
Abstract
Metabolic dysregulation (e.g., hyperglycemia, hyperinsulinemia, hyperlipidemia, etc.) is a hallmark of obesity-related diseases such as insulin resistance, type 2 diabetes, and fatty liver disease. In this study, we assessed whether dietary capsaicin attenuated the metabolic dysregulation in genetically obese diabetic KKAy mice, which have severe diabetic phenotypes. Male KKAy mice fed a high-fat diet for 2 weeks received a 0.015% capsaicin supplement for a further 3 weeks and were compared with nonsupplemented controls. Dietary capsaicin markedly decreased fasting glucose/insulin and triglyceride levels in the plasma and/or liver, as well as expression of inflammatory adipocytokine genes (e.g., monocyte chemoattractant protein-1 and interleukin-6) and macrophage infiltration. At the same time expression of the adiponectin gene/protein and its receptor, AdipoR2, increased in adipose tissue and/or plasma, accompanied by increased activation of hepatic AMP-activated protein kinase, a marker of fatty acid oxidation. These findings suggest that dietary capsaicin reduces metabolic dysregulation in obese/diabetic KKAy mice by enhancing expression of adiponectin and its receptor. Capsaicin may be useful as a dietary factor for reducing obesity-related metabolic dysregulation.
Introduction
O
Capsaicin, a major constituent of hot pepper, enhances energy metabolism through its β-adrenergic action, 5 and it also elicits anti-inflammatory activities, 6 –9 suggesting that capsaicin may be useful for the treatment of obesity-related inflammatory metabolic complications. More recently, our previous in vitro and in vivo work has demonstrated an inhibitory effect of capsaicin on adipose tissue inflammatory responses involving reduced inflammatory adipocytokine release from adipose tissue in diet-induced obese mice. 10,11 Such an effect of capsaicin presumably also contributes to its ability to prevent impairment of glucose tolerance in obese mice fed a high-fat diet. 11 However, it remains unclear whether capsaicin counteracts the severe metabolic disorders accompanying the diabetic state. The KKAy strain of mice was created by crossing KK mice with yellow obese mice (Ay mice). Mice of this strain develop obesity and type 2 diabetes accompanied by severe hyperglycemia, hyperinsulinemia, and hypertriglyceridemia, as well as insulin resistance; 12,13 thus these mice are widely used for studying metabolic irregularities in type 2 diabetes.
In this study, we demonstrate that dietary capsaicin reduces metabolic dysregulation in obese/diabetic KKAy mice and that its effect is associated with up-regulation of adiponectin in adipose tissue and of its receptor, AdipoR2, in liver.
Materials and Methods
Animal experiments
Eight-week-old male KKAy mice were purchased from CLEA Japan (Tokyo, Japan). They were housed at 22°C and 50% humidity, with a 12-hour light/dark cycle, and fed a high-fat diet (Research Diets, New Brunswick, NJ, USA) (45% calories from lard and soybean oil, 21% energy as fat, 48% as carbohydrate, 17% as protein, and 0.15% as cholesterol) for a total of 5 weeks. After 2 weeks of feeding, the mice were divided into two groups, a control group (KKAy) and a group supplemented with 0.015% dietary capsaicin (Wako, Osaka, Japan) for another 3 weeks. During these 3 weeks, the obese controls were given the same amount of food per day as was consumed by the capsaicin-supplemented obese mice over a period of 24 hours. Both sets of mice received water ad libitum. After 5 weeks the mice were killed under CO2 anesthesia, and their adipose tissues and other organs were isolated. Samples were stored in RNAlater® (Ambion, Austin, TX), frozen in liquid nitrogen, and stored at −80°C in 10% buffered neutral formalin for histological evaluation. All animal experiments were approved by the Animal Ethics Committee of the University of Ulsan (Ulsan, Republic of Korea) and conformed to guidelines of the U.S. National Institutes of Health.
Analysis of glucose, insulin, and triglyceride contents
Glucose and triglyceride were measured by biochemical assays and/or spectrophotometrically using commercial kits (Sigma, St. Louis, MO, USA). Blood was collected in heparinized tubes, and plasma was prepared immediately by centrifugation (model 5415D microcentrifuge, Eppendorf, Haupauge, NY, USA) at 3,000 rpm and 4°C for 15 minutes. Plasma insulin levels were measured with an ultrasensitive mouse insulin enzyme-linked immunosorbent assay kit (Mercodia, Uppsala, Sweden).
Reverse transcription–polymerase chain reaction analysis
Total RNA was extracted from 50–100-mg tissue samples using TRIzol® reagent (Invitrogen, Carlsbad, CA, USA). Total RNA (0.5 μg) was used for reverse transcription (RT); it was amplified by polymerase chain reaction (PCR) in a single reaction, using the Access RT-PCR system in a TaKaRa thermal cycler (TaKaRa Biomedicals, Tokyo). For semiquantitative analyses, MCP-1, IL-6, and adiponectin cDNAs were amplified, normalized to the mRNA level of the housekeeping gene 36B4, and expressed as ratios. 6 The linearity of the amplification of AdipoR1, AdipoR2, and β-actin was established in preliminary experiments. The following primer sequences obtained from the GeneBank database were used in the PCR amplification: mouse AdipoR1 (NM_028320), forward 5′-TACCTGCTACATGGCCACAGAC-3′, reverse 5′-CAGACGATGGAGAGGTAGATGAG-3′; and mouse AdipoR2 (NM_197985), forward 5′-CTACAGGCCCATCATGCTATG-3′, reverse 5′-TGAGAGACGATAATGGCTGC-3′). The amplification products were electrophoretically separated on 1.5% agarose gels, and the bands stained with SYBR® Green (Invitrogen, Eugene, OR, USA) were photographed with a Canon (Tokyo) PowerShot A640 camera. Band intensities were measured with an ImageJ analyzer (Biocompare, San Francisco, CA, USA). Levels of AdipoR1 and AdipoR2 mRNAs were normalized to the mRNA level of the β-actin control gene and expressed as fold of the control.
Measurement of MCP-1, IL-6, TNF-α, and adiponectin proteins
Levels of MCP-1, IL-6, TNF-α, and adiponectin were measured by enzyme-linked immunosorbent assay. The assays were conducted with an OptEIA™ (BD Bioscience Pharmingen, San Diego, CA, USA) mouse MCP-1 set, a mouse TNF-α set (BD Biosciences Pharmingen), a mouse IL-6 set (R&D Systems, Minneapolis, MN, USA), and a mouse adiponectin set (R&D Systems). Samples were thawed, diluted appropriately with assay diluent, and assayed. MCP-1, IL-6, and TNF-α were quantified using a standard curve obtained with the SOFTmax curve-fitting program (Molecular Devices, Sunnyvale, CA, USA). Amounts of cytokine were normalized for protein content, and the protein content of homogenates was determined using the BCA protein assay (Pierce, Rockford, IL, USA).
Flow cytometric analysis
Stromal vascular cells isolated from adipose tissue were incubated in the dark at 4°C on a bidirectional shaker for 30 minutes in Fc blocking solution (eBioscience, San Diego) and then stained with phycoerythrin-conjugated anti-mouse F4/80 (eBioscience) and/or fluorescein isothiocyanate-conjugated anti-mouse CD11b (eBioscience). After incubation with the antibodies, 1 mL of flow cytometric analysis (fluorescence-activated cell sorting) buffer was added to the cells. The cells were centrifuged at 1,200 rpm (model HBT15-8 centrifuge, Hanil Inc., Incheon, Republic of Korea) for 5 minutes and resuspended in 1 mL of fluorescence-activated cell sorting buffer. Washing was repeated twice, and the cells were analyzed on a FACSCalibur™ (BD Biosciences) with CellQuest software (BD Biosciences).
Western blot analysis
Total protein (20–50 μg) was subjected to western blot analysis using polyclonal antibodies. Liver AMP-activated protein kinase (AMPK) activity was detected with polyclonal antibodies to phosphorylated AMPK (AMPK-pThr172) and nonphosphorylated AMPK (Cell Signaling).
Histological analysis
Adipose tissue and liver, fixed in 10% formalin and embedded in paraffin, were cut into 8-μm-thick sections, which were mounted on two glass slides, stained with hematoxylin–eosin, and viewed with an Olympus (Tokyo) microscope at × 100 magnification.
Statistical analyses
Results are expressed as mean ± SEM values. Statistical analysis was performed with analysis of variance and Duncan's multiple-range tests. Differences were considered significant at P < .05.
Results
Body weight, adiposity, and plasma metabolic parameters
There was no significant difference in body weight gain and adiposity between the KKAy mice supplemented with 0.015% capsaicin (KKAy-CAP, 42.37 ± 0.70 g) and controls (KKAy, 45.12 ± 1.42 g) over the experimental period (Fig. 1A and B). To test whether dietary capsaicin reduced metabolic dysregulation, we compared fasting glucose, insulin, and triglyceride levels in the two groups. Levels of fasting glucose (Fig. 1C), insulin (Fig. 1D), and triglyceride (Fig. 1E) were significantly lower in the capsaicin-supplemented group. These findings suggest that dietary capsaicin reduces metabolic complications such as hyperglycemia, hyperinsulinemia, and hypertriglyceridemia in the obese diabetic KKAy mice and that capsaicin action is independent of body weight and/or fat mass reduction.
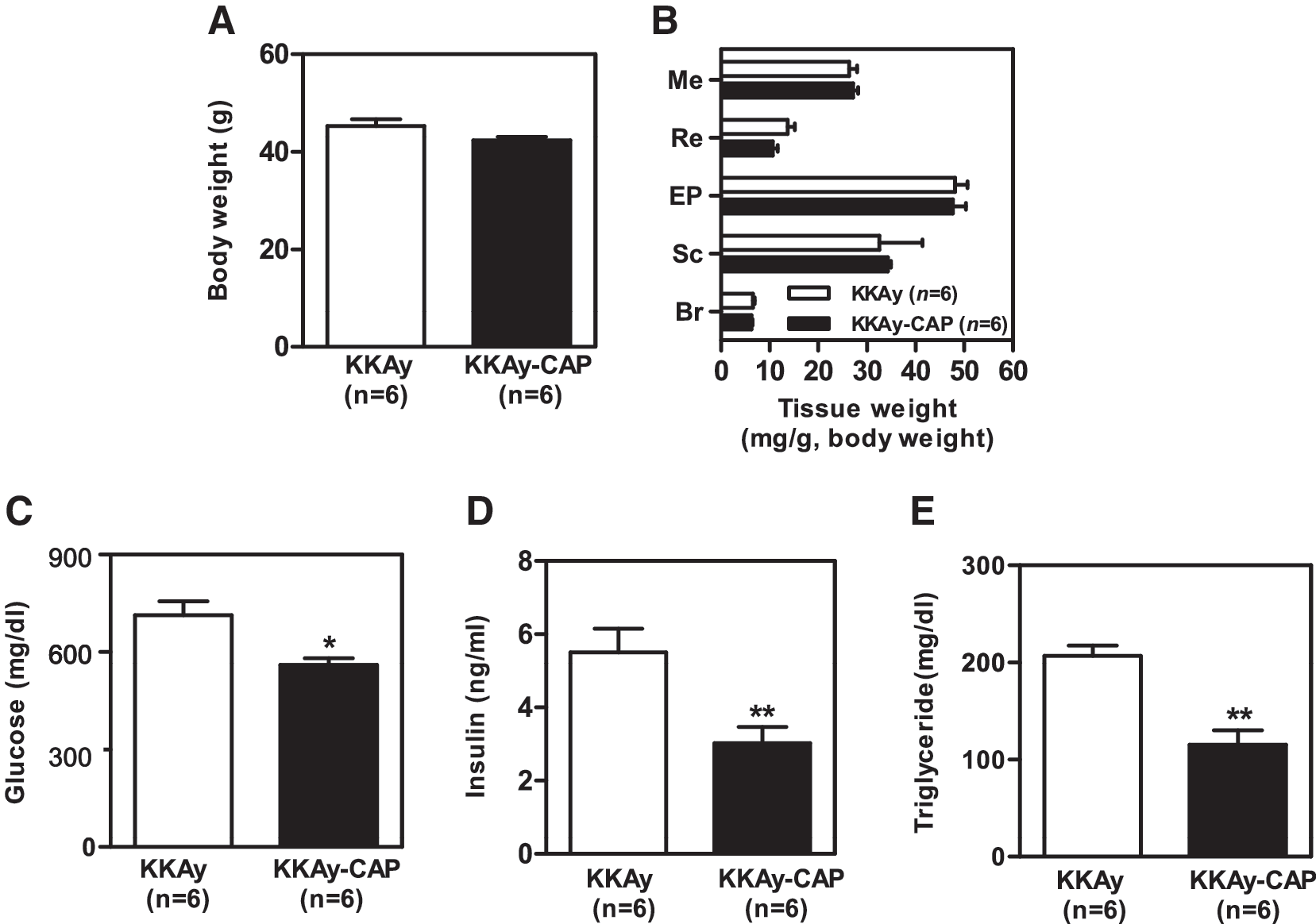
Effect of dietary capsaicin on (
Fatty liver formation and hepatic/adipose tissue inflammatory responses
Insulin resistance and diabetes trigger hepatic steatosis, which is associated with hepatic inflammation. To examine whether dietary capsaicin attenuates hepatic steatosis and inflammation, we measured hepatic triglyceride and inflammatory cytokine levels. Histological analysis revealed that lipid droplets in the liver, which were numerous in the obese controls, were reduced by capsaicin supplementation (Fig. 2A), and this was confirmed by the decrease in triglyceride content (Fig. 2B). Levels of IL-6 and MCP-1 mRNAs in the liver were also significantly lower in the capsaicin-supplemented group (Fig. 2C). These findings indicate that dietary capsaicin reduces hepatic steatosis and inflammation in the KKAy mice. To examine whether dietary capsaicin inhibits adipose tissue inflammatory responses, we examined the extent of macrophage infiltration into adipose tissue. Histochemical examination revealed fewer infiltrated cells in the adipose tissue of the capsaicin-supplemented mice than in that of the controls (Fig. 2D). The macrophage population (F4/80 + /CD11b+) was approximately 15% lower in the adipose tissue of the supplemented mice (Fig. 2E). We also measured adipocytokine gene expression in adipose tissue by RT-PCR. As shown in Figure 2F, MCP-1 mRNA was significantly lower in the capsaicin-supplemented mice, and IL-6 expression levels also showed a tendency to be lower. These findings indicate that dietary capsaicin reduces hepatic/adipose tissue inflammatory responses in obese/diabetic conditions.
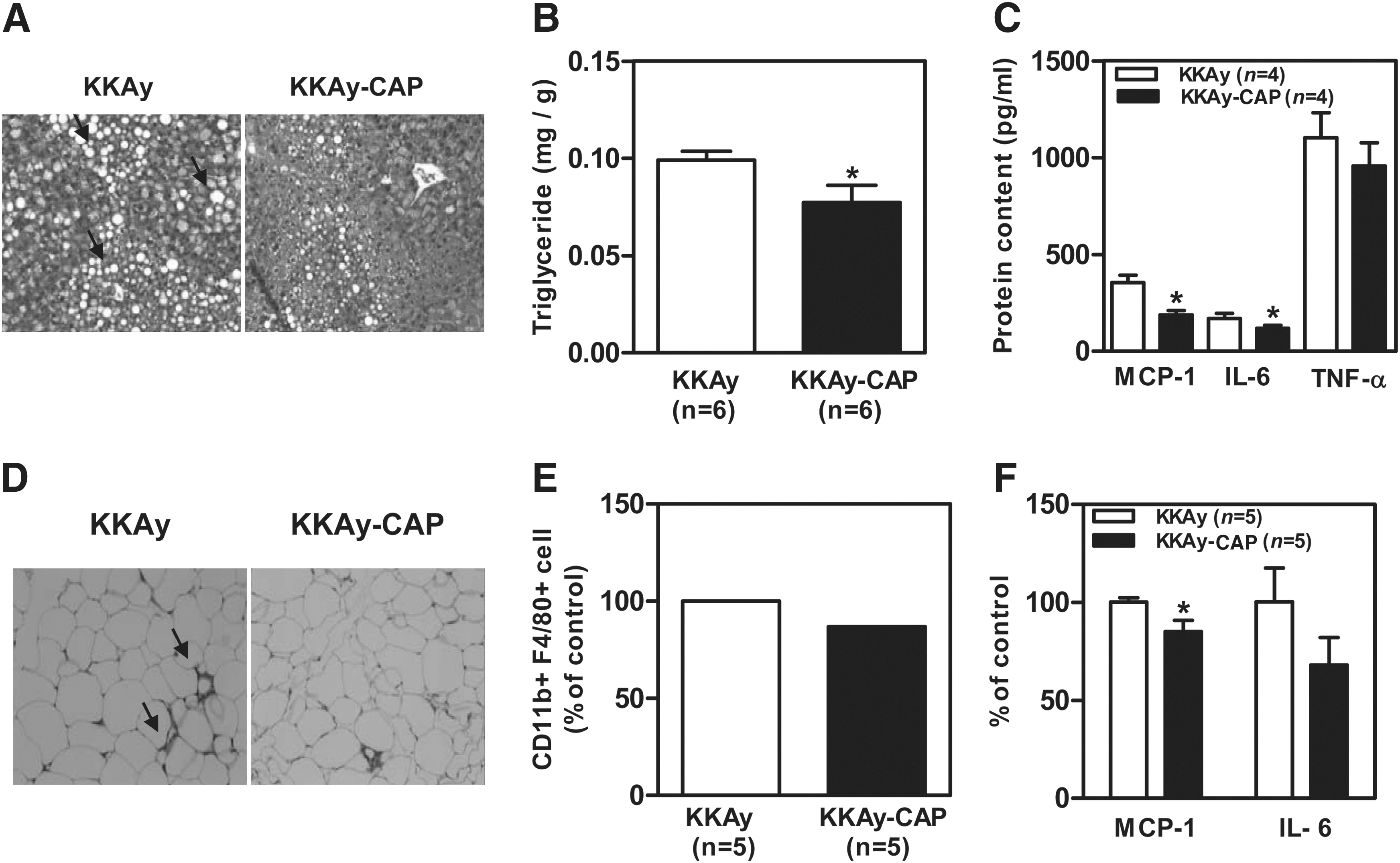
Protective effect of dietary capsaicin on fatty liver formation and hepatic/adipose tissue inflammatory responses. (
Expression of the genes for adiponectin and its receptor and activation of AMPK
To see whether the effects of capsaicin were associated with an action on adiponectin, we assessed the expression of the genes for adiponectin and its receptor (AdipoR1 and R2) and the levels of the corresponding proteins. The levels of adiponectin protein in plasma (Fig. 3A) and adiponectin mRNA in adipose tissue (Fig. 3B) were significantly higher in the capsaicin-supplemented mice. Capsaicin supplementation up-regulated AdipoR2 (Fig. 3C), but not Adipo R1 (data not shown) and caused a significant increase in the level of activated AMPK in the liver (Fig. 3D).
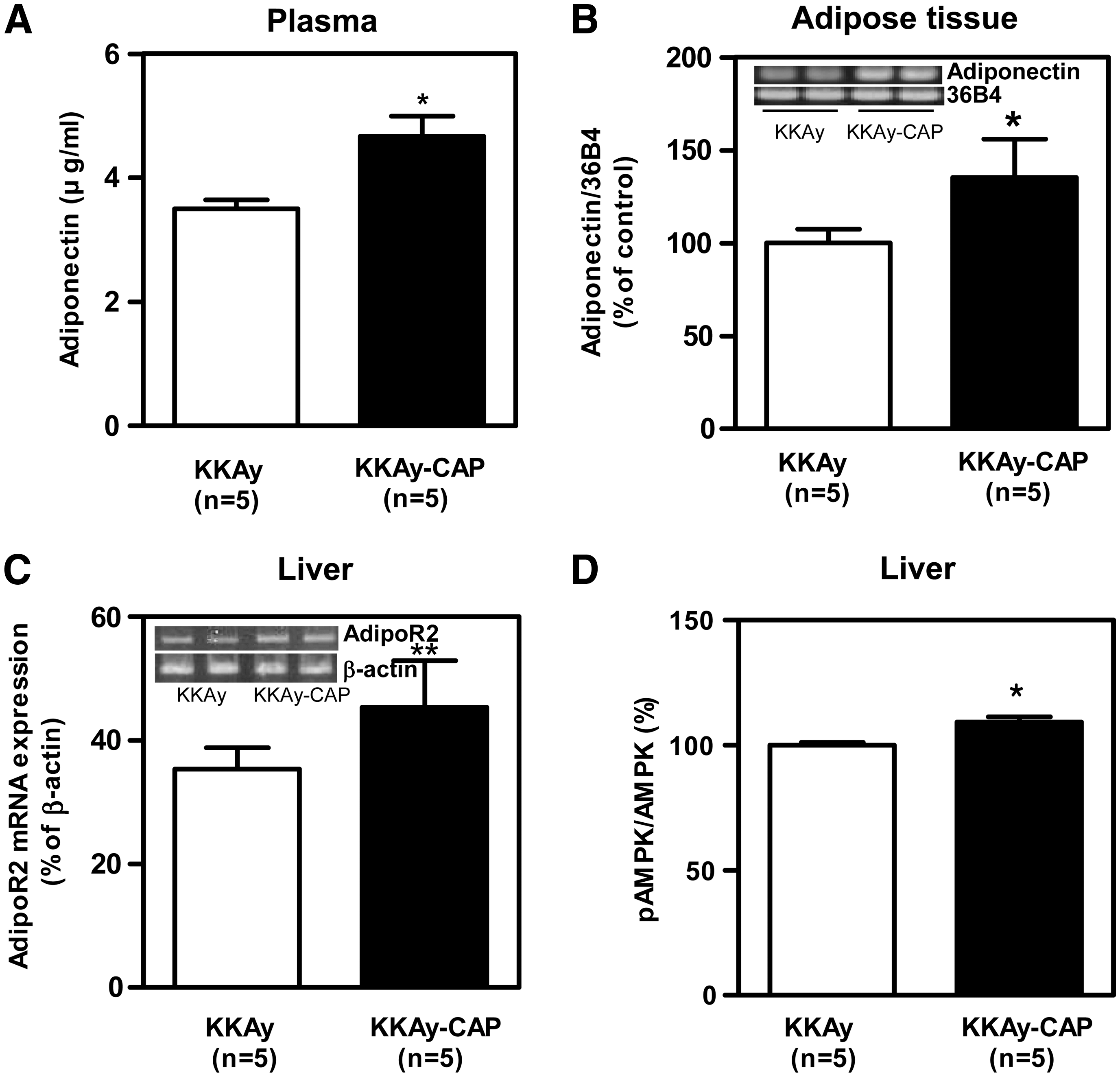
(
Discussion
Inflammation and metabolic dysregulation (e.g., hyperglycemia, hyperinsulinemia, and hypertriglyceridemia) are hallmarks of obesity-related disorders such as insulin resistance, type 2 diabetes, atherosclerosis, and fatty liver. In this study, we demonstrated that dietary capsaicin markedly attenuated hyperglycemia, hyperinsulinemia, and hypertriglyceridemia in genetically obese/diabetic KKAy mice and that this was accompanied by a reduction in hepatic triglyceride content. Adipose tissue and hepatic inflammatory responses were also reduced. Interestingly, it appears that the effect of capsaicin was independent of any reduction in body weight and adiposity. Because the metabolic disturbance in KKAy mice is more severe than in C57BL/6 obese mice, our findings suggest that capsaicin may be useful for the treatment of the metabolic complications associated with obese/diabetic conditions.
Adiponectin plays a pivotal role in lipid and glucose metabolism and thus influences glucose homeostasis and insulin sensitivity. 2,14 Increases in plasma adiponectin concentrations are associated with improvements in metabolic disturbances, including glucose intolerance, insulin resistance, fatty liver, and dyslipidemia. 15,16 Thus, up-regulating adiponectin could be an attractive strategy for protecting against metabolic dysregulation. We found that plasma adiponectin and the level of adiponectin mRNA in adipose tissue increased in response to dietary capsaicin, suggesting that up-regulation of adiponectin gene expression in adipose tissue contributes to increased plasma adiponectin levels. Several lines of evidence have suggested that peripheral tissue adiponectin receptors are also useful therapeutic targets. 17 Adiponectin receptors consist of AdipoR1 and R2. Both are expressed in liver and adipose tissue 2 and have been shown to be down-regulated in obese diabetic KKAy mice. 18 Interestingly, we observed that liver AdipoR2 mRNA increased in response to capsaicin supplementation. In liver, adiponectin binds mainly to membrane-bound AdipoR2 and activates AMPK and peroxisome proliferation-activated receptor α pathways, leading to reductions in acetyl-coenzyme A carboxylase and fatty acid synthase activities and induction of carnitine palmitoyl transferase I activity, thereby increasing oxidation of nonesterified fatty acids and decreasing de novo lipogenesis. 19 We observed that hepatic AMPK activation increased in the capsaicin-supplemented mice; this presumably reflected the decrease in hepatic triglyceride content and led to the decreased fatty liver formation. Given the increase in the systemic levels of adiponectin and of AdipoR2 in liver, the decreased hepatic fat accumulation with dietary capsaicin may be attributed to stimulation of adiponectin action via its interaction with AdipoR2. In addition to modulating metabolic activity, adiponectin also reduces inflammation by inhibiting the production of cytokines and reactive oxygen species and reducing inflammatory nuclear factor κB signaling, oxidative stress, and cell injury. 20,21 Hence a drop in the level of adiponectin is thought to increase adipose tissue inflammatory responses, and we found that dietary capsaicin suppressed inflammatory responses in the adipose tissue and liver of KKAy mice. This effect may at least in part be due to the increase in adiponectin action.
In conclusion, dietary capsaicin counteracts metabolic dysregulation in obese diabetic KKAy mice, and this effect is associated with up-regulation of adiponectin and AdipoR2 expression. Capsaicin may be useful as a dietary additive to reduce obesity-related metabolic disturbances.
Footnotes
Acknowledgments
This work was supported by the 2007 Research Fund of the University of Ulsan. T.K. was supported by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sport, Science and Technology of Japan (22228001).
Author Disclosure Statement
No competing financial interests exist.