
Abstract
The aim of this study was to examine the effect of Shengmai-san (SMS) on learning and memory impairment induced by scopolamine (1 mg/kg, i.p.) in mice. The passive avoidance task (PAT) and Morris water maze (MWM) test served as the behavioral models for testing memory. To elucidate the mechanism of its cognitive-enhancing activity, the effects of SMS (2, 4, or 8 g/kg) on activities of acetylcholinesterase (AChE) and antioxidant enzymes and levels of acetylcholine (ACh) and nitrite were evaluated in brain homogenate. Tacrine (THA) (10 mg/kg, p.o.) was used as a reference drug. SMS (4 or 8 g/kg) significantly prevented scopolamine-induced impairments as measured by the PAT and MWM (probe trial session). SMS (4 or 8 g/kg) also significantly reduced the oxidative-nitrative stress, as evidenced by decreased malondialdehyde and nitrite levels and by its prevention of decreases in glutathione and superoxide dismutase levels. The activity of AChE was decreased in scopolamine-treated mice but was inhibited significantly by SMS treatment (4 or 8 g/kg) in both salt- and detergent-soluble fractions of brain homogenates. Further SMS treatment (4 or 8 g/kg) significantly increased the ACh levels in the brain homogenate to a level similar to that observed in THA treatment. Thus, the significant cognitive enhancement observed after 7 days of administration of SMS is closely related to the strong antioxidant properties of SMS in addition to its inhibition of brain AChE activity. These findings stress the critical impact of SMS on higher brain functions such as learning and memory.
Introduction
A
Recently, El-Sherbiny et al. 7 reported that memory impairments in a scopolamine-induced animal model are associated with altered oxidative stress in the brain. Strong evidence supports the involvement of oxidative stress in degenerative changes in the forebrain cholinergic system. 8 The findings led to the notion that drugs with antioxidative effects might be beneficial for preserving brain function. Antioxidative enzymes display reduced activities in affected brain regions of AD patients. 9,10 Moreover, the decreased levels of intracellular oxidized protein under these conditions are associated with the improvement of cognitive and/or psychomotor functions. 11 Thus, immense efforts have been directed to find therapeutic agents that could reduce the oxidative damage and promote a functional recovery in degenerative disorders. Chemicals with specific targets are clinically used to treat dementia in AD, such as acetylcholinesterase (AChE) inhibitors; however, AD involves complex reactions. Hence, antioxidant-based complex formulas might be a more reasonable approach for the treatment of AD. Traditional herbal medicines are the target of the study as they are composed of several herbs with different functional properties and are basically antioxidative. 12
In traditional Chinese medicine, Shengmai-san (SMS), which is composed of Panax ginseng, Ophiopogon japonicus, and Schisandra chinensis, has been used for treating symptoms related to cardiovascular diseases such as heart failure, stroke, and shock. Recent studies have begun to provide evidence showing that SMS has protective effects against oxidative damage in the cells or tissues of the cardiovascular system and nervous system. 13 –15 Recently, Wu et al. 16 have reported the memory-enhancing potential of SMS in the vascular dementia model. Moreover, we have shown the preventive effect of SMS against cerebral oxidative damage in rats, 17 and in an in vitro model, SMS has been reported to protect against amyloid-β-induced cytotoxicity in PC12 cells. 18 However, no attempts have been made to investigate whether SMS exerts cholinergic effects in an amnesic model. Therefore, the present study was designed to investigate the effect of SMS in scopolamine-induced cognitive dysfunction and oxidative and nitrative stress, and also to establish cholinergic modulatory efficacy.
Materials and Methods
Animals
The experiments were carried out on male BALB/c mice (12–14 weeks old, weighing 25–30 g). The mice were housed in stainless steel cages and kept under controlled conditions at 22±3°C, 55±5% relative humidity, and a 12-hour light/dark cycle throughout the experiment. Food in the form of commercially available pellets and water were available ad libitum. The animal experiments were performed according to internationally accepted ethical standards and with approval from Niigata University of Pharmacy and Applied Life Sciences (Niigata, Japan).
Sample preparation and treatment
The SMS preparation, manufactured in the pharmaceutical factory of West China Medical University and commercially available as Bakumisan karyu, was kindly provided by Isukura Pharmaceutical Co., Ltd. (Tokyo, Japan). The three component herbs of the SMS preparation (6 g) are P. ginseng (3 g), O. japonicus (2 g), and S. chinensis (1 g). For each experiment, the SMS granules were weighed precisely and solubilized in distilled water administered orally for 7 days at different (2, 4, or 8 g/kg) doses. To evaluate the pharmacological efficacy of SMS, tacrine (THA) was used at the same time as a reference (positive control). THA (10 mg/kg, p.o., for 7 days) and scopolamine (1 mg/kg, i.p.) were dissolved in saline before treatment.
Drugs and chemicals
THA, physostigmine, scopolamine, acetylthiocholine iodide, and 5,5′-dithiobis(2-nitrobenzoic acid) were purchased from Sigma-Aldrich (St. Louis, MO, USA). All other chemicals used in the study were of analytical grade. Solutions of the drug and chemicals were freshly prepared before use.
High-performance liquid chromatography analysis
To evaluate the quality of the SMS preparation, schisandrin and gomisin A were determined by high-performance liquid chromatography analysis. The stock solutions of schisandrin and gomisin A were diluted to an appropriate concentration range (0–50 μg/mL) for standard curves. The SMS solution was centrifuged (Himac CT 13, Hitachi, Japan) at 10,000 rpm for 10 minutes, and both were analyzed by high-performance liquid chromatography using a Shimadzu (Kyoto, Japan) system equipped with a photodiode array detector (model SPD-M10Avp). The following conditions were used: Mightysil RP-18 column (Kanto Chemical Co., Tokyo, Japan) (250×4.6 mm i.d., 5 μm particle size), linear gradient elution with acetonitrile and water solution, and flow rate of 0.8 mL/minute. Aliquots of SMS and standard solutions (20 μL) were injected to the column. Temperatures of the column oven and autosampler tray were set at 30°C and 15°C, respectively. Ultraviolet detection was set at 254 nm.
Passive avoidance task
Training for and testing of passive avoidance performance were carried out in two identical light and dark square boxes. 19 On day 7, the mice were initially placed in the light chamber, and 10 seconds later the door between the compartments was opened. When mice entered the dark compartment, the door was closed, and an electrical foot shock (0.1 mA/10 g of body weight) for a period of 2 seconds was delivered through stainless steel rods (one training trial). Six mice were used per treatment. Mice received SMS or THA 1 hour before the training trial. Amnesia was induced using scopolamine (1 mg/kg) 30 minutes after SMS or THA administration. Twenty-four hours after the training trial, the mice were again placed in the light compartment. The escape latency to enter the dark compartment was measured. If the mice did not enter the dark compartment within 300 seconds, the experiment was stopped.
Morris water maze test
A spatial memory test was performed using the Morris water maze (MWM) test. The MWM is a circular pool (50 cm in diameter and 40 cm in height) with a featureless inner surface. The circular pool was made opaque by the addition of milk powder. The pool was divided into four quadrants of equal area. A white platform (5 cm diameter and 25 cm height) was centered in one of the four quadrants of the pool and submerged 1 cm below the water surface so that it was invisible at water level. The day prior to the experiment was dedicated to swim training for 60 seconds in the absence of the platform. In the following days, the mice were given single trial sessions each day for 4 consecutive days. During each trial, the time taken to swim to the platform (escape latency) was recorded. Once the mouse located the platform, it was permitted to remain on it for 10 seconds and then removed from the pool. 20 One day after the last trial sessions, mice were subjected to a probe trial session in which the platform was removed from the pool; mice were allowed to swim for 120 seconds to search for it. A record was kept of the swimming time in the pool quadrant where the platform had been previously placed. Six mice were used per treatment. Mice were treated with SMS or THA 1 hour before the training trial. Amnesia was induced using scopolamine 30 minutes after SMS or THA administration. All mice were tested for spatial memory 30 minutes after the administration of scopolamine.
Tissue preparation
The mice were sacrificed by cervical dislocation following the behavioral study. The whole intact brain was carefully removed and placed in an ice-chilled petri dish for cleaning. The cerebellum was rapidly removed, and the remaining brain was weighed, washed with isotonic saline, and homogenized (10% wt/vol) in sodium phosphate buffer (30 mmol/L, pH 7.0).
One-half of the volume of this homogenate was separated and used as a salt-soluble (SS) fraction for assay of AChE and antioxidant activity. Triton X-100 (1% [wt/vol], 30 mmol, sodium phosphate buffer, pH 7.0) was then added slowly while the homogenate was stirred on ice, in sufficient volume to make the final volume contain 10% homogenate. All homogenates were centrifuged at 100,000 g at 4°C for 60 minutes. This fraction was used as a detergent-soluble (DS) fraction. 21 The supernatant was collected and stored at 4°C. The protein concentrations were determined by the Bradford assay with bovine serum albumin as a standard (0.05–1.00 mg/mL).
AChE assay
AChE activity was determined using the colorimetric assay of Ellman, as previously described. 22 In brief, in 96-well plates, 25 μL of 15 mmol of acetylthiocholine iodide, 75 μL of 3 mmol of 5,5′-dithiobis(2-nitrobenzoic acid), and 75 μL of 50 mmol of Tris-HCl (pH 8.0) containing 0.1% bovine serum albumin were added, and the absorbance was measured at 405 nm after incubation for 5 minutes at room temperature. Any increase in absorbance due to the spontaneous hydrolysis of the substrate was corrected for by subtracting the rate of the reaction before adding the enzyme. Then, 25 μL of each sample (SS and DS fractions) was added, and the absorbance was measured again after 5 minutes of incubation at room temperature. All determinations were carried out twice per sample with three equivalent samples.
Estimation of nitrite
The accumulation of nitrite in the supernatant, an indicator of the production of nitric oxide, was determined by colorimetric assay using Greiss reagent as described by Green et al. 23 Equal volumes of supernatant and Greiss reagent were mixed, the mixture was incubated for 10 minutes at room temperature in the dark, and absorbance was determined at 540 nm spectrophotometrically. The concentration of nitrite in the supernatant was determined from the sodium nitrite standard curve and expressed as μmol/mg of protein.
Assay of ACh
ACh was determined by using the method of Hestrin as described by Vincent et al. 24 In brief, 10% brain homogenate in cold saline was prepared (10,000 rpm [Physcotron™ NS-50, Microtec, Chiba, Japan], twice for 10 seconds with a 30 seconds interval) in ice. The aliquots (0.8 mL) of brain homogenate were mixed with 1.4 mL of distilled water, 0.2 mL of 1.5 mmol of physostigmine, and 0.8 mL of 1.84 mol of trichloroacetic acid and blended adequately. After centrifugation, 1 mL of each supernatant was added to 1 mL of basic hydroxylamine. The mixture was incubated for 15 minutes at 25°C, and then 4 mol of HCl (0.5 mL) and 0.37 mol of FeCl3 (0.5 mL) were added. Absorbance was read at 540 nm and calibrated with the standard (0.2 μmol/mL ACh).
Antioxidant assay
The SS fraction of brain homogenate was used for several different antioxidant enzyme assays. The quantitative measurement of malondialdehyde (MDA), the end product of lipid peroxidation, in brain homogenate was performed according to the method of Ohkawa et al. 25 Reduced glutathione (GSH) was determined by the method of Ellman, 26 which is based on the development of a yellow color due to the reaction of 5,5′-dithiobis(2-nitrobenzoic acid) with compounds containing sulfhydryl groups. The glutathione peroxidase (GPx) activity was determined according to the method described by Lankin et al. 27 using tert-butyl hydroperoxide as a substrate. The superoxide dismutase (SOD) activity was determined using the water-soluble tetrazolium method. 28
Statistical analysis
The results are given as mean±SEM values. The data obtained were analyzed by one-way analysis of variance followed by Tukey's post hoc test. Differences were considered significant at the 5% level.
Results
The high-performance liquid chromatogram of SMS solution showed peaks representing schisandrin and gomisin A at 31.6 and 35.1 minutes, respectively, which coincided with those of the standard solutions. The contents of the reference ingredients schisandrin and gomisin A were 0.124 μg/g and 0.567 μg/g, respectively. The peak profile featuring in schisandrin and gomisin A was similar to that observed in our previous study. 18
Effects of SMS on memory impairment induced by scopolamine in the passive avoidance task
During acquisition trials, latency times were not different among the experimental groups of mice following acute treatment with SMS. For retention trials, the analysis of variance followed by Tukey's test revealed significant differences for latency time in the acute SMS treatment. The step-through latency of scopolamine-treated mice was significantly (F 5,30=11.07, P<.001) shorter than that of the control group. In scopolamine-treated mice treated with THA (positive control group), step-through latency was significantly (P<.001) higher than that of the scopolamine-treated group (Fig. 1). Moreover, the shorter step-through latencies induced by scopolamine were significantly attenuated by acute SMS treatment at 4 g/kg (P<.05) and 8 g/kg (P<.01).
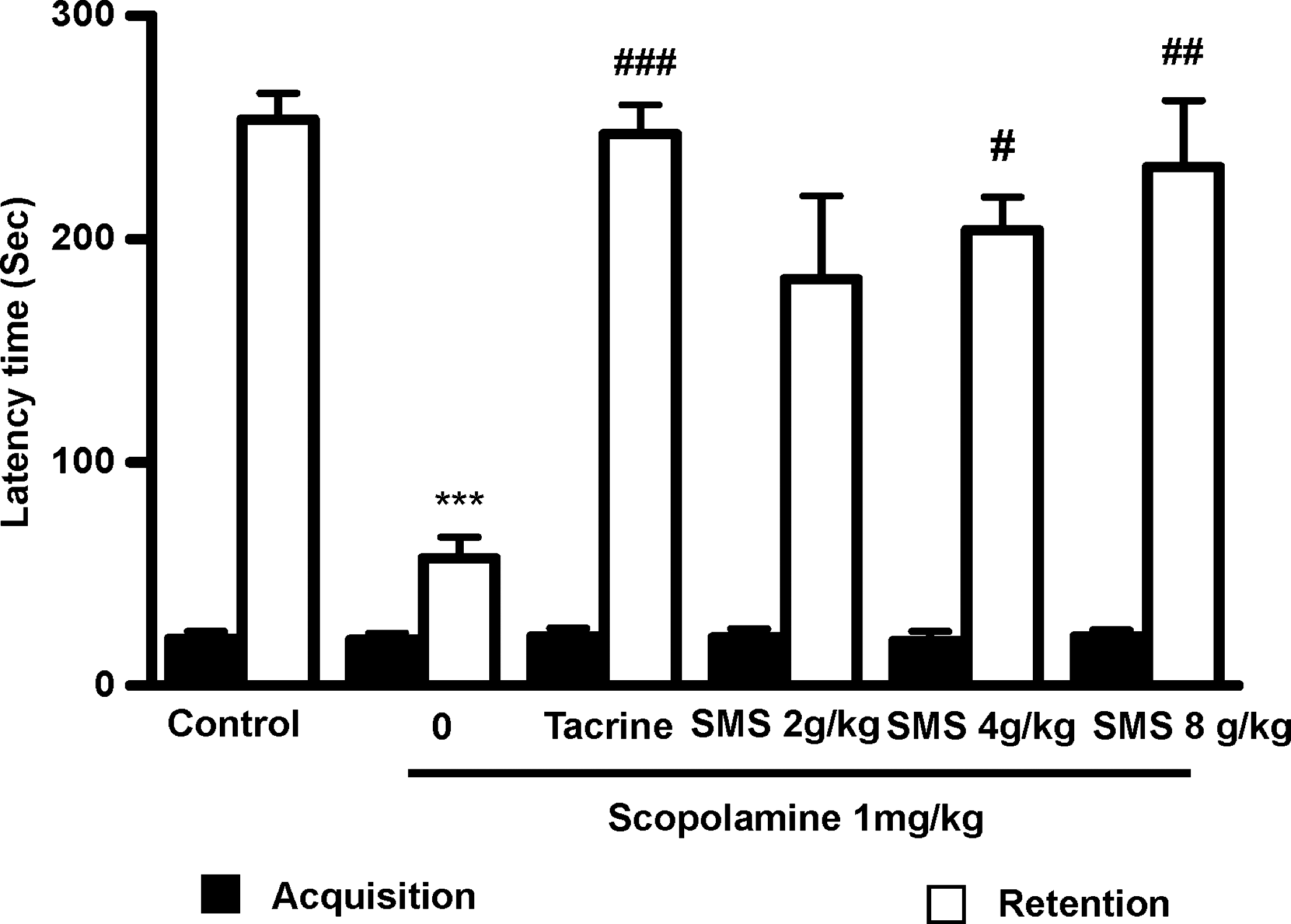
Effects of Shengmai-san (SMS) on scopolamine-induced memory impairment in the passive avoidance task response in mice. To study the effect of SMS on the scopolamine-induced memory deficit model, mice were administered SMS (2, 4, or 8 g/kg) or tacrine (10 mg/kg, p.o., as the positive control) 1 hour before the acquisition trial. Memory impairment was induced by scopolamine treatment (1 mg/kg, i.p.) 30 minutes after SMS or tacrine administration, and the acquisition trials were carried out 30 minutes after scopolamine treatment. At 24 hours after the acquisition trials, retention trials were carried out. Data are mean±SEM values (n=6). ***P<.001, statistically different from the control group. # P<.05, ## P<.01, ### P<.001, statistically different from the scopolamine-treated group.
Effects of SMS on memory impairment induced by scopolamine in the MWM test
The effect of SMS on spatial learning was evaluated using the MWM task. As shown in Figure 2A, the scopolamine-treated group exhibited longer escape latencies (day 1, 109±18.5 seconds; day 4, 74.7±12.1 seconds) throughout the training compared with the control group (day 1, 83.1±17.7 seconds; day 4, 23.5±8.4 seconds). The escape latencies with SMS (4 or 8 g/kg) significantly shortened the increased escape latency by scopolamine. THA also significantly reduced the escape latencies compared with those in the scopolamine-treated group. On the day following the final day of training trial sessions, swimming times within the target quadrant in the scopolamine-treated group were significantly (F 5,30=5.238, P<.01) shorter than those in the control group (Fig. 2B). Moreover, the shortened swimming time within the platform quadrant was significantly increased by SMS (4 or 8 g/kg) (P<.05) or THA (P<.01) treatment.
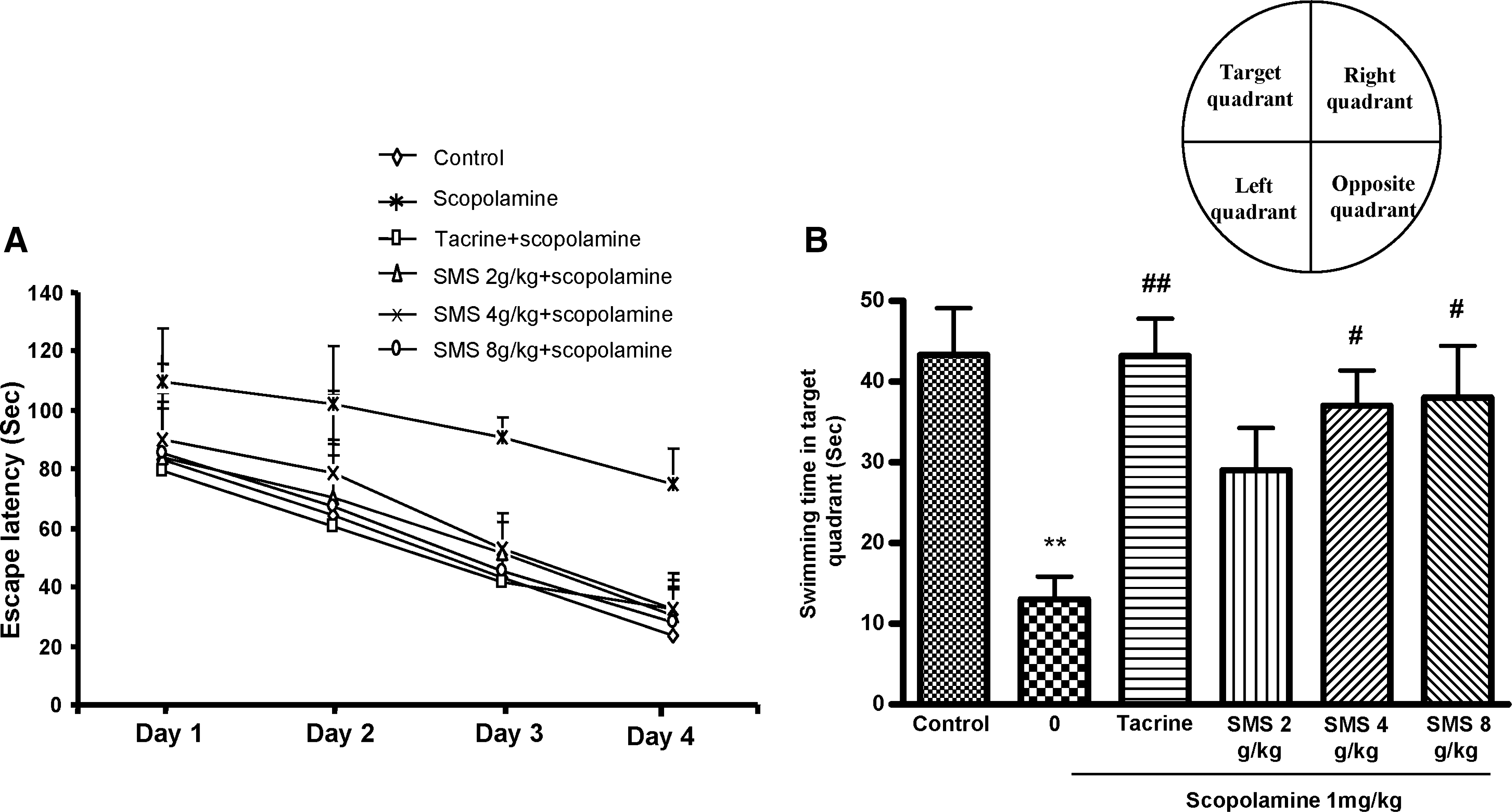
Effect of SMS on performance during (
AChE inhibitory effects of SMS
Figure 3A and B shows the effects of SMS on AChE activity in both SS and DS fractions of brain homogenates. There was a significant increase in AChE activity in both the SS (P<.01) and DS (P<.001) fractions of the scopolamine-treated group. The standard drug THA significantly (F 5,30=6.162, P<.001) reduced the AChE activity in both SS and DS fractions. Acute SMS administration resulted in a significant decrease in AChE-specific activity for both SS and DS fractions compared with those of the scopolamine-treated group. However, in both SS and DS brain homogenates, SMS at doses of 4 g/kg (54.2% and 35.5%) and 8 g/kg (55.1% and 38.7%) effectively inhibited the AChE activity, as did THA (55.3% and 40.6%).
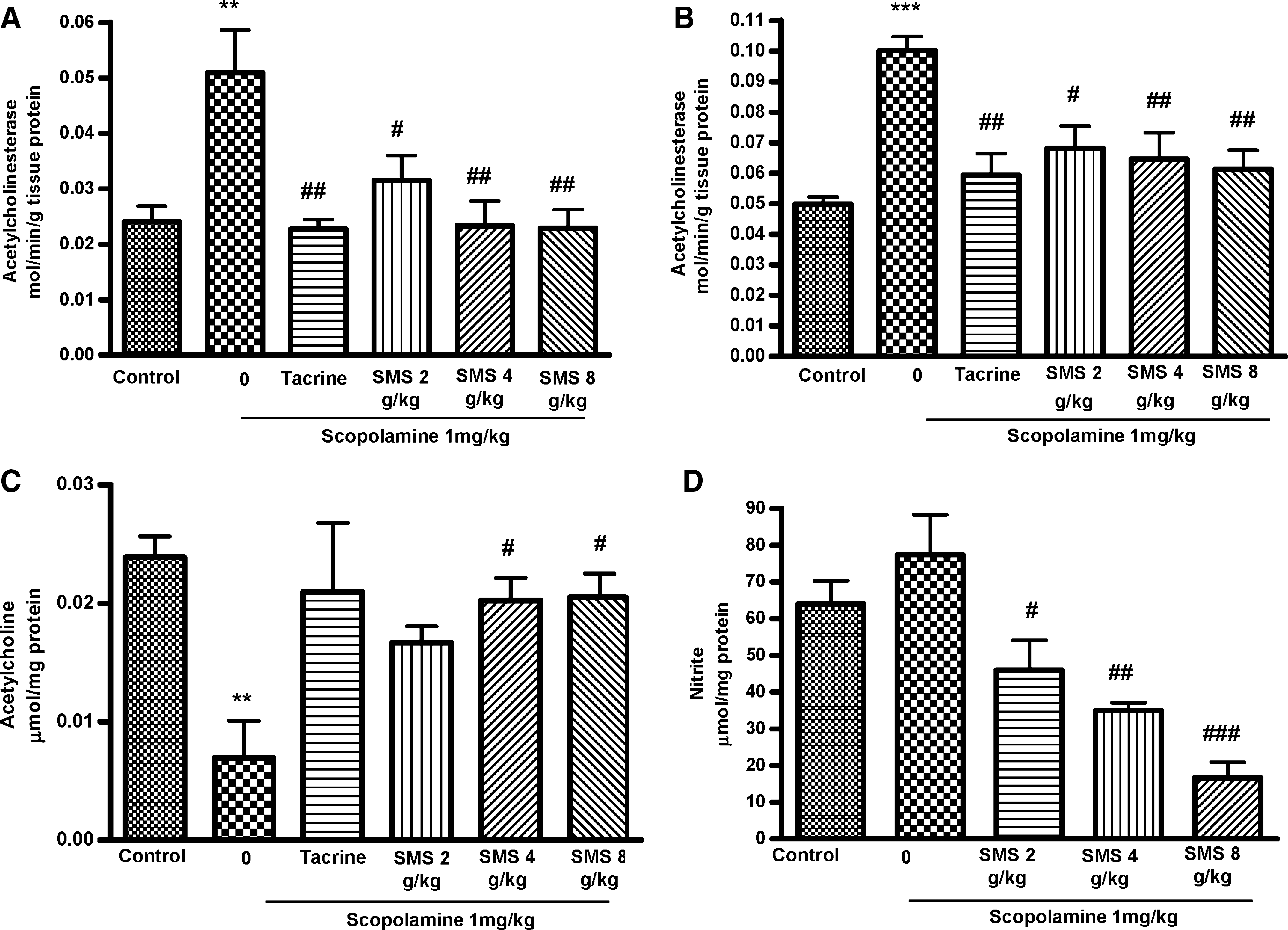
Effect of SMS (2, 4, or 8 g/kg) administration for 7 days on acetylcholinesterase activity in the (
Effect of SMS on ACh level
Figure 3C shows the effect of SMS on ACh level in brain homogenate. SMS at 4 g/kg (P<.05) and 8 g/kg (P<.05) significantly increased the ACh level compared with the scopolamine-treated group. There was a significant decrease in ACh levels in the scopolamine-treated (F 5,30=6.162, P<.01) group. The standard drug THA also significantly increased the ACh level compared with the scopolamine-treated group
Effect of SMS on nitrite level
There was no significant increase in nitrite level in the scopolamine-treated group (77.33±10.96 μmol) compared with that in the control group (64.02±6.31 μmol). However, there was a significant (F 5,30=11.36, P<.001) decrease in nitrite level in the SMS at 2 g/kg (P<.05)-, 4 g/kg (P<.01)-, and 8 g/kg (P<.01)-treated groups compared with that of the scopolamine-treated one, as shown in Figure 3D. SMS at 4 and 8 g/kg decreased nitrite levels by 54.85% and 78.3%, respectively.
Antioxidant effects of SMS on scopolamine-induced oxidative stress
To further elucidate the biochemical mechanism of the anti-amnesic activity of SMS in brain tissue, we measured their effects on lipid peroxidation and on the activities of antioxidant enzymes. The scopolamine-treated mice showed a significant increase in MDA levels (Fig. 4A) (P<.05) and significant decreases in glutathione (Fig. 4B) (P<.05) and SOD (Fig. 4D) (P<.01) levels, while the activity of GPx was also decreased, but the difference was not statistically significant (Fig. 4C). The acute treatment of amnesic mice with SMS at all doses significantly preserved the activities of GSH and SOD at higher concentration (8 g/kg) at levels comparable to those of control mice. Treatment with SMS significantly prevented the activity of GPx in amnesic brain. Furthermore, SMS treatment significantly decreased the MDA levels compared with those of the scopolamine-treated group, as shown in Figure 4.

Effects of acute SMS (2, 4, or 8 g/kg) treatment on concentrations of (
Discussion
Growing evidence shows that herbal drugs can effectively treat AD. According to the theory of traditional Chinese medicine, SMS is believed to be effective for invigorating the vital energy and producing and converging the vital essence, which is critical for physiological regulation of the circulatory system; also, SMS pretreatment was shown to suppress the cerebral oxidative stress in rats after ischemia-reperfusion. 17 In addition, SMS prevented oxidative damage of protein, lipids, DNA, and apoptosis caused by H2O2 and β-amyloid in PC-12 cells. 29 However, no attempts have been to investigate whether SMS has antioxidant or cholinergic activities in the amnesic model. Therefore, the present study was designed to investigate the effect of SMS in scopolamine-induced cognitive dysfunction and oxidative and nitrative stress and also to establish the relationship with cholinergic modulation. The findings of the present study clearly indicate that acute treatment with SMS significantly improved the recall of the learning tasks in the passive avoidance task and the MWM test. Furthermore, the biochemical investigation revealed that SMS significantly decreased the AChE activity and increased the ACh level in brain homogenates. It also exhibited antioxidative and nitrative properties, as evidenced by significant protection of different antioxidant enzymes and a decrease in nitrite levels. These results suggest that antioxidant and cholinergic signaling is involved in the ameliorating effects of SMS on cognitive dysfunctions.
Scopolamine, a muscarinic cholinergic receptor antagonist, impairs learning and memory in rodents and humans, especially the processes of learning acquisition and short-term memory. Cholinergic neurons in the central nervous system are involved in learning and memory of both humans and animals. 30,31 A major factor in age-related senile central nervous system dysfunction and the early stages of AD may be a disruption in the cholinergic neurotransmission system. One model based on this cholinergic hypothesis is the use of the muscarinic receptor antagonist, scopolamine, and an amnesic murine model as a tool for investigating age-related senile central nervous system dysfunction. 32,33 THA, the first drug approved by the Food and Drug Administration for the treatment of AD, was used as a reference drug in the present study. THA is a cholinesterase inhibitor that increases the availability of ACh in muscarinic neurons and has efficacy in mildly to moderately impaired AD patients. 34 The BALB/c mice are superior learners, showing highly structured learning-related behavior when exposed to a novel or changing situation, and have their advantage in behavioral study to the benefit of cognition. 35 In the present study scopolamine (1 mg/kg, i.p.) effectively attenuated the cognition in the BALB/c mice, and the results were in accordance with previous reports. 36,37
The passive avoidance task is a useful tool for the estimation of standard learning and memory and is used as an indicator of short- and long-term memory. 38 In the present study, reduced step-through latency by scopolamine was prevented to approximately 62% and 69% by acute treatment of SMS at 4 and 8 g/kg, respectively. THA, as a positive control, prevented the step-through latency to 77% of the vehicle-treated control group, a finding consistent with previously published data. 39 The MWM learning task was used to assess hippocampal-dependent spatial learning ability. The escape latency observed from day to day measures reference or long-term memory. 40,41 The SMS shortened the escape latencies prolonged by scopolamine treatment. At the probe trial session, SMS at 4 g/kg and 8 g/kg increased swimming time within the zone of the previously placed platform to approximately 65% and 66%, respectively, compared with that of the scopolamine group, as did THA (69%). Collectively, these behavioral studies suggest that SMS dose-dependently improves the short- and long-term memory in scopolamine-induced amnesic mouse models.
The cholinergic neurotransmission system in the basal forebrain plays an important role in learning and memory. It has been suggested that the impairments in learning, memory, and behavior observed in patients with dementia are caused, at least in part, by changes within the cholinergic system. 42,43 AChE inhibitors that increase the availability of ACh in central cholinergic synapses are used to treat AD. 38,44 Cholinergic transmission is terminated mainly by ACh hydrolysis via the enzyme AChE. This enzyme is essential for maintaining the normal function of the nervous system because it rapidly terminates the action of ACh released into the synapse, and it participates in the underlying processes in AD. 45
In this experiment, we assessed the effects of SMS on ACh level and AChE activity both with and without scopolamine-induced learning and memory impairments in the mouse brain. Scopolamine significantly reduced the ACh level in brain identically with a previous study, 46 and treatment with SMS at 4 g/kg (65%) and 8 g/kg (66%) significantly increased the ACh level, as did THA (66%). It is well documented that the AChE occurs in different molecular isoforms having differential localizations in neuronal cells. 47 Two major isoforms are globular monomer (G1) protein and globular tetramer (G4) of the same monomer subunit. The G1 isoform is reported to be present in the cytoplasm of neuronal cells, whereas the G4 isoform is predominantly membrane-bound. 48 The experimental procedure of Das et al. 21 was followed in the present study to measure each isoform in different cellular fractions. Results showed that SMS dose-dependently attenuated the AChE (G1 and G4 isoforms) levels in both SS and DS brain homogenates, and the effect was comparable to that of the reference compound THA. Altogether, our data suggest that the ameliorating effects on memory of SMS could be explained, at least in part, by their inhibition of AChE activity.
Many clinical studies have reported strong evidence that oxidative stress is involved in the pathogenesis of AD. 49 –51 In these reports, the activities of GPx and GSH were found to be elevated or changed in those brains showing severe AD pathology; this likely reflects a protective response to increased peroxidation within the brain. As such, the progression of neurodegenerative diseases was found to be inhibited by free radical scavengers and antioxidant agents. Memory impairment induced by acute scopolamine administration in rats is associated with altered levels of GSH in the brain and with the activities of antioxidant enzymes. 7 In our experimental conditions, scopolamine administration resulted in a significant increase in level of MDA, an important marker for lipid peroxidation, and in a reduction in activity of both GSH and SOD. The administration of SMS produced a significant fall in MDA content and prevented the activities of GSH and SOD in amnesic mice. Although the activity of GPx was not changed by the administration of scopolamine, treatment with SMS elevated the activity of GPx level comparable to that of normal control mice. From the above results, it could be postulated that SMS prevented cerebral oxidative damage induced by scopolamine through not only scavenging of reactive oxygen species, but also through the modulation of antioxidant substrate and enzymes such as cellular GSH and SOD levels, respectively. This is supported by our previous report in which SMS showed strong antioxidant activity for both in vitro and in vivo models. 52 Although SMS is effective in attenuating AChE activity in vivo, there was no significant AChE inhibitory action in in vitro systems (data not shown), and the precise mechanism remains to be determined. Hence the SMS is basically an antioxidant formula having secondary AChE-inhibiting action in vivo and might be a candidate for AD therapy.
In conclusion, our results suggest that SMS has anti-amnesic activity and that it may hold significant therapeutic value for alleviating memory impairments. In addition, the mechanism of anti-amnesic effects of SMS may be, partly, involved in the activation of the ACh receptor system via inhibition of the AChE system and by the regulation of the antioxidant system. This compound with an anti-AChE activity and other beneficial effects on neurodegenerative disease, including an antioxidative effect, could be a new-generation, excellent drug candidate for AD therapy. Rigorous verification of such properties might better clarify the mechanism of action of SMS and support the rationale of clinical use of this traditional formula in neurodegenerative disorders.
Footnotes
Acknowledgments
This study was supported by a grant from the Promotion and Mutual Aid Corporation for Private Schools. The authors thank the Rotary Yoneyama Doctor Course Scholarship Association. Furthermore, we thank Isukura Co., Ltd. for providing the Shengmai-san.
Author Disclosure Statement
No competing financial interests exist.