
Abstract
In a preliminary study we found that incubating raw ginseng in oil facilitated autolysis and extensive metabolism of ginseng, releasing flavor and lipophilic compounds into the oil so that it could be used as an ingredient for high value-added foods, while the residue could be utilized for making ginseng extract. Here, we report the effect of oil (grapeseed oil [GSO]) extract of ginseng (OEG) on body weight gain and lipid metabolism in a mouse model. OEG, but not GSO, inhibited porcine pancreatic lipase. Plasma triglyceride (TG) levels were lower in male ICR mice at 1, 2, 3, and 4 hours after oral administration of the lipid emulsion plus OEG (1 g/kg of body weight) than in the group administered only the lipid emulsion or lipid emulsion plus GSO. Next, male C57BL/6J mice were fed a standard diet, a high fat (HF) diet containing 30% lard, or diets including 30% OEG or GSO based on the standard diet for 14 weeks. Consumption of OEG-containing diet significantly lowered the body weight gain, feed efficiency, visceral fat accumulation, plasma TG, and hepatic and white epididymal adipocyte sizes, as well as expression of peroxisome proliferator-activated receptor γ (PPARγ) in liver and adipose tissue. In conclusion, dietary OEG improved obesity-related parameters in blood, liver, and adipose tissue in a mouse model and suppressed obesity induced by HF diet, possibly by regulating lipid metabolism by modulating PPARγ protein expression.
Introduction
O
Recently, numerous strategies, including diet therapy, exercise, and drugs for controlling lipid digestion or appetite, have been proposed to treat obesity. However, many antiobesity measures have side effects because they alter the biochemistry of metabolism or control appetite. 8 Some medicinal herbs are believed to have beneficial effects on obesity, blood pressure, blood glucose concentration, and hyperlipidemia with minimal side effects. 4,9 However, the majority of these plant extracts have yet to be scientifically evaluated.
Ginseng is a widely used medicinal herb in Asia, with demonstrated benefits including antihyperglycemia 10 and inhibition of lipid peroxidation in many model systems, 11 improvement of immune function, 12 and antiobesity effects mediated by regulation of peroxisome proliferator-activated receptor (PPAR) γ. 13 –16 The primary bioactive components in ginseng are ginsenosides, polysaccharides, polyacetylenic alcohol, fatty acids, peptides, and essential oils. 17 A large number of studies have focused on ginsenosides, which are the saponins of ginseng, exhibiting immunomodulation, 18 cardiovascular protection, 19 and antitumor effects. 20 Moreover, it has been shown that ginseng's volatile oil inhibits gastric cancer cell growth by regulating metabolism of DNA, carbohydrates, and energy. 21
Plant-derived essential oils are rich sources of volatile terpenoids and phenolic compounds. The essential oil and lipid-soluble compounds are known to have potential to prevent obesity and have been used in aromatherapy for obese middle-aged women. Some volatile compounds extracted from plants may have antioxidant activity that could mitigate obesity-related complications, including atherosclerosis and some cancers. 22 –25
Oil extract of ginseng (OEG) used in this study was prepared by immersing sliced raw ginseng in grape seed oil (GSO) for 1 month at 30°C. Our preliminary study showed that raw ginseng underwent extensive autolysis probably by endogenous enzymatic action and/or fermentation by facultative anaerobic microbes on the surface of ginseng while being immersed in oil, producing various metabolites and releasing lipophilic compounds into the oil medium. Thus, OEG prepared in this way not only gives off a strong ginseng fragrance but also can contain numerous bioactive components derived from ginseng. Moreover, immersing ginseng in oil resulted in extensive changes in the metabolite profile of raw ginseng saponins and increased some forms of saponins (e.g., Rg3, Rh). Whereas some harmful compounds such as polyaromatic hydrocarbons can be generated during classical red ginseng processing, the current processing may produce a new ginseng product with health benefits similar to those of red ginseng but without the concern for polyaromatic hydrocarbons. A further advantage of this processing is that ginseng oil, a by-product, could be used as high value-added vegetable oil for cooking, salad dressing, and even cosmetics because of the ginseng flavor and bioactive functions.
The extended incubation of sliced ginseng in GSO resulted in the production of numerous volatile and/or lipophilic compounds as well as extensive metabolism of saponins. The most abundant compounds in ginseng essential oil were butanal, hexanal, pinenes, myrcene, ethanol, acetone, and acetic acid. 26 –28
The present study analyzed the bioactive components in OEG and their in vivo and in vitro antiobesity effects.
Materials and Methods
Preparation of oil extract of ginseng
Freshly harvested white ginseng (4 years old) was obtained from local farmers in Kumsan, Republic of Korea and fermented in the laboratory as previously described with slight modifications. 29 In brief, after washing with tap water and air-drying for 12 hours at room temperature, each root was sliced into four parts and incubated in GSO at 30°C for 30 days. The liquid portion (OEG) was separated from the residue and used for the study.
Extraction of volatile compounds from samples
The GSO and OEG were subjected to solid-phase microextraction to recover volatile flavor compounds. They were placed in 15-mL headspace vials specially designed for solid-phase microextraction, and the vials were sealed prior to extraction. The extraction was performed by agitating the vials at 250 rpm and 70°C for 15 minutes. Solid-phase microextraction fiber composed of polydimethylsiloxane-divinylbenzene (diameter, 65 μm) was used. The desorption of the volatile compounds was performed at the injection port for 1 minute.
Analysis of volatile compounds
The volatile compounds absorbed on solid-phase microextraction were analyzed by gas chromatography/mass spectrometry (GC-3800/Saturn 2200, Varian, Palo Alto, CA, USA). The conditions for gas chromatography/mass spectrometry analysis are shown in Table 1.
Pancreatic lipase inhibitory activity
The inhibitory activity of sample against pancreatic lipase activity was determined by comparing the releasing rate of fluorescent 4-methylumbelliferone from 4-methylumbelliferyl oleate in the absence and presence of sample. 30 In brief, the reaction mixture consisted of 0.1 mL of 0.1 mM 4-methylumbelliferyl oleate, 0.04 mL of a McIlvane buffer (0.1 M citrate–Na2HPO4, pH 7.4), and 0.01 mL of a sample solution. Addition of 0.05 mL of lipase, all in a final volume of 0.2 mL, started the reaction. After incubation at 37°C for 20 minutes, 1 mL of 0.1 N HCl and 2 mL of 0.1 M sodium citrate were consecutively added. The amount of 4-methylumbelliferyl released by the lipase was measured fluorometrically at an excitation wavelength of 320 nm and an emission wavelength of 450 nm. The inhibitory activity (percentage) was calculated as (1 – A/B)×100, where A and B represent the activities of the enzyme with and without sample, respectively. The assay was conducted with four replicates for each treatment.
Measurement of plasma triacylglycerol levels after oral administration of lipid emulsion to ICR mice
Lipid emulsion was prepared with 3.0 mL of corn oil, 40.0 mg of cholic acid, 100.0 mg of cholestryl oleate, and 3.0 mL of saline in the absence or presence of GSO or OEG (final concentration, 1 g/kg of body weight). Male mice fasted overnight were orally administered 200 μL of lipid emulsion. Blood samples were taken from the tail vein at 0, 1, 2, 3, and 4 hours after administration of the lipid emulsion with or without GSO or OEG using a heparinized capillary tube and centrifuged at 5,500 g for 5 minutes in a Model Micro 17R (Hanil Science Industrial Co. Ltd., Inchon, Korea) to obtain the plasma. The plasma TG was determined using a TG test kit (Asan Pharmaceutical, Seoul, Korea).
Antiobesity effects of OEG in mice
The protocol for this study was approved by the Animal Care and Use Committee of Kyungpook National University (Daegu, Korea). Male C57BL/6J mice, 6 weeks old (weighing 15–18 g), were purchased from Nara Biotech (Seoul) and were housed five mice per cage under a 12/12-hour light/dark cycle in a temperature-controlled room (22±2°C). Mice were allowed free access to feed and water. After 1 week of feeding chow diet, the mice were divided into the following four groups: CON, standard laboratory diet (control) group; HF, group fed AIN76A-based diet containing 30% shortening/corn oil (high fat) (20% shortening, 10% corn oil); GSO, group fed AIN76A-based diet containing 30% GSO (20% GSO, 10% corn oil); and OEG, group fed AIN76A-based diet containing 30% OEG (20% oil extract of ginseng, 10% corn oil). All the mice were fed pelletized chow for 14 weeks. The diets were stored at −20°C to avoid oxidation of lipids. The body weight of each mouse was recorded once a week, and the total amount of feed consumed was recorded twice a week.
Biochemical analyses of plasma
On the last day of the feeding period, the mice were fasted overnight. The next day, they were anesthetized with diethyl ether and sacrificed by cervical dislocation. Blood samples were collected from eyes into heparinized tubes, centrifuged at 3,000 g for 15 minutes at 4°C, and the plasma was stored at −20°C until analysis. Total cholesterol (TC), TG, high-density lipoprotein cholesterol (HDLC), and the activities of glutamate-oxaloacetate transaminase (GOT) and glutamate-pyruvate transaminase (GPT) in plasma were determined by a spectrometric method using a commercial diagnostic kit (Asan Pharmaceutical).
Examination of hepatic and adipose tissue morphology
The liver was dissected, rinsed with physiological saline, and weighed. The WATs (epididymal and subcutaneous) and liver were fixed in a buffer solution of 10% formalin. Fixed liver and adipose tissues were serially sliced at thicknesses of 2.0 and 5.0 μm using a microtome (RM 2125RT, Leica, Nussloch, Germany), respectively. The samples were stained with Harry's hematoxylin–eosin, and the stained area was measured by using light microscopy and an image analysis program.
Immunohistochemistry
Liver blocks were placed in formalin fixative and thereafter paraffin-embedded for archival use. Sections 2 μm thick were mounted on Super Frost Plus slides (Capillary Gap Plus microscope slides, Fisher Scientific, Waltham, MA, USA), baked at 65°C for 60 minutes, deparaffinized by xylene, rehydrated through graded alcohol rinses, and purified using 0.3% H2O2. Heat-induced antigen retrieval was performed by immersing the slides in 10 mM citrate buffer (pH 6.0) and microwaving in a microwave oven for 10 minutes. The slides were then cooled at room temperature for 30 minutes. After blocking with blocking reagent (Ultra Tek horseradish peroxidase, Immunotech, Vaudreuil-Dorion, Quebec, Canada, USA) for 10 minutes at room temperature, antigen was detected by a 1-hour incubation at room temperature with the relevant primary monoclonal rabbit anti-PPARγ antibody (diluted 1:200 in Tris-buffered saline buffer), followed by reacting with a secondary antibody conjugated to a peroxidase complex (Dako, Copenhagen, Denmark). Color development was done using diaminobenzidine plus chromogen. Whenever necessary, slides were counterstained with light hematoxylin. Standardization of the incubation and development times for each antibody allowed accurate comparisons in all cases. The slides were then dehydrated in a series of ethanol, in xylene, and cover-slipped.
Western blotting
Tissues were homogenized in precooled lysis buffer (20 mM Tris-Cl, 145 mM NaCl, 10% glycerol, 5 mM EDTA, 1% Triton-X, and 0.5% Nonidet P-40). After 30 minutes of incubation on ice, the homogenates were cleared by centrifugation at 14,000 g for 10 minutes at 4°C, and the supernatants were denatured in sample buffer for 5 minutes at 95°C. Proteins were separated by electrophoresis on a 10% sodium dodecyl sulfate–polyacrylamide gel for 1.5 hours at 100 V and transferred onto nitrocellulose membranes (Amersham Biosciences, Freiburg, Germany) for 1 hour at 100 V. Membranes were incubated with antibodies to PPARγ or β-tubulin at dilutions of 1:1,000 overnight at 4°C. The bands were detected using a chemiluminescence kit (Pierce, Cheshire, UK). Densitometry analysis was performed with Lab Image software (FUJI-LAS 4000mini, Fuji, Tokyo, Japan).
Statistical analysis
Data were expressed as mean±SD values. Whenever possible, data were subjected to analysis of variance, followed by Duncan's multiple range test, using SPSS software (SPSS Inc., Chicago, IL, USA). P<.05 was considered to be statistically significant.
Results
Profile of volatile components in OEG
Volatile compounds present in GSO and grape seed OEG are shown in Table 2. In total, 88 compounds were tentatively identified by gas chromatography/mass spectrometry. The compounds identified from ginseng oil and GSO could be grouped into terpenoids (12), alcohols (20), aldehydes (12), acids (30), and others (14). The levels of propyl acetate (21.6%), ethyl acetate (17.5%), acetic acid (8.6%), propionic acid (1.8%), n-propyl alcohol (18.9%), 2-butanol (12.7%), and ethyl alcohol (8.9%) were relatively high in ginseng oil, compared with GSO. In particular, most terpenoids were detected only in ginseng oil, indicating that the compounds are mainly derived from ginseng.
T, trace.
Pancreatic lipase inhibitory activity and mouse plasma TG levels after oral administration of lipid emulsion
We tested whether OEG could inhibit pancreatic lipase and might affect lipid digestion. A dose-dependent inhibition of pancreatic lipase activity was observed as shown in Figure 1. The enzyme activity was reduced by 20–77% in comparison with the CON group in the presence of OEG at the concentration of 0.1–100.0 mg/mL (Fig. 1A). GSO did not inhibit pancreatic lipase activity at concentrations below 100.0 mg/mL, although it suppressed the enzyme activity by 23.9% at 100.0 mg/mL.
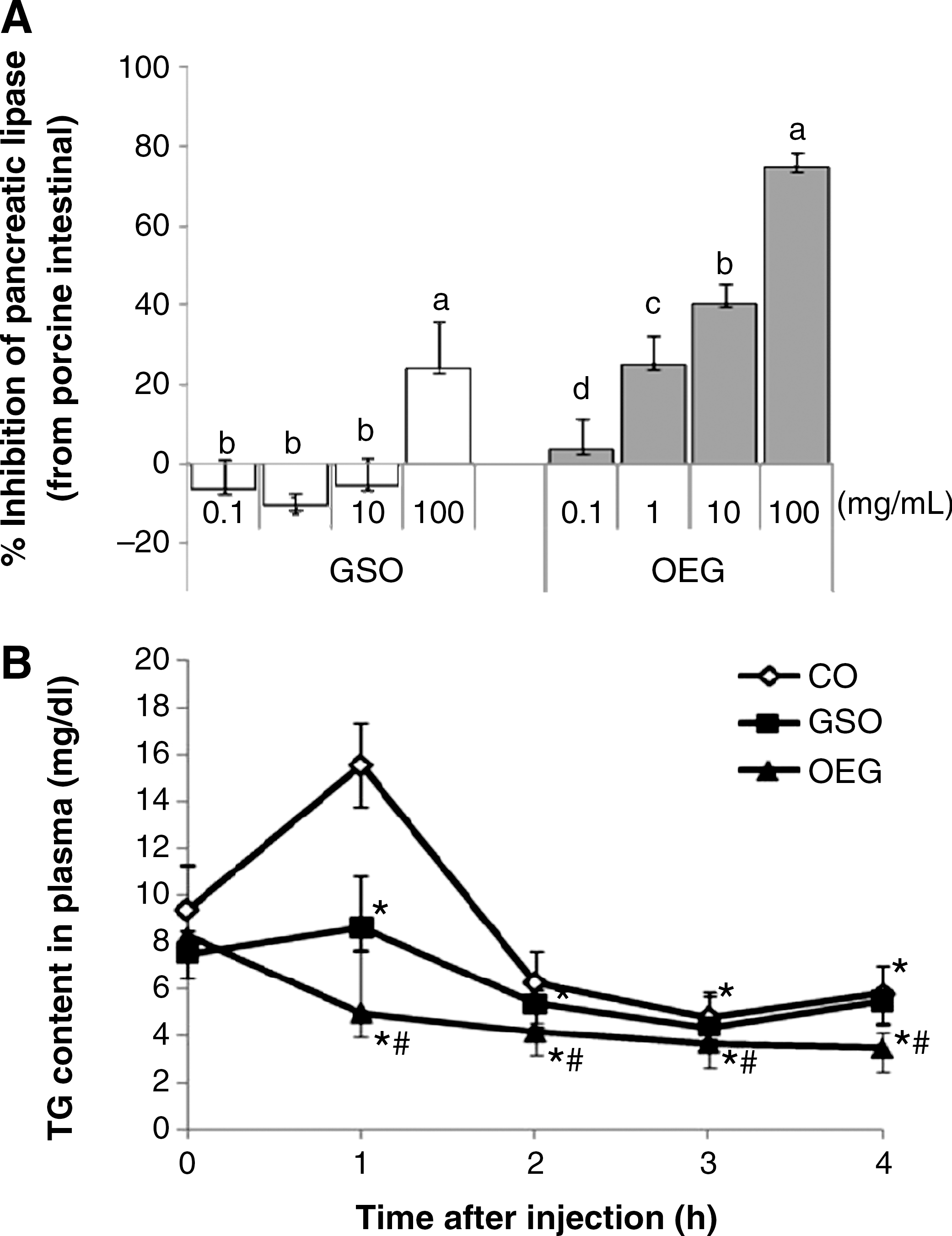
Oil extract of ginseng (OEG) inhibition of porcine pancreatic lipase activity in vitro and effect on plasma triglyceride (TG) levels after oral administration of a lipid emulsion. (
Because pancreatic lipase, a major lipid-digesting enzyme, was inhibited by OEG, we attempted to confirm the inhibition of TG absorption by OEG in mice. We studied the time course characteristics of the plasma TG level when lipid emulsion containing corn oil, GSO, or OEG was orally administered to mice (Fig. 1B). OEG prevented increases in plasma TG concentrations at 60 and 120 minutes after oral administration of lipid emulsion.
Body weight and food intake
Mice in all experimental groups were apparently healthy, showing no pathological signs or abnormalities during the entire experimental period. The changes in body weight and food intake during the experimental period are shown in Figure 2. All experimental groups had similar body weights at the beginning of the study (21.5±0.1 g). Although feeding HF diet containing 5% (wt/wt) corn oil and 10% (wt/wt) shortening caused significant weight gain (42.9±2.4 g) compared with animals fed the standard diet (31.8±2.6 g) throughout the study, the groups fed HF diet containing GSO or OEG gained less body weight, by 12.2% and 17.1%, respectively. There were no significant differences among the groups in food consumption (Fig. 2B). The feed gain ratio, calculated by dividing the weight increase by the food intake, for the mice given HF diet was 1.85 times higher than that for the mice fed the control diet (Fig. 2C). Taking the feed gain ratio of the control diet group as 100%, the mice fed high shortening (HF) diet showed a feed gain ratio of 185%, and those given GSO or OEG diet had feed gain ratios of 151.4% and 130.5%, respectively.
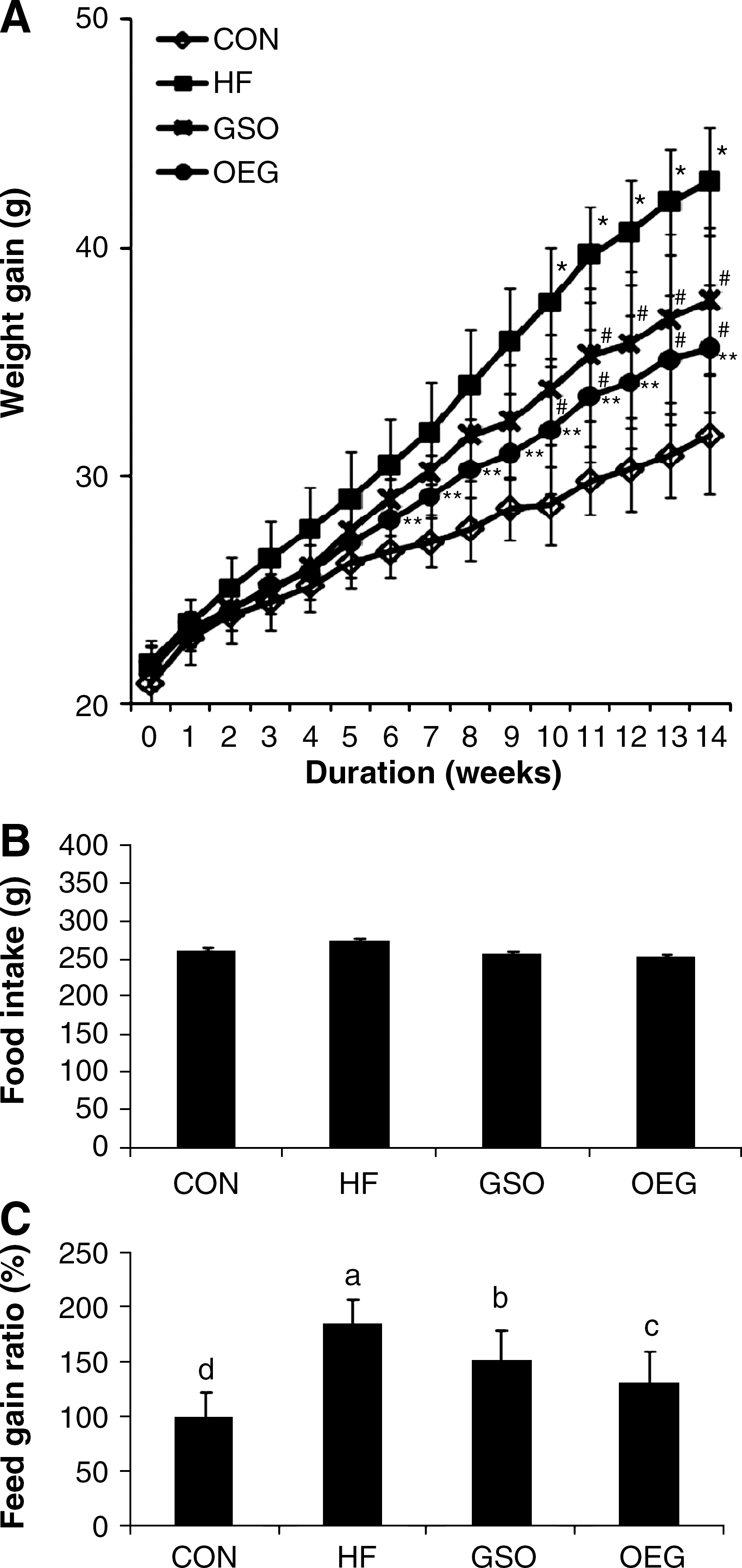
Effect of OEG-containing diet on body weight gain and feed intake of C57/BL6J mice. (
Effect of OEG-containing diet on fat accumulation in body
The changes in fat storage among the four experimental groups are shown in Table 3. The sum of the fat pad weights (epididymal WAT, perirenal WAT, interscapular browmn adipose tissue, subcutaneous WAT, and mesenteric fat) was significantly higher in the HF diet group than in the CON diet group, consistent with the body weight changes of experimental groups. The fat pad weights in the GSO and OEG groups were significantly less than in the HF group by 12.2% and 23.4%, respectively. Although there was no significant difference in epididymal fat mass between the OEG and HF diet groups, regional fat mass (perirenal WAT, interscapular brown adipose tissue, subcutaneous WAT, and mesenteric fat) was significantly lower in mice given OEG than HF diet (data not shown). Furthermore, body fat mass in mice fed OEG diet was significant lower than that of mice fed HF diet, although greater than the CON group. Mice fed OEG diet also tended to accumulate less body fat than the group fed GSO diet, but the difference was not statistically significant (Table 3).
Data are mean±SD values (n=9).
Values not sharing a common superscript letter in the same row differ significantly (P<.05).
AI, (total cholesterol – high-density lipoprotein [HDL] cholesterol)/HDL cholesterol); GOT, glutamate-oxaloacetate transaminase; GPT, glutamate-pyruvate transaminase; HTR, HDL cholesterol/total cholesterol.
Effect of OEG-containing diet on plasma lipids, GOT, and GPT
The levels of plasma lipids, GOT, and GPT are shown in Table 3. Plasma TG levels were highly correlated with final body weight and total fat pads. Plasma TG was highest in the HF diet group, but significantly lower in those fed the OEG diet than those fed HF with GSO, and tended to be lower, but not significantly, than in those fed CON diet. The plasma TC concentrations at week 14 in the CON, HF, GSO, and OEG diet groups were 86.4, 112.0, 95.0, and 93.7 mg/dL, respectively. Thus GSO and OEG diets significantly reduced the concentration of blood TC in C57BL/6J mice, compared with the HF diet (P<.05). HDLC levels in the mice fed OEG diet were not significantly different from those in mice fed CON and GSO diets.
Plasma GPT activity was significantly lower in the OEG diet group than in the HF diet group, but GOT activities were not different among experimental groups.
Histopathology of adipocytes and liver
The size of adipocytes in the HF diet group was significantly larger than those of the CON diet group (Fig. 3A), but tended to be smaller in mice fed OEG diet compared with animals fed HF or GSO diet.
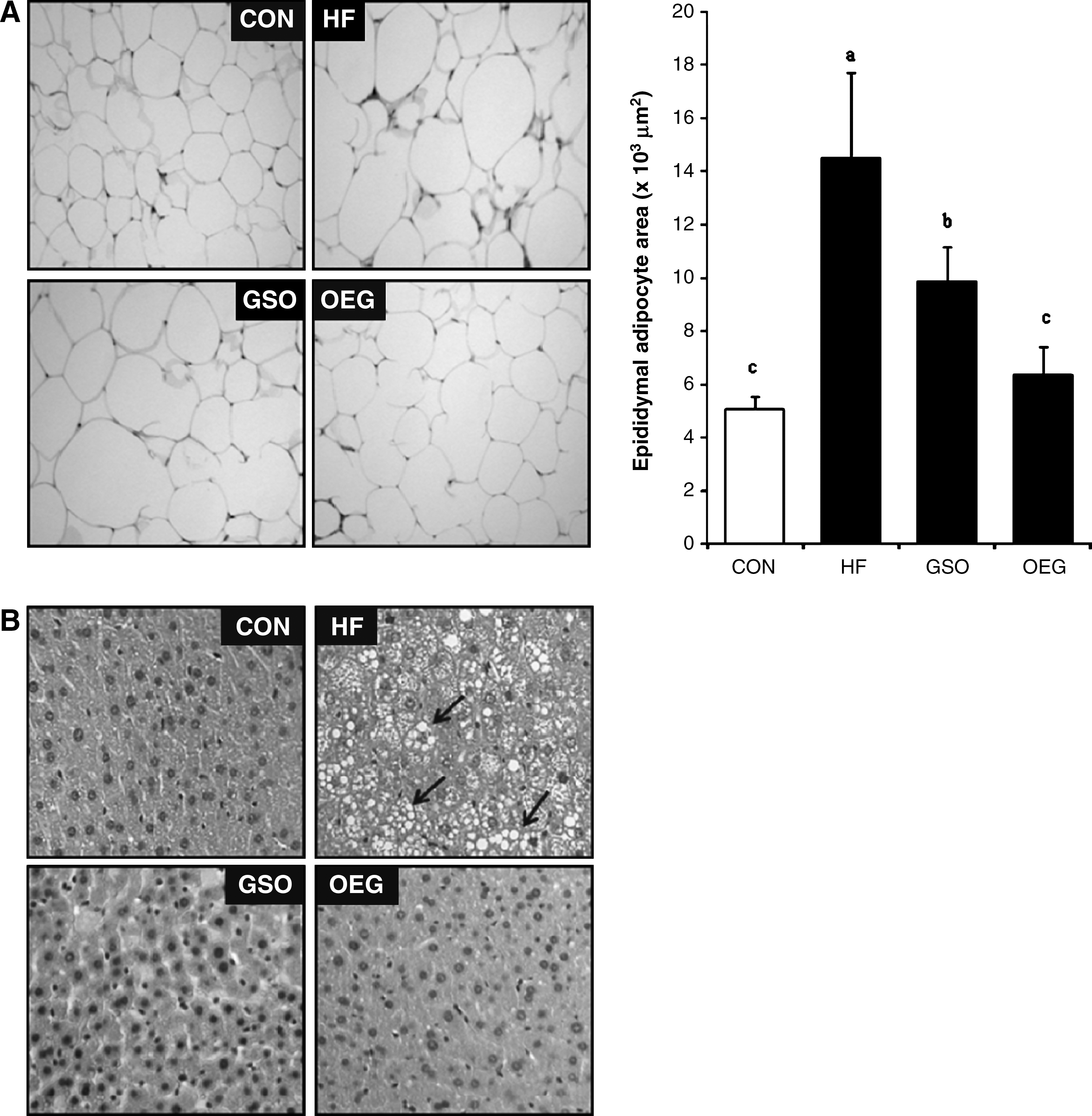
Effects of OEG on epididymal adipocytes and hepatic tissue morphology. There were nine mice per group. (
Histological examination results are shown in Figure 3B. Hepatocytes were arranged in an orderly fashion in the CON group, but cells from the HF diet group showed massive alterations such as swelling and accumulation of lipid droplets. This hepatic aberration in the HF diet group was not found in groups fed diets containing GSO or OEG. Rather, the hepatic morphology of the OEG diet group was similar to that of the CON group. These morphological results strongly suggest that the feeding with OEG inhibits the hepatic TG accumulation and adipose tissue expansion through affecting the process of hypertrophy rather than hyperplasia.
Immunohistochemical staining for PPARγ in liver tissue
Immunohistochemical analysis of liver tissue sections revealed that the expression of PPARγ, as brown granules in cytoplasm and nucleus, was apparently decreased in GSO and OEG diet groups, whereas it was obviously increased in response to 98 days feeding of the HF diet (Fig. 4A). In subcutaneous fat tissue, PPARγ expression was relatively low in the GSO and OEG diet groups, compared with the HF diet group. Furthermore, the expression of PPARγ in mice fed OEG diet was lower than that of animals fed GSO diet (Fig. 4B).
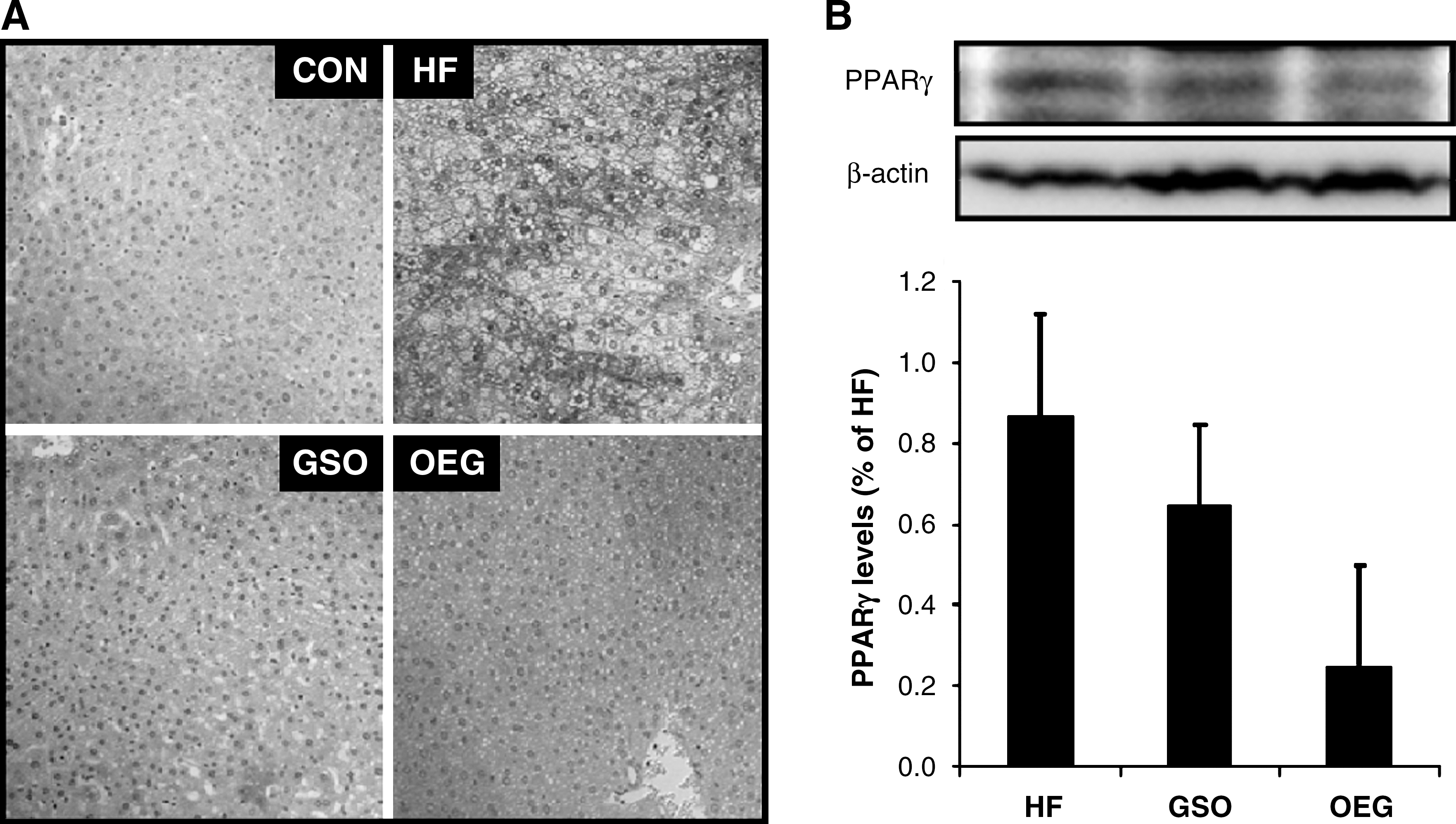
Effects of OEG on peroxisome proliferator-activated receptor γ (PPARγ) expression in (
Discussion
The most important finding of this study was that OEF significantly lowered weight gain in mice fed HF diet. These results are similar to the weight reduction effects of ginsenosides in animal models. 31,32 Previous studies have reported that HF diets increase body fat content without a higher energy intake. 33,34 The present study showed that the body fat mass in mice fed OEG-containing diet was significantly lower than that of mice fed the shortening diet, although it was greater than that of the CON group fed standard AIN76A diet. A known amount of food was given to the mice every 3 days, and the amount of food intake was monitored. When the CON diet group was assumed to have a 100% feed/gain ratio, the group given the shortening diet was found have a 185.0% feed/gain ratio, whereas the groups fed the GSO and OEG diet had values of 151.4% and 130.5%, respectively (Fig. 2C). These results imply that the relatively slow weight gain in the mice fed the OEG diet was not caused by suppression of appetite.
Previous studies reported that inhibiting the absorption of TGs could be an effective weight loss method. 35,36 In the present study, the mice given the OEG diet exhibited lower TG absorption than the control group, probably through inhibition of pancreatic lipase.
Most excess energy is stored in the form of TGs in adipose tissue, and increased adipose tissue mass arises through an increase in the hypertrophy and hyperplasia, which explains the interest in adipocyte proliferation and differentiation. 37 The morphology of the adipose tissues indicated that OEG inhibited the accumulation of TGs and adipose tissue expansion by modulating hypertrophy rather than hyperplasia.
Increased plasma TC or low-density lipoprotein cholesterol is a major risk factor for coronary heart disease. 38,39 Furthermore, obesity is a cause of dyslipidemia characterized by increased TG and decreased HDLC concentrations. 40 There are some reports that ginseng saponins decrease blood cholesterol levels in cholesterol-fed rabbits by increasing cholesterol excretion through stimulation of bile acid formation; 41,42 long-term administration of red ginseng products lowered cholesterol levels in obese women. 43 However, OEG did not contain appreciable amounts of ginseng saponins. Therefore, the antiobesity effect and improved lipid profiles after consumption of OEG-containing diet must be exerted by ginseng compounds other than saponins. For instance, lipophilic compounds released from ginseng during OEG preparation might be responsible for the effects described above.
High levels of acetate and its derivatives in ginseng oil were unexpected findings and may be a result of fermentation by facultative anaerobic microorganisms such as yeast and Propionibacterium contaminating the surface of raw ginseng. 44 There is also the possibility that altered lipid profiles of OEG might affect body weight gain and body fat accumulation.
In order to understand the action mechanism of OEG in lipid metabolism, the effect of OEG on the expression of PPARγ was measured. Recent studies showed that the antiobesity effect of ginseng is related to decreased PPARγ signaling in adipocyte as well as down-regulation of glutamate transporters. 15,45,46 As shown in Figure 4, OEG resulted in a significant suppression of PPARγ expression. Therefore, it was deducted that OEG might exert its antiobesity effect by inhibiting PPARγ expression as well as suppressing dietary fat digestion by lipase inhibition.
Among the PPAR isomers α, γ, and δ, PPARγ is the major transcription factor linked to adipocyte differentiation and is found almost exclusively in adipose tissue, indicating that it plays a crucial role both in the induction of adipose-specific genes and in the manifestation of the mature adipose phenotype. 47 –50 PPARγ is also expressed at low levels in liver. Animal studies have shown that activation of hepatic PPARγ promotes lipid accumulation, resulting in hepatic steatosis. 51 PPARγ2 also stimulates adipogenesis in fibroblasts, suggesting that it regulates the development of adipose tissue, unlike PPARγ1. 52 The C/EBP family of transcription factors, which consists of four members, plays a role in adipocyte differentiation and in the initiation of 3T3-L1 adipocyte differentiation. Once these transcription factors bind to DNA, they induce the synthesis of various adipogenic proteins relevant to adipocyte fat accumulation. 53 –55 Although the antiobesity effect of ginseng has been attributed to ginsenosides, a class of triterpene saponins, including compound K and Rg3, other terpenes, acetate, and its derivatives found in ginseng might exert antiobesity effects, possibly by modulating PPARγ. 14 In fact, acetic acid has been reported to suppress body fat accumulation by up-regulating fatty acid oxidation enzymes. 56 Thus, the possible interaction between the specific volatile component of ginseng and PPARγ signaling pathways needs be elucidated in further studies.
In conclusion, our study demonstrated that a diet supplemented with OEG led to less weight gain, lower plasma lipid levels, and smaller body fat cell size in C57BL/6J mice, compared with the groups fed HF diet alone or HF diet plus GSO. This study provides convincing evidence that OEG exerts antiobesity activity through the suppression of dietary TG absorption and/or regulation of PPARγ expression.
Footnotes
Acknowledgments
We thank the French Korean Aromatics Co. (Drs. Woo Sup Kwon and Min Ho Song) for analysis of volatile compounds. This study was supported by a grant (A050376) from the Korea Health Industry Development Institute funded by the Korean Government, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.