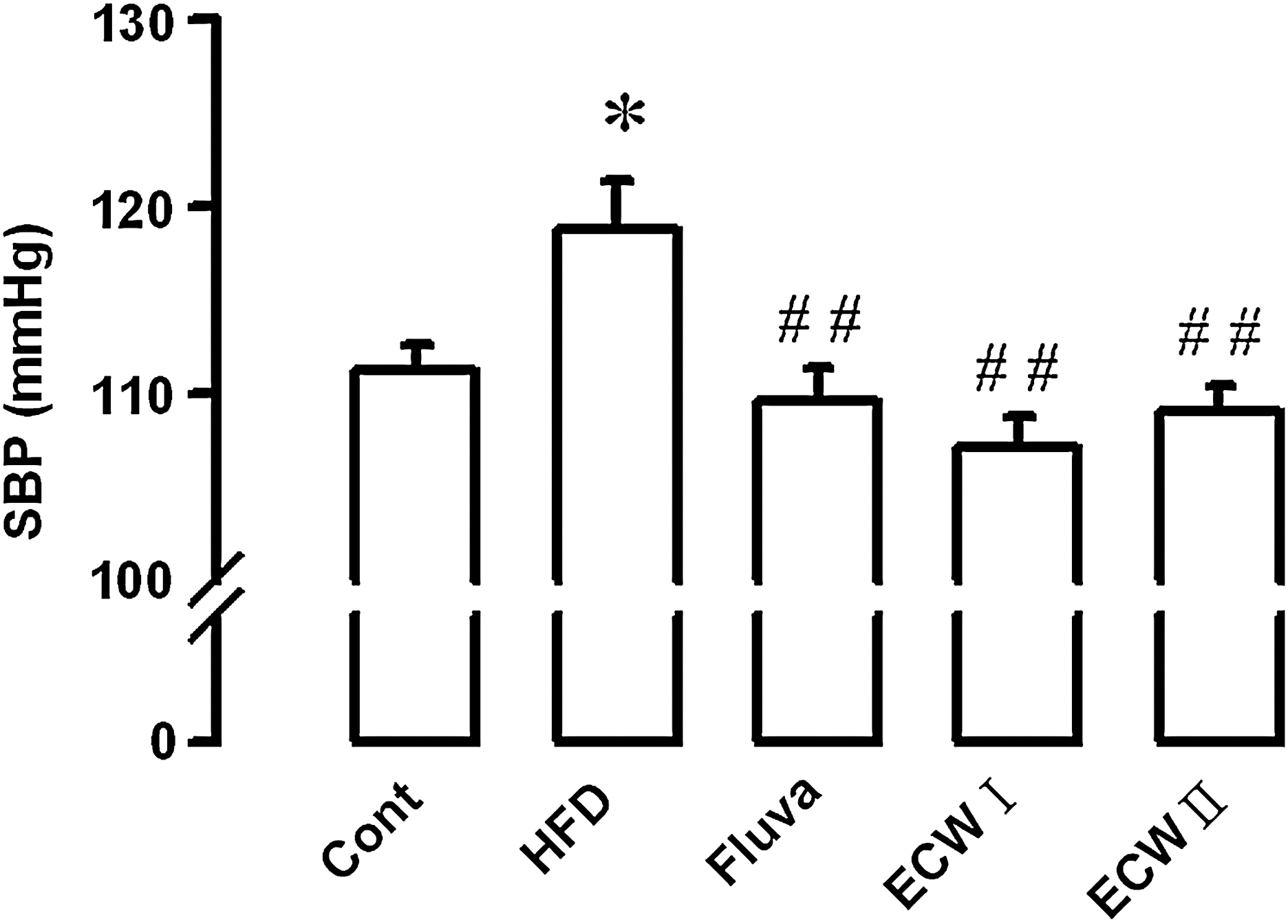
Effect of ECW on systolic blood pressure (SBP) of HFCD-fed apoE −/− mice. Data are mean±SE values (n=10). *P<.05, versus control (Cont); ## P<.01, versus HFCD alone. Fluva, fluvastatin.
Effect of ECW on microscopic changes in aorta from HFCD-fed mice
Microscopic examination of hematoxylin and eosin–stained aortic sections revealed roughened endothelial layers in HFCD-fed apoE −/− mice. HFCD-fed apoE −/− mice had significant increases of tunica intima thickness. Chronic treatment with ECW maintained the smooth and soft character of the tunica intima and decreased the intima-media thickness in aortic sections (Fig. 4).
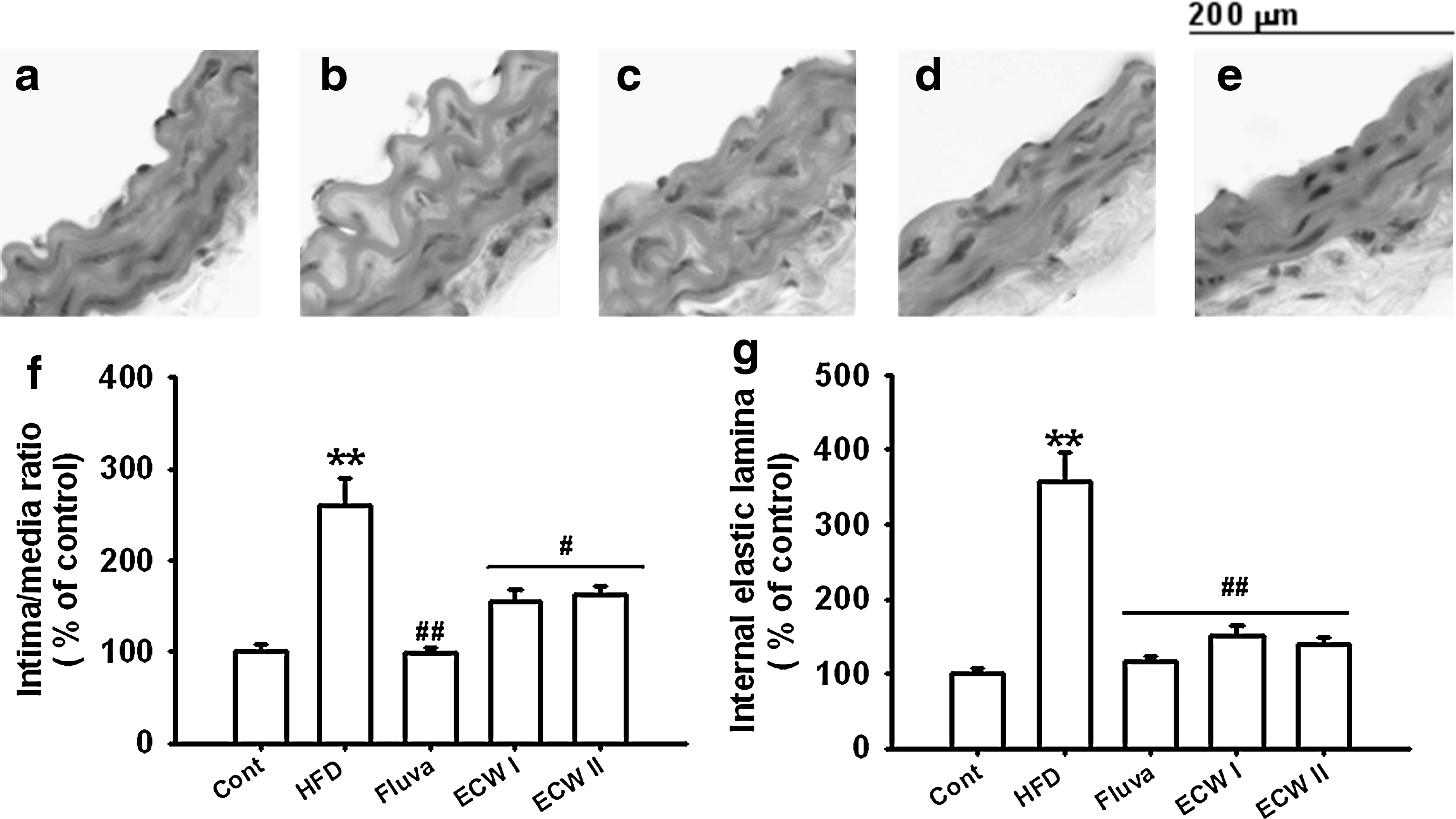
Representative microscopic photographs of hematoxylin and eosin–stained sections of the aorta of HFCD-fed apoE
−/− mice. Magnification×1,000.
Effect of ECW on changes in vascular tone of HFCD-fed mice
Vasorelaxation responses to ACh were measured in the thoracic aorta of HFCD-fed apoE −/− mice (Fig. 5a). Significant impairment of vasorelaxation was evident in thoracic aorta of HFCD-fed apoE −/− mice compared with RD-fed normal mice. Chronic ECW treatment restored the vasorelaxation response. The vasorelaxation response to sodium nitroprusside was unchanged in HFCD- and RD-fed mice. Chronic ECW treatment also did not affect the vasorelaxation response to sodium nitroprusside in HFCD-fed apoE −/− mice (Fig. 5b).
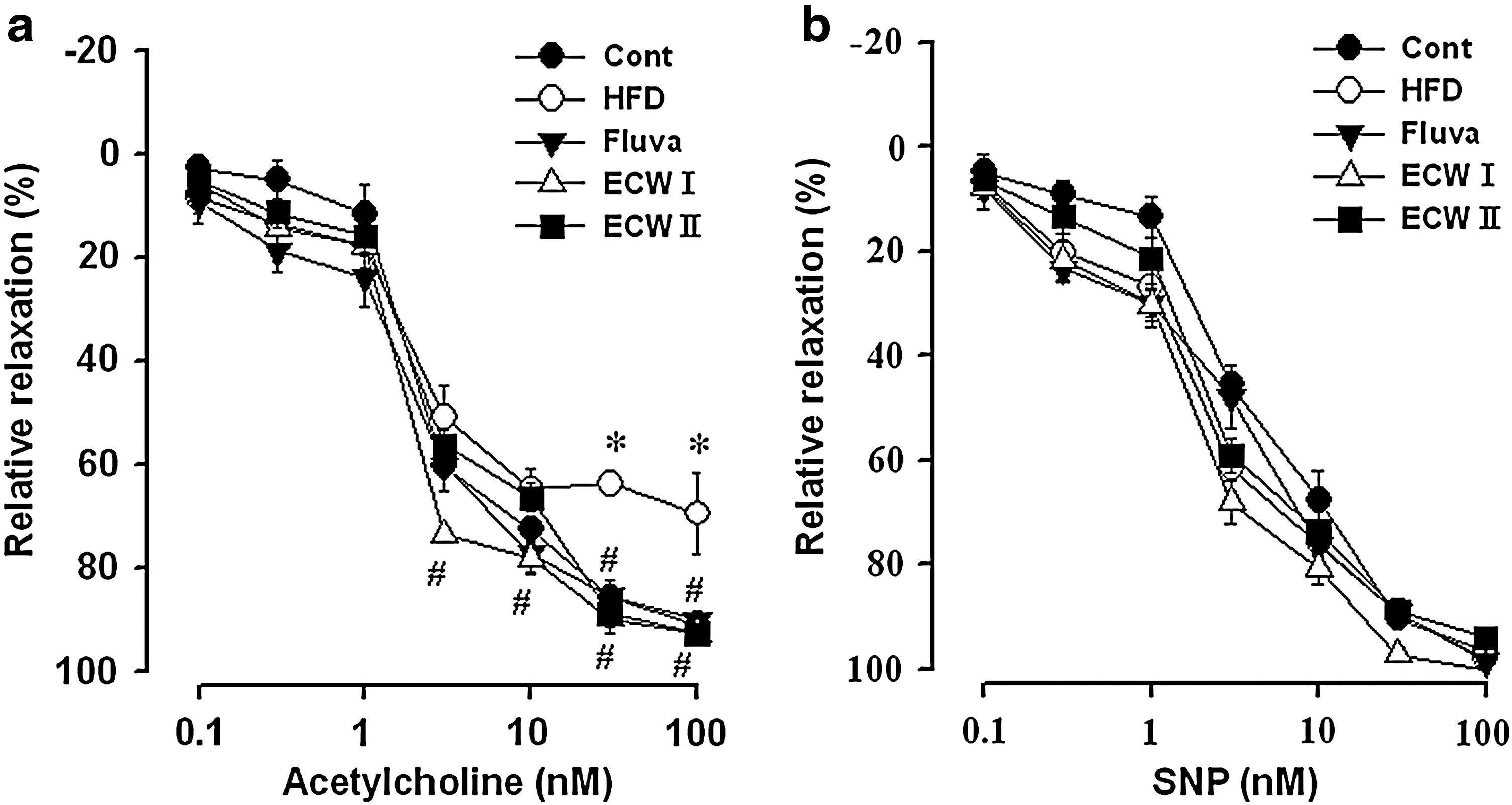
Effects of ECW on relaxation of thoracic aorta induced by
Effect of ECW on changes in eNOS expression in aorta from HFCD-fed mice
Figure 6 shows representative micrographs of eNOS expression in aortic sections of HFCD-fed apoE −/− mice using immunohistochemistry. The eNOS expression was suppressed in the aorta of HFCD-fed compared with RD-fed mice (Fig. 6a and b). Fluvastatin and ECW (200 mg/kg/day) treatment markedly restored eNOS expression levels by 182% and 223%, respectively (P<.01) (Fig. 6c and e).
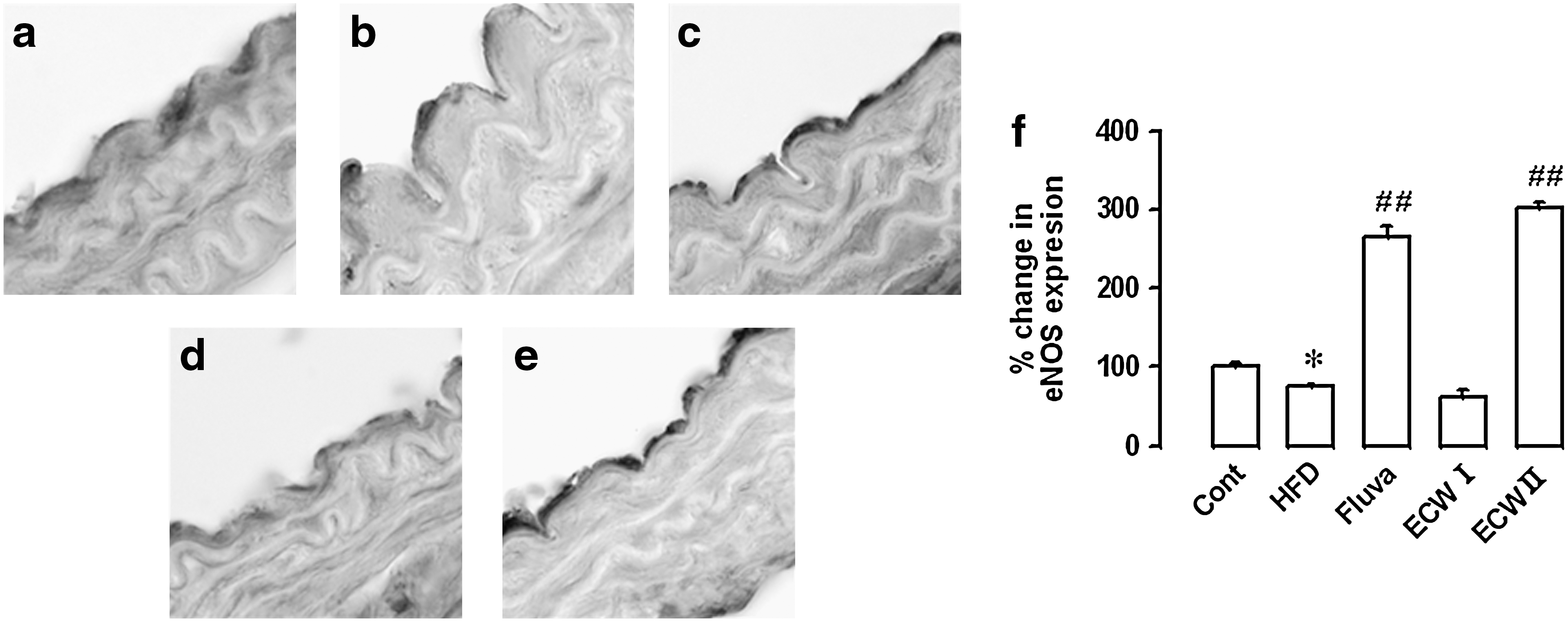
Effect of ECW on endothelial nitric oxide synthase (eNOS) expression in the aorta of HFCD-fed apoE
−/− mice:
Effect of ECW on expression of cellular adhesion molecules in aorta from HFCD-fed mice
Cellular adhesion molecule expression was evaluated quantitatively by western blot in isolated aortic tissues. Expression of E-selectin in HFCD-fed apoE −/− mice was significantly increased compared with normal mice fed RD. The group treated with 200 mg/kg/day ECW showed a lower band intensity of E-selectin compared with apoE −/− mice fed HFCD alone. The densitometric analysis also indicated that 200 mg/kg/day ECW significantly decreased E-selectin expression in comparison with apoE −/− mice fed HFCD alone. Similarly, VCAM-1 and ICAM-1 were significantly increased in HFCD-fed apoE −/− mice. Treatment with high-dose ECW significantly decreased VCAM-1 and ICAM-1 expression (Fig. 7).
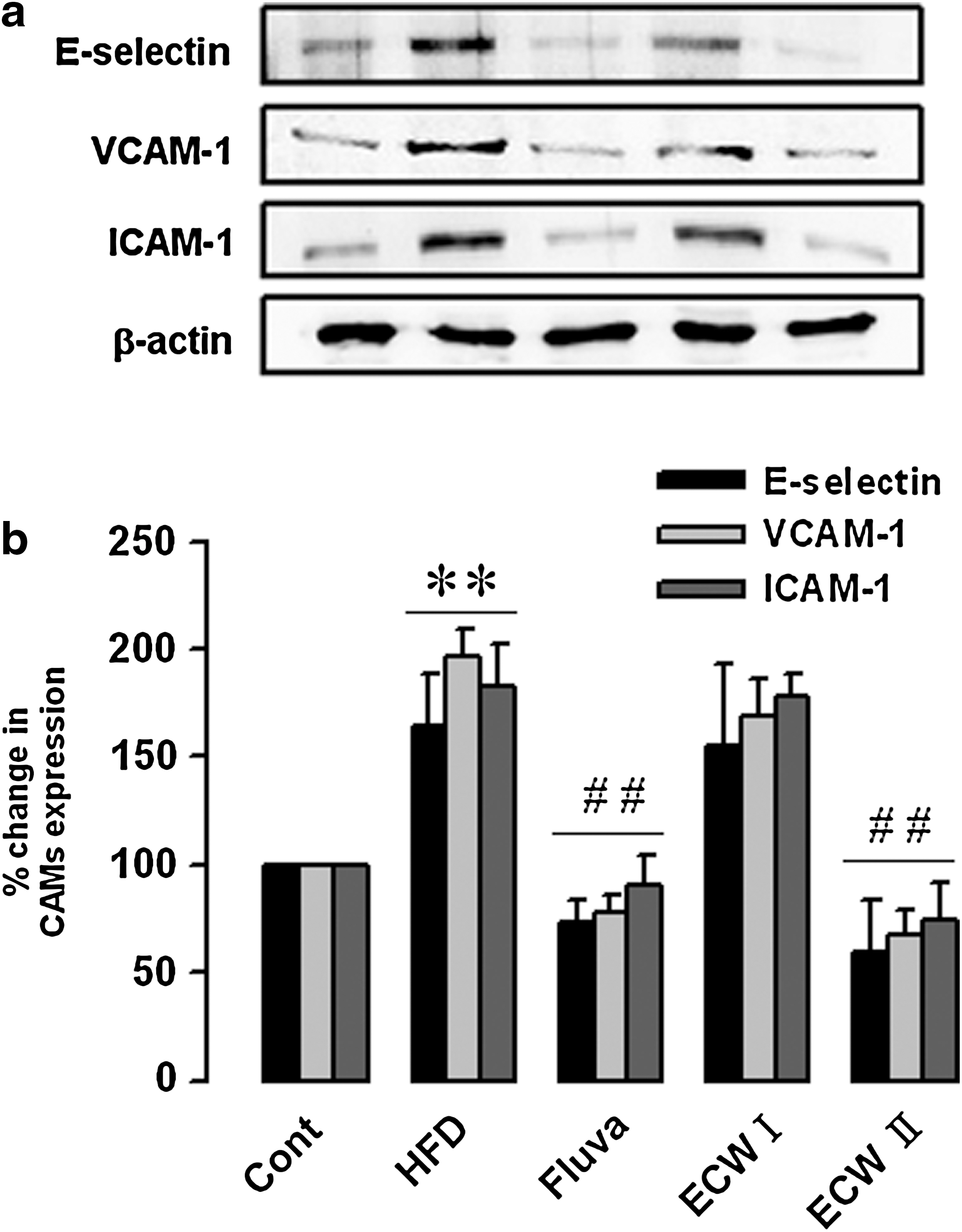
Effect of ECW on expression of cellular adhesion molecules (CAMs) in the aorta of HFCD-fed apoE
−/− mice:
Effect of ECW in ET-1 expression in aorta from HFCD-fed mice
Immunohistochemistry results showed that ET-1 expression was significantly increased in the aorta of apoE −/− mice fed with HFCD alone compared with control mice fed with RD. ECW I and II treatment markedly decreased the levels of ET-1 by 65% and 58%, respectively (Fig. 8).
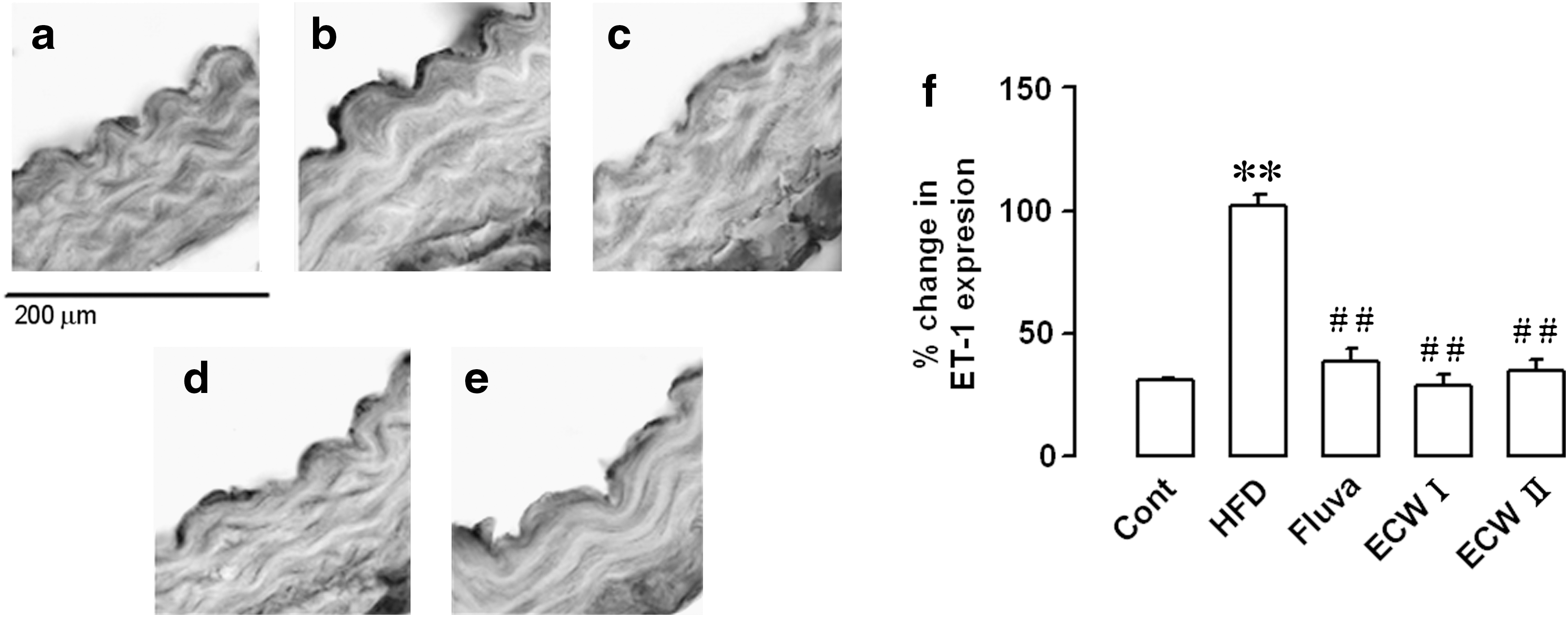
Effect of ECW on ET-1 expression in the aorta of HFCD-fed apoE
−/− mice:
Discussion
In the present study, we investigated the effects of ECW treatment on the pathophysiological changes and vascular dysfunction in HFCD-fed apoE −/− mice. Prior to the appearance of atherosclerotic changes in the vascular wall, a high cholesterol level induces vascular functional changes that may lead to vascular remodeling and increases permeability of LDL-cholesterol. 21 Elevated total and LDL-cholesterol levels impair endothelial function, and LDL-cholesterol is deposited in the blood vessel wall as part of atherosclerotic plaques. 22,23 However, increased HDL-cholesterol could suppress the atherosclerotic process by facilitating translocation of cholesterol from peripheral tissues like arterial walls to the liver for catabolism. 24,25 In apoE −/− mice, which lack the means to metabolize lipids, a fat-enriched diet can elevate the LDL-cholesterol level in plasma. 26,27 In the present study, chronic treatment with ECW significantly reduced both total and LDL-cholesterol levels and increased the HDL-cholesterol level in HFCD-fed apoE −/− mice. Increased expression of ET peptide, receptors, and converting enzyme activity has been reported in human atherosclerosis and in the aorta from apoE −/− mice with advanced lesions. Blockade of ET receptors markedly reduces lesion formation in LDL receptor–deficient mice, which is related to atherosclerosis. 28 In this study, ET-1 expression was significantly increased in the aorta of HFCD-fed apoE −/− mice; however, treatment with ECW markedly decreased the levels of ET-1. These findings, at least in part, indicate that ECW may protect against the initiation and development of atherosclerosis by improving lipid metabolism.
The vascular endothelium, which lies between circulating blood and vascular smooth muscle and senses changes or abnormalities in blood flows and pressures, plays an important role in modulation of vascular tone. 29 We found that mean systolic blood pressure was elevated by HFCD but significantly normalized by ECW treatment. In addition, HFCD also caused endothelial dysfunction as evidenced by decreased ACh-induced vascular tone. ECW exerted endothelium-dependent vasodilation in thoracic aortic smooth muscle. There were no significant differences of sodium nitroprusside–induced dilation between ECW-treated groups and the control group. These finding suggested that the hypotensive effects of ECW are mediated by ACh and further via the endothelium-dependent NO/cyclic GMP pathway. In fact, endothelial dysfunction was initially identified as impaired vasodilation to specific stimuli such as ACh or bradykinin; therefore, improvement of endothelial function is predicted to regulate lipid homeostasis. 30 Impaired relaxation of aorta is induced by ACh in obese diabetic fatty rats as a consequence of endothelial dysfunction. 31 It has been well documented that endothelium-dependent vascular relaxation is abnormal in both hypercholesterolemia and atherosclerosis because the ability of NO to maintain vascular tone is impaired. 32,33 Thus, our findings suggest a protective role of ECW on hypertension and vasoconstriction in this animal model of atherosclerosis.
Insulin resistance is associated with accelerated atherosclerosis. Impairment of NO production leads to insulin resistance such as that seen in eNOS knockout mice
34
and humans treated with the eNOS antagonist
The integrity of the internal elastic lamina may represent a barrier to cell migration and formation of intimal proliferative lesions in various models for atherosclerosis. 37 Carotid intima-media thickness of the thoracic aorta also has been shown to correlate with prognosis and extent of coronary artery disease. 38 Chronic ECW treatment maintained smooth and soft intimal endothelial layers as well as decreased intima-media thickness in aortic sections, suggesting an important role of ECW in the prevention of atherosclerosis. However, Rohani et al. 38 reported that no significant associations were seen between soluble VCAM-1 levels and intima-media thickness in the carotid and femoral arteries. Nevertheless, it is clear that endothelial dysfunction would include not only reduced vasodilation but also inflammation and atherosclerotic lesions. 30,39 Therefore, blocking of inflammatory mediators can decrease atherosclerotic lesion size. We hypothesized that the vasorelaxant effect of ECW contributes to its anti-inflammatory and anti-atherosclerotic effects. Activation of the endothelium at sites of inflammation allows numerous leukocytes to adhere to vascular endothelium and transmigrate through the endothelium, thus aggravating endothelial dysfunction and tissue injury. 27,40 Leukocyte infiltration to sites of inflammation is regulated in part by specific endothelial–leukocyte adhesion molecules, including E-selectin, VCAM-1, and ICAM-1. 41 Western blotting analysis showed that HFCD-fed apoE −/− mice increased E-selectin, VCAM-1, and ICAM-1 expression levels in thoracic aorta, and these effects were significantly prevented by 200 mg/kg/day ECW treatment. These finding suggested a potentially important role of ECW in anti-inflammatory and anti-atherosclerotic activity in hyperlipidemia and/or hypertension. Previously, we reported that Morus alba L. could improve diet-induced hypertension, hyperlipidemia, and vascular dysfunction through inhibition of cell adhesion molecule expression and induction of vascular relaxation. 42 Further study is needed to characterize the vascular protective mechanisms mediated by herbs.
The relative contribution, if any, of these effects of ECW to the enhanced eNOS and reduced ET-1 vasoconstrictor expression that we observe in aorta of HFCD-fed apoE −/− mice remains to be determined. Further experiments are also required to identify whether differences in intracellular signaling pathways activated by ECW, compared with ET-1, are sufficient to explain the apparent selective natural candidates of hypertension and/or hyperlipidemia in animal models of atherosclerosis. Blockade of ET receptors markedly reduces atherosclerosis in LDL receptor–deficient mice.
Here, we demonstrated that HFCD-fed apoE −/− mice developed severe hypertension, hypercholesterolemia, and a metabolic profile consistent with type 2 diabetes. ECW ameliorates endothelial dysfunction via improvement of the NO/cyclic GMP signaling pathway in a diet/genetic model of hyperlipidemia. In addition, ECW substantially inhibited the development of atherosclerosis, possibly by inhibiting ET-1, cell adhesion molecules, and lesion formation, suggesting a vascular protective role for this herb in the treatment and prevention of atherosclerotic vascular disease.
Footnotes
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea funded by the Ministry of Education, Science and Technology (grant number 2010-0029465) and by grant number K10040 to D.G.K. funded by the Korea Institute of Oriental Medicine.
Author Disclosure Statement
No competing financial interests exist.