
Abstract
Some probiotics and their cell components are known to modulate lipid metabolism in vitro and/or in vivo. This study was carried out to investigate possible anti-adipogenic action of a probiotic cell extract, Lactobacillus plantarum KY1032 cell extract (KY1032-CE), in vitro using 3T3-L1 cells. Lipid regulation in the cell culture system was assessed by AdipoRed assay and Oil red O staining of intracellular lipids and real-time polymerase chain reaction and western blot analysis of adipogenesis-related factors. AdipoRed assay revealed that KY1032-CE treatment significantly decreased lipid accumulation in maturing 3T3-L1 preadipocytes in a dose-dependent manner. Oil red O staining demonstrated that KY1032-CE reduced the number of lipid-containing rounded cells. KY1032-CE down-regulated the mRNA and protein expression of four adipocyte-specific genes: peroxisome proliferator-activated receptor-γ2, CCAAT/enhancer binding protein-α, fatty acid synthase, and adipocyte-fatty acid binding protein. Accordingly, these results indicate that KY1032-CE can reduce fat mass by modulating adipogenesis in maturing preadipocytes. Further studies are needed to elucidate its mode of actions in efficacy tests of KY1032-CE in vivo.
Introduction
P
Although the relationship between probiotics and obesity has not been extensively studied, some probiotics were found to be effective in regulating adipose tissue in overweight adults 6 as well as in animal models of obesity. 7,8 Adipose tissue consists of adipocytes, which store triglycerides as fuel for energy reserves. However, excess adipose tissue leads to insulin resistance, thereby increasing the risk of type 2 diabetes and cardiovascular diseases. 9 In many obesity-related studies, the fibroblastic 3T3-L1 preadipocyte line is widely used to investigate the mechanism of preadipocyte proliferation and adipocyte genesis because of the ability of this cell line to undergo complete differentiation into mature adipocytes, which differentiate according to a program coordinated by adipogenic molecules, including various growth factors, cytokines, and hormones. 10,11
Adipogenic factors have long been recognized as important markers for body fat reduction in obesity. Excess adipose tissue mass is mainly caused by the differentiation of precursor cells into new adipocytes (adipogenesis), which involves expression of several transcription factors, including CCAAT/enhancer binding protein-α (C/EBPα), 12 peroxisome proliferator-activated receptor-γ (PPARγ), 13 and sterol regulatory element binding protein-1c. 14 C/EBPα and PPARγ regulate the expression of adipocyte genes such as adipocyte-fatty acid binding protein (A-FABP), 15 and sterol regulatory element binding protein-1c controls the expression of lipogenic genes such as fatty acid synthase (FAS). 16 The amount of adipose tissue mass can be reduced by inhibiting adipogenesis, 17 which may be regulated via these transcription factors.
L. plantarum KY1032 is one of the strains that are isolated from kimchi, a traditional fermented cabbage product in the Republic of Korea. L. plantarum species are lactic acid bacteria, which are among the most common types of microorganisms used as probiotics. Some strains of L. plantarum were reported to have the ability to produce antibacterial peptides, 18 modulate intestinal microbiota in dairy goats, 19 and induce cytokines. 20 In particular, administration of L. plantarum to mice led to reduced adipose tissue weights. 21,22 However, no study has yet investigated how L. plantarum cell extract functions when it is used to direct treat 3T3-L1 cells in a culture system. Accordingly, we investigated the anti-adipogenic effect of L. plantarum KY1032 cell extract (KY1032-CE) in 3T3-L1 preadipocytes. Its effects on cell viability and lipolysis were also evaluated in either maturing preadipocytes or mature adipocytes.
Materials and Methods
Isolation of L. plantarum KY1032 and preparation of the cell extract
L. plantarum KY1032 was isolated from kimchi. Its biological properties were characterized with various biochemical and physiological tests using API 50CHL (BioMérieux, Marcy l'Etoile, France), and identification was confirmed with DNA sequencing of 16S rRNA. KY1032-CE was prepared as described elsewhere. 23 In brief, after L. plantarum KY1032 was grown anaerobically in deMan–Rogosa–Sharpe medium at 37°C for 18 hours, the cells were collected by centrifugation and washed with phosphate-buffered saline (PBS). The cells were counted and suspended in PBS at 1010 colony-forming units/mL, followed by sonication and centrifugation. Supernatants containing the KY1032-CE were filter-sterilized (pore size, 0.45 μm), lyophilized, and kept at −20°C until use.
Cell line and cell culture
3T3-L1 mouse embryo fibroblasts were obtained from the American Type Culture Collection (Manassas, VA, USA). Cells were cultured as described elsewhere. 24 In brief, cells were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS) until confluent. Two days after confluence (day 0), the cells were stimulated to differentiate with DMEM containing 10% FBS, 1 μg/mL insulin, 0.5 mM 3-isobutyl-1-methylxanthine, and 1 μM dexamethasone for 2 days. On day 2, medium was replaced with DMEM containing 10% FBS and 1 μg/mL insulin and incubated for 2 days, followed by culturing with DMEM containing 10% FBS for an additional 4 days (day 8), at which time more than 90% of cells were mature adipocytes with accumulated fat droplets. Cells were maintained at 37°C in a humidified 5% CO2 atmosphere.
Quantification of lipid content
For the quantification of lipid content, the AdipoRed assay reagent (Lonza, Walkersville, MD, USA) was used. Tests were performed in 96-well plates. Two-day postconfluent preadipocytes were treated with differentiation medium containing either vehicle or KY1032-CE during the adipogenic phase. On day 6, treatment medium was removed, and plates were carefully rinsed with PBS. Wells were then filled with 200 μL of PBS, and 5 μl of AdipoRed assay reagent was added and incubated for 10 minutes at room temperature. Fluorescence was measured with excitation at 485 nm and emission at 572 nm.
Cell viability assay
For the assessment of cell viability, the CellTiter 96® AQueous nonradioactive cell proliferation assay kit (Promega, Madison, WI, USA) was used. Tests were performed in 96-well plates. 3T3-L1 preadipocytes were incubated with either vehicle or KY1032-CE for 6 days during the adipogenic phase. Thereafter, treatment medium was removed and replaced with 100 μL of fresh DMEM containing 10% FBS plus 20 μL of 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt solution. Cells were incubated in the dark for 1 hour at 37°C, and the absorbance was measured at 490 nm in a Synergy™ HT plate reader (Bio-Tek Instruments, Winooski, VT, USA) to determine the formazan concentration, which is proportional to the number of live cells.
Oil red O staining
Cells were treated with either vehicle or KY1032-CE for 6 days during adipogenesis as described above. Thereafter cells were washed with PBS, fixed with 10% formalin, and stained with Oil red O and hematoxylin as described by Suryawan and Hu. 25 After cells were mounted with glycerol gelatin, images for each dish were captured using a microscope (AxioObser Z1, Carl Zeiss, Oberkochen, Germany).
Lipolysis assay
Fully differentiated adipocytes (day 8) were treated for 48 hours with either vehicle or KY1032-CE in 96-well plates. At the end of incubation, the free glycerol released was assayed with a glycerol reagent A (Zen-Bio, Research Triangle Park, NC, USA) following the manufacturer's instructions. In brief, 100 μL of glycerol reagent A was added and incubated at room temperature for 15 minutes, and absorbance was measured at 540 nm in a Synergy HT plate reader to determine the glycerol content of the sample.
RNA extraction
Postconfluent preadipocytes were incubated with either vehicle or KY1032-CE for 6 days during adipogenesis. After treatment, RNA samples were extracted using an RNAqueous® kit (Ambion, Austin, TX, USA), and the purity of RNA was increased by removing unnecessary DNA with a Turbo DNA-free™ kit (Ambion) according to the manufacturer's protocols. The integrity of the RNA extracted from all samples was verified and quantified using a GeneQuant Pro spectrophotometer (GE Healthcare, Piscataway, NJ, USA).
Reverse transcription and real-time polymerase chain reaction
Two micrograms of total RNA in a 20-μL reaction mixture was reverse-transcribed into cDNA using the high-capacity RNA-to-cDNA kit (Applied Biosystems Inc., Foster City, CA, USA) following the manufacturer's instructions. The cDNA was amplified on a 7500 Real Time PCR System (Applied Biosystems) using mouse-specific Taqman probe sets for PPARγ2 (catalog number Mm00440945_m1), C/EBPα (catalog number Mm00514283_s1), FAS (catalog number Mm00662319_m1), and A-FABP (catalog number Mm00445880_m1) according to the manufacturer's protocol under the following conditions: 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds, and 60°C for 1 minute. All samples were assayed in triplicate, and mRNA expressions were normalized by using glyceraldehyde 3-phosphate dehydrogenase (catalog number Mm99999915_g1) as an endogenous control to correct the differences in the amount of total RNA added to each reaction. All polymerase chain reactions for each sample were performed simultaneously on the same plate.
Western blot analysis
Cells were treated with either vehicle or KY1032-CE for 6 days during the adipogenic phase as described above. To prepare the whole-cell extract, cells were washed with PBS and suspended in a lysis buffer (Cell Signaling, Beverly, MA, USA) including 1 mM phenylmethylsulfonyl fluoride. After 30 minutes of rocking at 4°C, the mixtures were sonicated and centrifuged. The supernatants were collected as the whole-cell extracts, separated by NuPAGE® 4–12% Bis-Tris gel (Invitrogen, Carlsbad, CA, USA), and transferred to nitrocellulose membranes (pore size, 0.45 μm) (Invitrogen). The membranes were blocked with 5% nonfat dry milk in Tris-buffered saline with Tween-20 buffer (10 mM Tris-HCl [pH 7.5], 150 mM NaCl, and 0.05% Tween-20) and incubated overnight with various antibodies against PPARγ2, C/EBPα, FAS (Cell Signaling), A-FABP, and glyceraldehyde 3-phosphate dehydrogenase (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4°C. Chemiluminescent signals were developed using an enhanced chemiluminescence kit (GE Healthcare, Waukesha, WI, USA).
Statistical analysis
Results were presented as mean values with their SEs. Statistical analysis was performed using SPSS software (SPSS Inc., Chicago, IL, USA). Data were analyzed by one-way analysis of variance, and the differences between experimental groups were evaluated using Duncan's multiple range test at the P<.05 level.
Results and Discussion
KY1032-CE was obtained by lysing L. plantarum KY1032 suspended in PBS at 1010 colony-forming units/mL and then used to treat 3T3-L1 preadipocytes during the adipogenic phase. Six days following treatment with KY1032-CE at 0.01% and 0.1%, lipid accumulation was decreased, according to the AdiopoRed assay, by 11±3.6% (P<.01) and 30.6±9.2% (P<.001), respectively (Fig. 1A). However, KY1032-CE treatment did not alter cell viability significantly (data not shown), indicating that the inhibitory effect of KY1032-CE on lipid accumulation is due not to reduced cell viability, but mainly to decreased adipogenesis. As shown in Figure 1B, KY1032-CE caused a greater decrease in lipid-containing rounded cells compared with control cells when visualized by staining. In addition to these anti-adipogenic effects, we investigated whether KY1032-CE exerts a lipolytic effect in mature 3T3-L1 adipocytes. KY1032-CE slightly increased lipolysis, by approximately 7%, when it was added to fully differentiated 3T3-L1 adipocytes (data not shown).
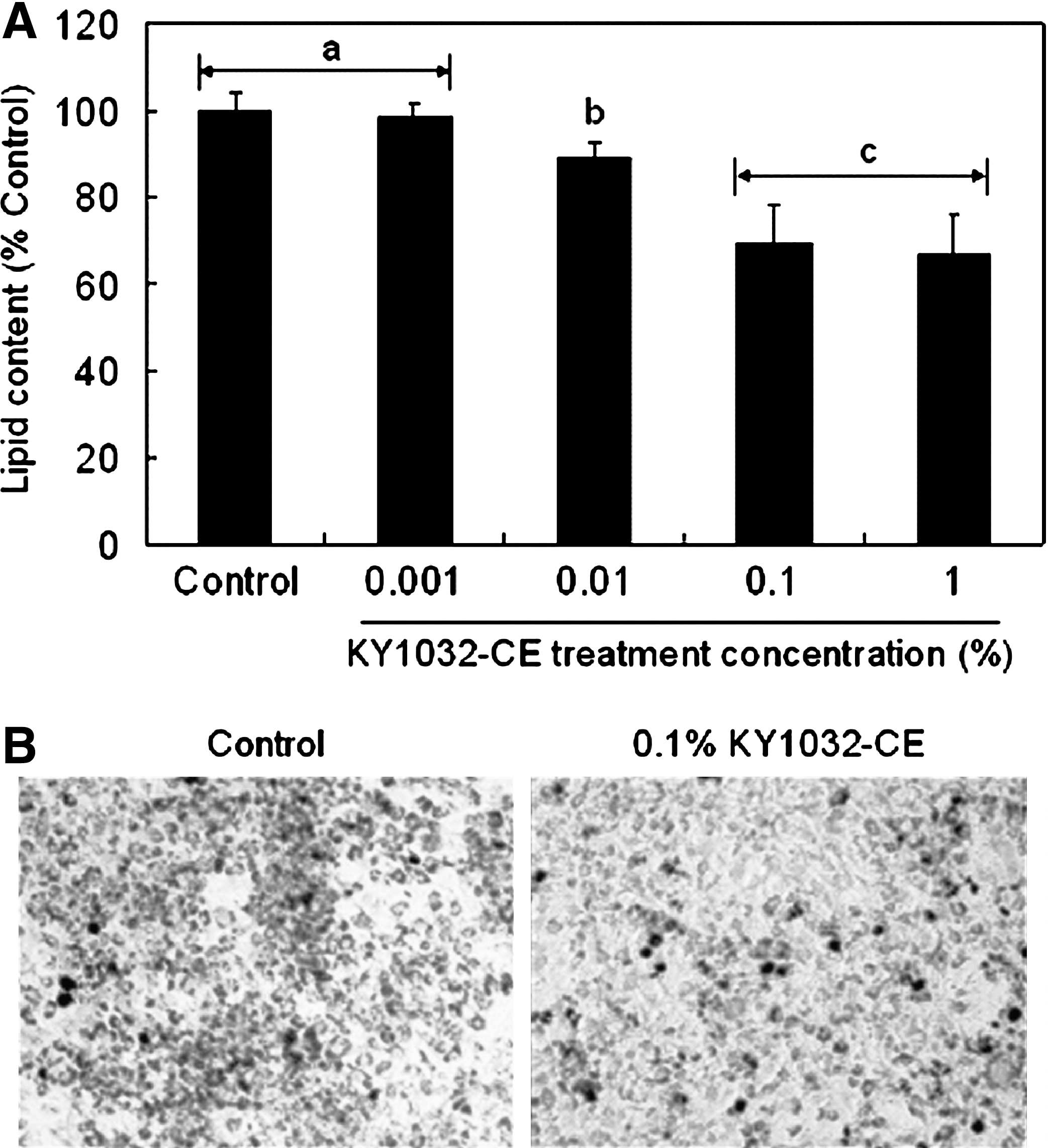
Effect of L. plantarum KY1032 cell extract (KY1032-CE) on lipid content of maturing preadipocytes. (
To examine whether the anti-adipogenic effect of KY1032-CE was related to molecular changes, real-time polymerase chain reaction and western blot analysis were used to measure the expression levels of four particular adipocyte-specific genes: PPARγ2, C/EBPα, FAS, and A-FABP. It is interesting that KY1032-CE treatment led to a decrease in PPARγ2, C/EBPα, FAS, and A-FABP mRNA expression by 28.3±5.1%, 23±8.2%, 30.8±7.2%, and 36.3±6.1% (P<.001), respectively (Fig. 2A). Consistent with mRNA expression data, the profiles of their protein expression levels were also significantly down-regulated by KY1032-CE treatment (Fig. 2B). The decreased lipid accumulation in 3T3-L1 cell by KY1032-CE treatment was partly due to the down-regulation of these adipocyte-specific genes.

Effect of KY1032-CE on the expression of adipocyte-specific genes. (
In the present study, we demonstrated how KY1032-CE can exert an anti-adipogenic effect in 3T3-L1 cells. KY1032-CE was used to avoid a problem for coculture of mammalian cells with L. plantarum KY1032 cells, lowering the pH of medium. Changes in cell morphology indicate that KY1032-CE significantly decreased lipid accumulation in maturing 3T3-L1 preadipocytes in a dose-dependent manner with simultaneous decreases in the mRNA and protein expression levels of these adipocyte-specific genes in maturing preadipocytes. PPARγ2 and C/EBPα, as transcriptional activators, are major coordinators of adipocyte gene expression during adipocyte differentiation. 26 FAS and A-FABP, as downstream adipocyte genes, contribute to synthesis of fatty acid and regulation of diverse lipid signals, respectively. According to the results, the down-regulation of these genes by KY1032-CE can prevent maturation of 3T3-L1 preadipocytes. This clearly indicates that KY1032-CE led to decreases in both mRNA and protein expression of PPARγ2, C/EBPα, FAS, and A-FABP, thereby suppressing the differentiation of preadipocytes to adipocytes.
The loss of fat mass can also be partly attributed to lipolysis, in which triglycerides are broken down into glycerol and fatty acids in adipocytes. Several natural compounds such as genistein and quercetin are already known to exert anti-obesity effects by stimulating lipolysis in adipocytes as well as inhibiting adipogenesis in preadipocytes. 27 However, KY1032-CE treatment of mature adipocytes did not induce a significant increase in lipolysis, indicating that KY1032-CE may not likely break down preexisting fats in adipocytes.
The generally accepted definition of probiotics is live and nonpathogenic microorganisms ingested by the target species, which may be human or animal. However, there is considerable published evidence that probiotic preparations comprising dead cells or their metabolites can also exert a biological response, in many cases similar to that seen with live cells. 28,29 Consequently, probiotics consisting of either live or dead cells or their cell extracts containing metabolites may play important roles in modulating biological response or health maintenance in the host. There will always be the possibility that some dead cells or cell metabolites are being ingested with the live cells when one consumes the probiotic products. Because various biological responses have been shown by dead, frequently heat-killed probiotics, despite the general assertion that probiotics are live microorganisms, preparations of probiotics seem to be another avenue of interest for developing the functional foods.
Among the various beneficial health effects of probiotics, 30,31 their biological impact on obesity has generated considerable interest. Regarding the anti-obesity property of probiotics, some ironic results have been reported, suggesting increased risk of obesity when children and farm animals were supplemented with probiotics for the purpose of diarrhea treatment and growth enhancement, respectively. 32 However, these data were not sufficient to confirm that probiotics can increase the risk of obesity because in the former case the period of time for treating children with probiotics was too short to evaluate the propensity of probiotics to cause obesity, and in the case of the latter, probiotics used in livestock industry just promoted growth and lean mass, not adiposity. 33,34 It is plausible that manipulation of the intestinal microbiota or modulation of metabolic response by probiotic components could be a novel approach for prevention of obesity, although its mode of action needs to be elucidated in an in vivo study. Some reports demonstrated that dairy products fermented with lactic acid bacteria exert anti-obesity effects. 35
In conclusion, this present work demonstrated for the first time that the cell extract of a new probiotic strain, L. plantarum KY1032, inhibited adipogenesis in 3T3-L1 cells. Furthermore, this effect was, at least partly, mediated through suppressing PPARγ2, C/EBPα, FAS, and A-FABP expression. More studies are required for further scientific validation of the anti-adipogenic or anti-obesity effects of L. plantarum KY1032 and its extract in vivo.
Footnotes
Acknowledgments
We thank K.-S. Myoung (Korea Yakult Co., Ltd., Yongin, Republic of Korea) for critical discussion. This research was supported by Korea Yakult and the SRC Program (Center for Food & Nutritional Genomics: grant number 2010-0001886) of the National Research Foundation (NRF) of Korea funded by the Ministry of Education, Science and Technology.
Author Disclosure Statement
D.-Y.P., Y.-T.A., and C.-S.H. are employees of Korea Yakult Co., Ltd. S.-M.J. and M.-S.C. declare no competing financial interests exist.