
Abstract
Buckwheat is a healthy food commonly eaten worldwide. The antitumor activity of buckwheat polysaccharides (BWPSs) has not yet been evaluated. In recent years, inducing differentiation of leukemic cells has become one of the most important therapeutic approaches for curing leukemia, and this strategy effectively inhibits leukemia cell proliferation and growth because the differentiation inducer changes leukemic cell morphology and cellular characters by inducing cellular maturity. The ability of BWPS to induce the differentiation of human leukemic THP-1 cells (monocyte [MNC]/macrophage-like cells) was investigated by both direct and indirect treatments in this study. In the indirect treatment, BWPS significantly stimulated cytokine secretion (differentiation inducer) in MNCs from peripheral blood mononuclear cells in MNC-conditioned medium (BWPS-MNC-CM) following a 24-hour treatment, and THP-1 cell differentiation and maturity were significantly increased after 5 days of treatment with the BWPS-MNC-CM. On the other hand, BWPS directly induced THP-1 cell differentiation and maturity following 3-day and 5-day treatments in a dose-dependent manner and exerted phagocytic activity and superoxide anion production in these mature cells. These findings indicate that BWPS has potential for differentiation therapy in leukemia.
Introduction
Leukemia is the most common hematological malignancy. Current therapeutic options include chemotherapy, differentiation inducers, and stem cell transplantation. Among these, the strategy of differentiation induction is less toxic and safer than the other methods. 6,7 Several materials such as rice extracts, vitamin D, and retinoic acid have the ability to stimulate cytokine secretion by activating monocytes (MNCs) from peripheral blood mononuclear cells (PBMCs), including tumor necrosis factor-α (TNF-α), interferon-γ (IFN-γ), and interleukin (IL)-1β. 8 –10 Additionally, several polysaccharides isolated from edible materials have been reported to induce cytokine secretion by PBMCs, including polysaccharides from Salicornia herbacea and Cordyceps sinensis, 11,12 and these cytokines (TNF-α, IFN-γ, and IL-1β) have been found to induce differentiation of several leukemic cell lines (U937, THP-1, and K562 leukemic cells). 13
Several lines of evidence indicate that many foods stimulate immune function in addition to providing nutrition and energy, and it has been reported that black soybean, 14 rice, 8 mushrooms, 15 and Poria cocos 16 inhibit cancer cell proliferation and induce the differentiation of human leukemic cells. Moreover, some research indicates that Ganoderma lucidum can induce THP-1 cells to differentiate into MNC/macrophage- and/or MNC/dendritic-like cells, 13,17 revealing that the activity of leukemic THP-1 cells is attenuated by inducing differentiation. To our knowledge, the antitumor activity of buckwheat polysaccharides (BWPSs) is unclear. Thus, we hypothesized that possible differentiation induction of leukemic THP-1 cells by BWPS would result in THP-1 cells being transformed into MNC/dendritic- and/or MNC/macrophage-like cells. In the present study, the antileukemic activity of BWPSs through direct cytotoxic and/or indirect immune-potentiating effects was examined.
Materials and Methods
Reagents
Ficoll–Hypaque (1.077 g/mL) was purchased from Pharmacia Fine Chemicals (Piscataway, NJ, USA). Trypan blue, RPMI 1640 medium,
Preparation of polysaccharides
Seeds of buckwheat from the Fagopyrum tataricum Gaertn. species were freeze-dried and ground. Approximately 25 g of buckwheat powder was mixed with 250 mL of distilled water, and the crude polysaccharides were extracted at 95°C for 30 minutes. After extraction, the aqueous extract was concentrated under vacuum, and then the crude polysaccharides of buckwheat (BWPSs) were obtained by 95% ethanol precipitation. The collected precipitates (BWPSs) were again vacuum-concentrated and freeze-dried. The BWPS was stored at −20°C until used.
Assays for crude protein and total sugar of BWPS
Total sugar content of BWPS was analyzed by the phenol–sulfuric acid method. In brief, 0.5 mL of sample was mixed with 0.5 mL of 5% phenol dissolved in water. Sulfuric acid was added rapidly and reacted for 30 minutes at room temperature. The absorbance of the reaction was measured at 490 nm by a spectrophotometer (model U-1800, Hitachi Ltd., Tokyo, Japan), and the total sugar content was quantified in reference to a calibration curve of glucose standards. Protein content of the BWPS was quantified. In brief, 190 μL of Lowry reagent solution was mixed with 10 μL of sample. The mixture was incubated for 10 minutes at room temperature before 100 μL of Folin–Ciocalteu phenol reagent was added. The absorbance of the mixture was measured at 750 nm. The protein contents of the samples were calculated from a standard (bovine serum albumin) curve.
Preparation of PBMC-conditioned medium
PBMCs were separated from healthy human peripheral blood by density gradient centrifugation with Ficoll–Hypaque (1.077 g/mL). PBMCs were washed with phospahte-buffered saline three times, and the PBMC-conditioned medium (PBMC-CM) was obtained after PBMCs were incubated in RPMI 1640 medium supplemented 10% fetal bovine serum, 2 mM
Treatments of THP-1 cells
THP-1 cells were incubated in RPMI 1640 medium supplemented with 10% fetal bovine serum, 2 mM
Assays for cytokine levels
The levels of TNF-α, granulocyte–macrophage colony-stimulating factor (GM-CSF), and IL-1β in PBMC-CM and the levels of IL-10, IL-12, TNF-α, and IL-1β in THP-1 culture medium were determined with commercial enzyme-linked immunosorbent assay kits from R&D Systems (Minneapolis, MN, USA).
Differentiation induction
After 5 days of differentiation induction with BWPS, the cells were collected and stained with Liu's stain dye. Cell morphology was observed under an inverted microscope with a magnification of ×1,000. Based on morphology, the cells were classified as (1) immature blasts, (2) intermediate, and (3) mature MNC/macrophage- or MNC/dendritic-like cells by the methods of Hsu et al. 13 and Chan et al. 17
Assay for phagocytosis
After 5 days of differentiation induction with BWPS or PBMC-CM, the phagocytic activity was measured by the method of Liao et al. 8 The cells were washed with phosphate-buffered saline, resuspended in medium (8 × 105/mL) at day 5, and then incubated with yeast (1.0 × 106/mL) at 37°C for 30 minutes. Finally, cells were placed on a glass slide and observed to determine phagocytic activity. Phagocytosis (as a percentage) was quantified by counting the percentage of yeast-containing cells from 200 cells.
Assay for superoxide anion level
The level of reactive oxygen species was assayed with NBT. In brief, THP-1 cells (1.5 × 105/mL) were treated with PBMC-CM or BWPS for 5 days, and then cells were collected and resuspended in medium (8 × 105/mL). The cells were then incubated with NBT solution containing 1 μM phorbol myristate acetate at 37°C for 2 hours. NBT is reduced by reactive oxygen species to form blue-black formazan, which was dissolved with dimethyl sulfoxide, and the absorbance value was measured at 570 nm. 8 The level of reactive oxygen species generation was compared to that of the normal group.
Assay for specific antigen markers
The specific differentiation marker CD11b and CD14 in cells were determined by flow cytometric analysis as previously described. 13 In brief, THP-1 cells (1.0 × 105/mL) were treated with PBMC-CM and BWPS for 5 days, and then cells were washed with phosphate-buffered saline and incubated with phycoerythrin-conjugated anti-human CD11b and CD14 antibody (BD Biosciences PharMingen, San Diego, CA, USA) for 1 hour. Redundant antibody was removed by washing with phosphate-buffered saline. The expressions of CD11b and CD14 were analyzed by the flow cytometry.
Statistical analysis
Data are expressed as mean ± SD values. Analysis of variance was used to evaluate the differences between multiple groups. If significance was observed between multiple groups, Duncan's multiple range test was used to compare the means of two specific groups. P < .05 was considered to be significant.
Results
Cytotoxicity of PBMCs
The cytotoxicity of BWPS for PBMCs is shown in Figure 1. No significant cytotoxic effects on PBMCs were observed with BWPS treatment for 24 hours (50–200 μg/mL). No significant agglutination of PBMCs was triggered after treatment with BWPS for 24 hours. In contrast, PHA (20 μg/mL) significantly induced agglutination of PBMCs (data not shown). These results showed that BWPS is a safe substance that can be used for stimulating PBMC activity.
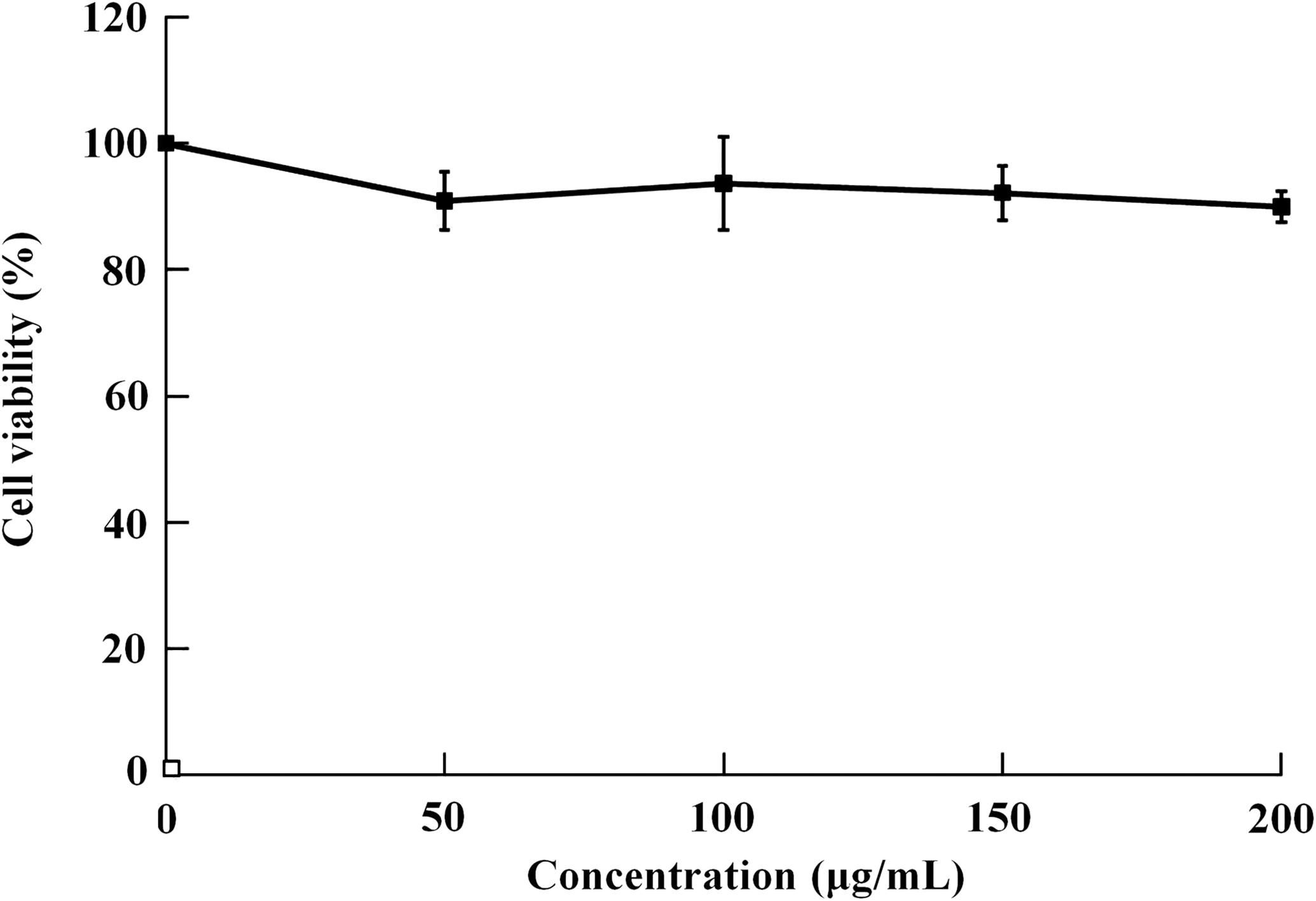
Cell viability of peripheral blood mononuclear cells after treatment with various concentrations of buckwheat polysaccharide (50, 100, 150, and 200 μg/mL) for 24 hours (n = 3). The inhibitory activity of phytohemagglutinin (20 μg/mL) was 24.3%.
Proliferation inhibition on human leukemia THP-1 cells
The growth inhibitory effects of THP-1 cells by direct and indirect BWPS treatments were investigated. Proliferation of THP-1 cells was significantly inhibited by direct treatment with BWPS for 3 days (Fig. 2A). The cell viability was lowest (89.2%) on day 3 of BWPS treatment (200 μg/mL). Furthermore, the inhibition rate of THP-1 cell proliferation appeared to be dose dependent; the highest level of inhibitory activity for BWPS was 32.6% after induction for day 5. On the other hand, BWPS-PBMC-CM also exerted inhibitory activity on THP-1 proliferation on day 1 (Fig. 2B). Moreover, the inhibitory activity of BWPS (200 μg/mL)-PBMC-CM on THP-1 was significantly greater than that of BWPS (200 μg/mL) on days 3 and 5. The cell viabilities were 67.5% and 63.2% with 150 μg/mL and 200 μg/mL of BWPS-PBMC-CM on day 5, respectively. The PHA (20 μg/mL)-PBMC-CM inhibited 51.4% of THP-1 cell proliferation on day 5. These results revealed that BWPS efficiently stimulated immunity by affecting human PBMCs; therefore the PBMC-CM was capable of differentiation induction.

Cell viability of THP-1 cells assessed by direct and indirect treatments for 1, 3 and 5 days, respectively. (
Cytokine levels in PBMC-CM and THP-1 culturing medium
The levels of IL-1β (487.6 pg/mL) and TNF-α (1220.4 pg/mL) production were significantly increased in a dose-dependent manner after treatment with BWPS (200 μg/mL) in PBMC-CM for 24 hours (Table 1). The two cytokines both induced THP-1 differentiation into MNCs/macrophages. GM-CSF is a major inducer for MNC/dendritic cell differentiation, and the level of GM-CSF (88.6 pg/mL) was not evidently elevated in PBMC-CM compared to the normal group by BWPS (200 μg/mL) treatment. These results indicated that BWPS may exert the differentiation potentials of MNC/macrophage cells through PBMC activation because BWPS increased PBMCs' secretion of IL-1β and TNF-α, but not of GM-CSF. On the other hand, TNF-α and IL-1β are produced and secreted by mature MNCs and/or macrophages, and IL-10 and IL-12 are generated by mature dendritic cells. The levels of TNF-α (687.4 pg/mL) and IL-1β (360.1 pg/mL) were both increased in THP-1 cells treated with BWPS (200 μg/mL) for 5 days (Table 2). These results revealed that BWPS itself may have potential for stimulating differentiation of THP-1 cells into MNCs/macrophages.
Peripheral blood mononuclear cells were treated with various concentrations of buckwheat polysaccharide (BWPS) and phytohemagglutinin (PHA) (20 μg/mL) for 24 hours. Cells in the normal group were not treated with BWPS or PHA. Data are mean ± SD values (n = 3).
GM-CSF, granulocyte–macrophage colony-stimulating factor; IL-1β, interleukin-1β; ND, not determined; PBMC-CM, peripheral blood mononuclear cell–conditioned medium; TNF-α, tumor necrosis factor-α.
THP-1 cells were treated with various concentrations of BWPS and PHA (20 μg/mL) for 5 days. Normal cells were not treated with BWPS or PHA. Data are mean ± SD values (n = 3).
Degree of differentiation of THP-1 cells
As demonstrated in Table 3, BWPS-PBMC-CM triggered the differentiation of immature blast THP-1 cells into mature MNCs and macrophage cells (Fig. 3). After differentiation induction, nucleolus shrinkage of THP-1 differentiating cells (mature MNCs or macrophages) is found, and the morphology of differentiating THP-1 cells appears elliptical or scrambled in shape, and the pseudopod of the macrophage is observed. 13,17 The percentage of immature THP-1 cells markedly decreased from 96.4% (normal PBMC-CM) to 34.8% in the BWPS (200 μg/mL)-PBMC-CM group, and the percentage of mature MNC/macrophage-like cells increased from 2.4% (normal PBMC-CM) to up to 54.3% (BWPS [200 μg/mL]-PBMC-CM); these findings revealed the potential of BWPS to induce differentiation through activation of PBMCs. In contrast, THP-1 cell differentiation induction was 58.4% in the PHA (20 μg/mL)-PBMC-CM treatment group. Therefore, BWPS produced a weaker increase in differentiation induction of THP-1 after a 5-day treatment than BWPS-PBMC-CM. The percentages of differentiation induction were 26.7% and 40.2% in the 150 μg/mL and 200 μg/mL BWPS treatment groups, respectively.
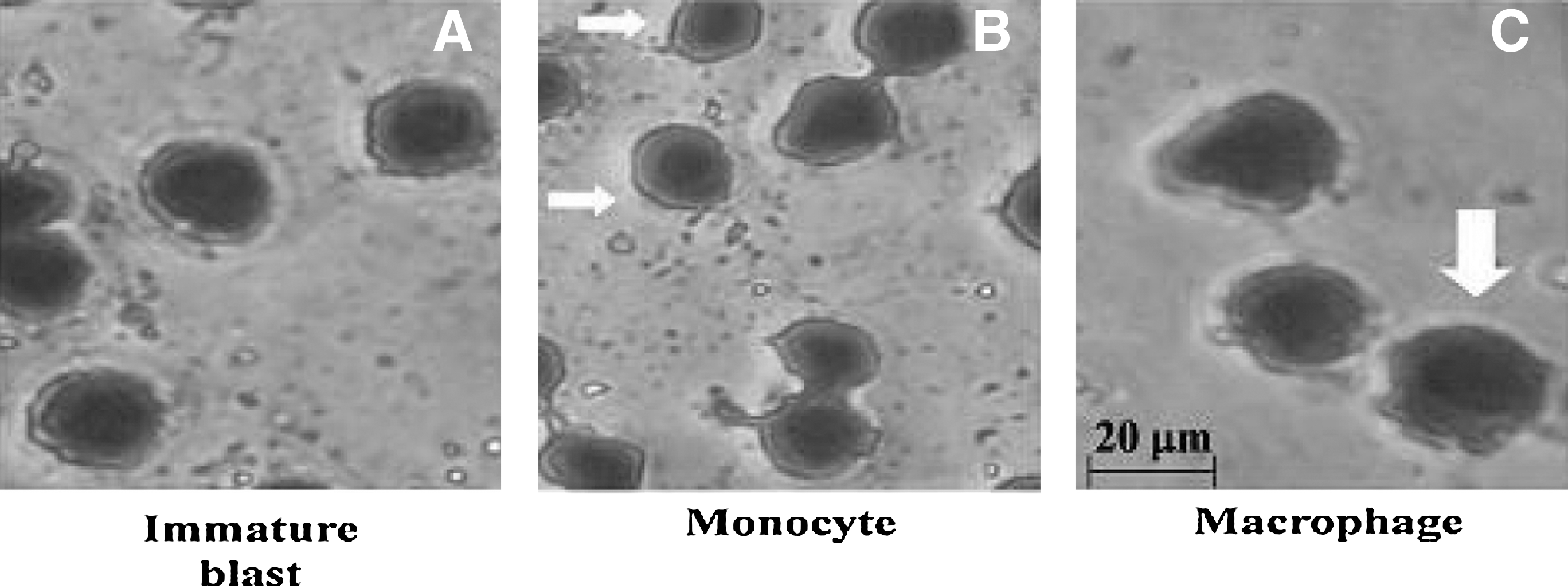
Morphological changes of THP-1 cells: (
Data are mean ± SD values (n = 3). Cell morphology was observed by Liu's stain dye.
For direct induction, THP-1 cells were treated with various concentrations of BWPS and PHA (20 μg/mL) for 5 days. Normal cells were not treated with BWPS or PHA.
For indirect induction, PBMCs were treated with various concentrations of BWPS and PHA (20 μg/mL) for 24 hours, and the PBMC-CM (35%) was collected to treat THP-1 cells for 5 days. Normal cells were treated with normal PBMC-CM for 5 days.
NBT reduction test
Stimulation of activated MNC/macrophage-like cells by phorbol myristate acetate resulted in superoxide anion generation. The level of superoxide anion was assayed with NBT. NBT was reduced to form blue-black formazan by the generated superoxide anion and could be dissolved in dimethyl sulfoxide. We investigated the activity of MNC/macrophage-like cells in generating superoxide anion; the results are shown in Figure 4. Immature THP-1 cells generated very little superoxide anion, but treatment with BWPS-PBMC-CM resulted in a marked increase in the percentage of superoxide anion. The effect of BWPS (200 μg/mL)-PBMC-CM on superoxide anion was greater than that of BWPS (58.7% and 36.5%, respectively).
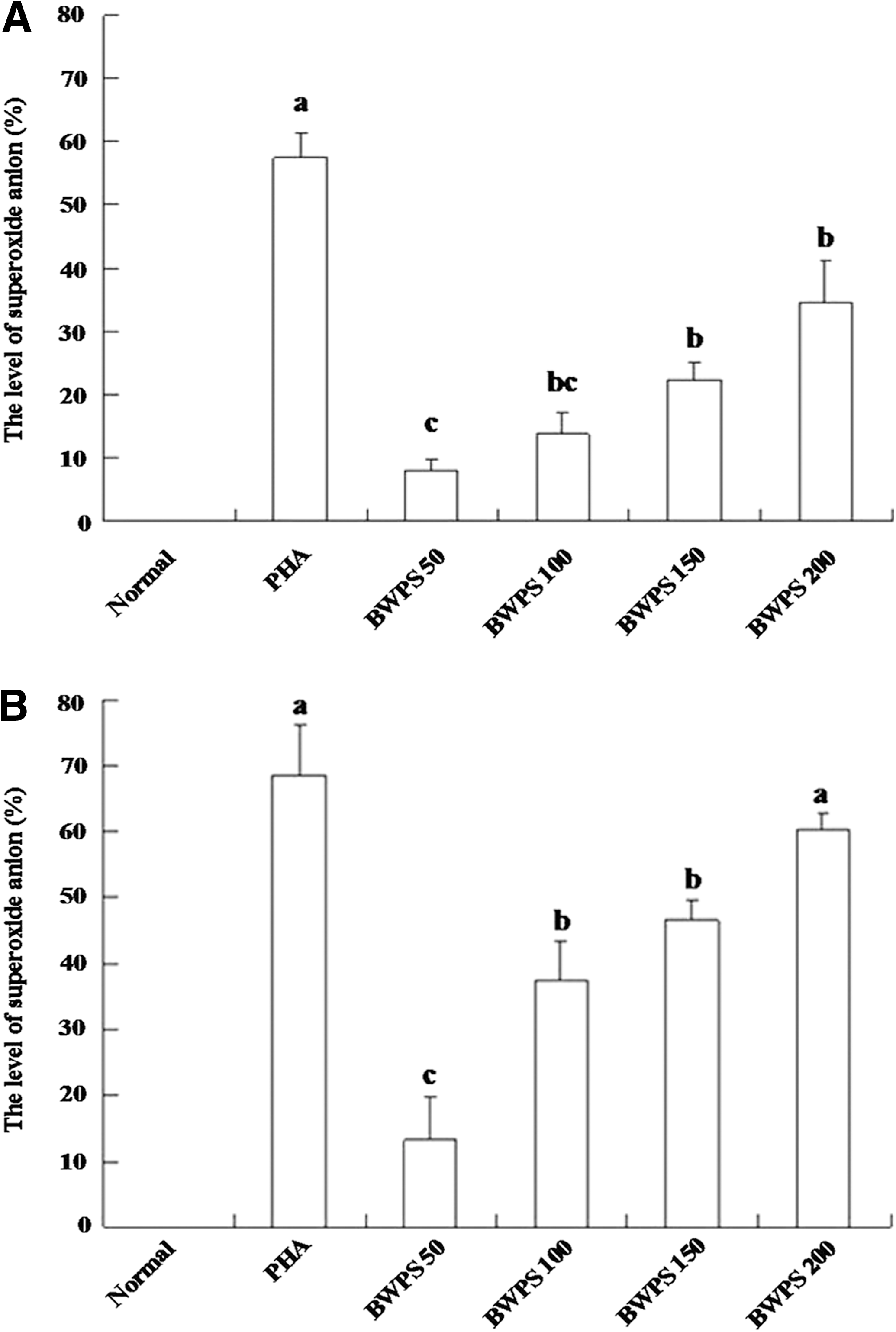
Levels of superoxide anion production in differentiating THP-1 cells by direct and indirect treatments at day 5 (n = 3). (
Phagocytic activity
Phagocytosis is a functional index of mature MNC/macrophage-like cells. Untreated THP-1 cells exhibited no significant phagocytotic activity against yeast, but treatments with BWPS-PBMC-CM and BWPS obviously increased this activity (Fig. 5). The percentage of phagocytosis was 47.4% in the BWPS (200 μg/mL)-PBMC-CM group, which was greater than that in the BWPS-treated group (27.6%).
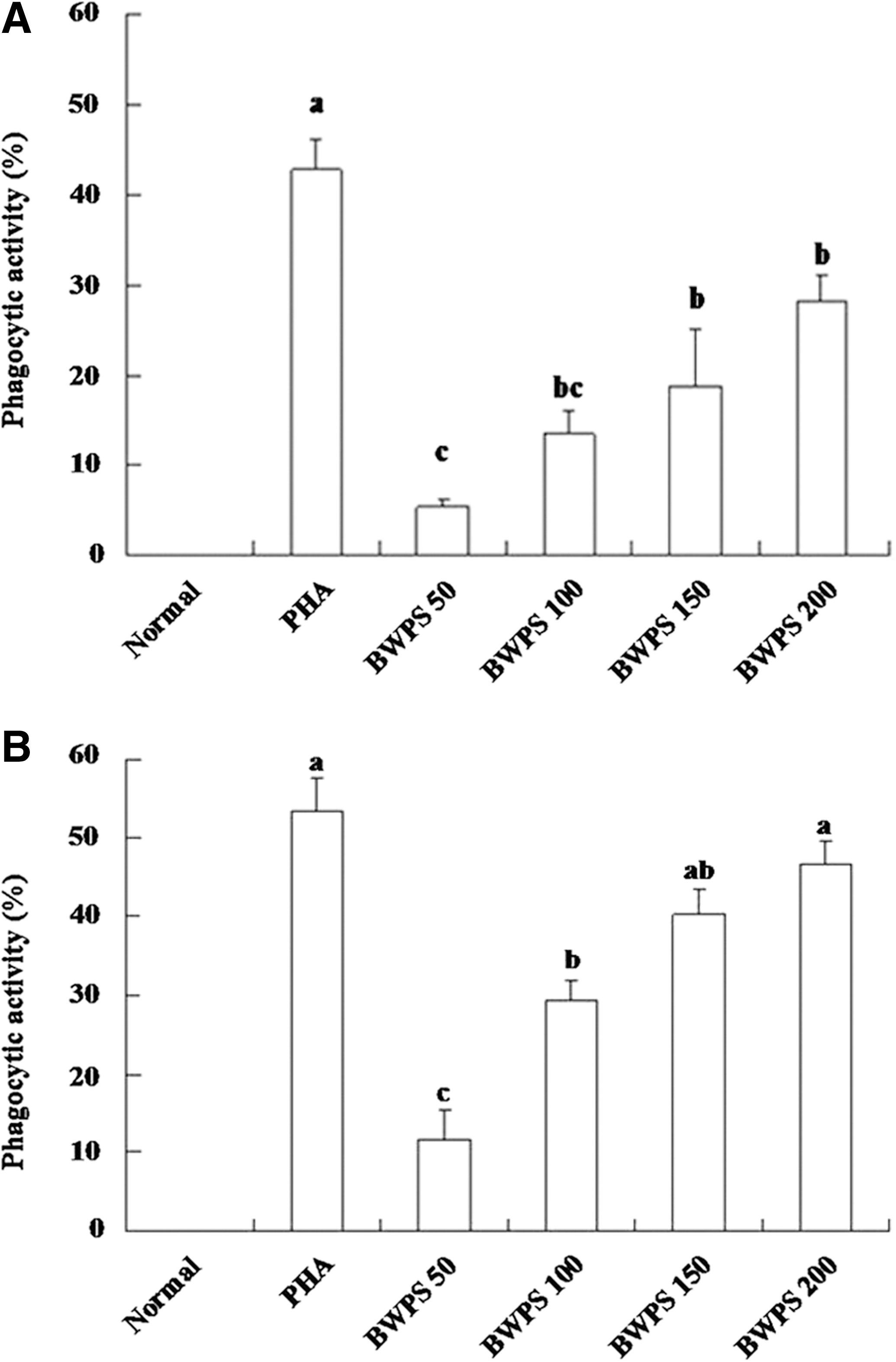
Phagocytic activity of differentiating cells by direct and indirect treatments at day 5 (n = 3). (
Expression of specific differentiation markers
Specific cell markers related to macrophage differentiation were investigated, and the results are shown in Table 4. There were 56.8% and 74.2% elevations in levels of CD11b and CD14 antigens in the group treated with PHA for 5 days. In comparison, the expressions of CD11b and CD14 were significantly increased by 47.2% and 68.5%, respectively, in the BWPS (200 μg/mL) treatment group. On the other hand, PHA-PBMC-CM markedly induced expression of CD11b (64.3%) and CD14 (91.1%) antigens. Similar results were found in the BWPS (200 μg/mL)-PBMC-CM group: CD11b and CD14 expression were increased by 78.7% and 87.4%, respectively. These findings demonstrated that BWPS can induce THP-1 differentiation by activating PBMCs.
Data are mean ± SD values (n = 3).
For direct induction, THP-1 cells were treated with various concentrations of BWPS and PHA (20 μg/mL) for 5 days. Normal cells were not treated with BWPS or PHA.
For indirect induction, PBMCs were treated with various concentrations of BWPS and PHA (20 μg/mL) for 24 hours, and the PBMC-CM (35%) was collected to treat THP-1 cells for 5 days. Normal cells were treated with normal PBMC-CM for 5 days.
Discussion
The potential of lipopolysaccharide for differentiation induction has been previously demonstrated.
18
To rule out possible endotoxin contamination in the BWPS-PBMC-CM and BWPS, polymyxin B (50 μg/mL) was added to evaluate the inhibition and differentiation of THP-1 cells. Polymyxin B is well known as a lipopolysaccharide activity-blocking agent. The aforementioned differentiation-inducing and proliferation-inhibitory effects of THP-1 cells by BWPS-PBMC-CM or BWPS were not blocked by polymyxin B treatment (Supplementary Fig. S1 and Supplementary Table S1; Supplementary Data are available online at
BWPS is nontoxic and safe for PBMCs (Fig. 1) and may potentially be useful in stimulating PBMC activity. Furthermore, treating with either BWPS-PBMC-CM or BWPS inhibited THP-1 cell proliferation by indirect or direct strategies (Fig. 2). These results suggested that BWPS might show potentials for attenuating morbid symptoms caused by leukemia. There are two possible ways for BWPS to inhibit THP-1 cells: (1) directly repressing cell proliferation and (2) stimulating secretion of differentiation-inducing cytokines from PBMCs to indirectly inhibit THP-1 cells. BWPS-PBMC-CM markedly inhibited THP-1 cell proliferation, and the finding was attributed to the action of BWPS through triggering the secretion of TNF-α and IL-1β from PBMCs (Table 1) and making these cytokines capable of differentiation induction (MNC/macrophage-like differentiation). In contrast, BWPS did not significantly elevate PBMC secretion of GM-CSF in BWPS-PBMC-CM. Results of this study suggest that G. lucidum polysaccharide stimulates MNC/dendritic-like differentiation of THP-1 leukemic cells by increasing GM-CSF secretion from PBMCs, indicating that GM-CSF is an important inducer of MNC/dendritic-like cell differentiation. 17 However, significant secretion of GM-CSF from PBMCs by BWPS treatment was not found (Table 2), and MNC/dendritic-like cell differentiation of THP-1 cells was not observed with BWPS-PBMC-CM or BWPS treatment. These findings suggested that THP-1 cells were induced to differentiate into MNC/macrophage-like cells by TNF-α and IL-1β in BWPS-PBMC-CM (indirect induction). Furthermore, although BWPS also induced differentiation of THP-1 cells into MNC/macrophage-like cells, the differentiation activity was lower than in BWPS-PBMC-CM (Table 3). Treating with BWPS and BWPS-PBMC-CM elevated both differentiation markers (CD11b and CD14) of MNC/macrophage-like cells (Table 4). An increase in differentiating induction activity by BWPS-PBMC-CM (indirect treatment) was greater than with BWPS (direct treatment) treatments, revealing the antileukemic activity of BWPS-PBMC-CM was stronger than that of BWPS, which contributed to the stimulatory response of BWPS for PBMCs, and this activity may be caused by the synergistic actions of BWPS and differentiating cytokines.
Polysaccharides of G. lucidum, Grifola umbellate, and Cordyceps ophioglossoides contain β-(1,3)- and α-(1,6)-
BWPS contained a bit of crude protein (32 mg/g) and a lot of sugar (964 mg/g) as determined by the phenol–sulfuric acid and Folin phenol reagent methods. This result indicates that high-quality polysaccharide (BWPS) can be identified and obtained from buckwheat. The findings noted here demonstrated that BWPS has the potential to stimulate the secretion of cytokines from PBMCs to induce leukemic THP-1 cell differentiation. Another study found that polysaccharide of G. lucidum (reishi) directly stimulated differentiation of THP-1 cells via activating Toll-like receptors. 25 BWPS induction of MNC/macrophage-like cell differentiation in THP-1 cells might be also mediated through activation of Toll-like receptors. And, the cytokines IL-1β and TNF-α were significantly increased in differentiating cells following BWPS treatment (Table 2). Moreover, the activation of PBMCs (TNF-α, IFN-γ, and IL-1β secretion) by S. herbacea polysaccharide stimulation has also been observed. 11 Our results demonstrated that BWPS stimulated PBMCs to release IL-1β and TNF-α (Table 1).
Several studies associating differentiation induction with particular differentiation mechanisms have recently been reported. The protein c-Jun N-terminal kinase activity is necessary for vitamin D-induced leukemia cell differentiation. 26 In addition, extracellular signal-regulated kinase-1 and -2 inactivation and p38 mitogen-activated protein kinase activation also mediate differentiation of K562 leukemia cells. 27 Although the differentiation-inducing potential of BWPS was found in this study, the signaling mechanism of differentiation induction should be further investigated.
In summary, stimulation of MNCs by both BWPS and BWPS itself inhibits human THP-1 leukemia cell proliferation and induces differentiation by day 5 of treatments. We currently plan to construct an in vivo model by an implantation of tumor cells to evaluate the effects of BWPS on induction of leukemia differentiation and inhibition of cancer cell proliferation via stimulating host immune responses.
The structural component of the polysaccharide responsible for the activity of BWPS remains unidentified and should be further investigated. Moreover, BWPS may be digested through intestinal passage; whether this digestion is beneficial to bind to gastrointestinal epithelial cells to trigger mucosal immunity and enter the circulation to activate PBMCs needs to be elucidated.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.