
Abstract
Amentoflavone, a biflavonoid from Selaginella tamariscina, is known to possess several bioactivities such as antitumor, anti-inflammatory, and antifungal effects. However, the mechanism of the anticancer effects of amentoflavone on human cervical cancer cells has not been studied in detail. In this study, we demonstrated that amentoflavone induces apoptosis in SiHa and CaSki cervical cancer cells by suppressing human papillomavirus protein E7 expression. The cyclins and tumor suppressors were modulated by amentoflavone in SiHa and CaSki human cervical cancer cells: cyclin and hyperphosphorylated retinoblastoma (p-pRb) were down-regulated, whereas cyclin-dependent kinase inhibitors and p53 were enhanced. Amentoflavone up-regulated peroxisome proliferator-activated receptor γ (PPARγ) and phosphatase and tensin homolog deleted on chromosome 10 (PTEN) expression levels while inhibiting E7-mediated cyclooxygenase-2 (COX-2)/interleukin-32 (IL-32) expressions were downregulated, and Akt phosphorlylation was decreased in an amentoflavone-induced apoptotic process, suggesting that amentoflavone may be a PPARγ activator. Additionally, the expression of the anti-apoptotic factor Bcl-2 was decreased, whereas that of the well-known apoptotic factor Bax was increased, thereby releasing cytochrome c into cytosol in amentoflavone-treated cervical cancer cells. Furthermore, amentoflavone treatment led to the activation of caspase-3 and -9 and proteolytic cleavage of poly(ADP-ribose) polymerase. The expression level of the extrinsic death receptor Fas (CD95) was not altered by amentoflavone treatment. When these findings are taken together, the biflavonoid amentoflavone activates PPARγ/PTEN expressions and induces apoptosis via suppressing E7 expression, cell cycle arrest at sub-G1 phase, and mitochondria-emanated intrinsic pathways in SiHa and CaSki human cervical cancer cells. These findings suggest that amentoflavone has potential for development as a therapeutic agent for human cervical cancer.
Introduction
C
Flavonoids, a class of plant secondary metabolites, exhibit a variety of activities including antibacterial, antiviral, antioxidant, and anticancer activities. 8 Many flavonoids possess antitumor activity against various human cancer cell lines. 9,10 These flavonoids also can act as agonists of nuclear receptor family members such as peroxisome proliferator-activated receptors (PPARs), retinoic acid receptors, and estrogen receptors. 11 PPARs, one of the nuclear receptors, play pivotal roles in the regulation of cellular differentiation, development, metabolism, and tumorigenesis. 12 PPARs are classified into three subtypes: PPARα, PPARβ/δ, and PPARγ. Each subtype is considered as an ideal target of cancer therapy, and PPARγ is the best studied. 13 Thus, the relationship between flavonoids and PPARγ is significantly connected to the promotion of anticancer effects.
In this study, the effect of amentoflavone on SiHa and CaSki human cervical cancer cells was examined. The biflavonoid amentoflavone, isolated from Selaginella tamariscina, exhibits several well-known biological activities including antitumor and anti-inflammatory properties. 14,15 However, the precise mechanism of the anticancer effects of amentoflavone in human cervical cancer cells remains obscure. Here we demonstrate that amentoflavone inhibited cell proliferation through PPARγ activation, thereby inducing sub-G1 cell cycle arrest, and also induced apoptotic cell death via inhibition of E7-mediated cyclooxygenase-2 (COX-2)/interleukin-32 (IL-32) and Akt-mediated pathways.
Materials and Methods
Reagents
CellTiter 96® AQueous One Solution cell proliferation assay [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt (MTS)] reagent was purchased from Promega (Madison, WI, USA). Phenazine methosulfate, Hoechst stain solution, and propidium iodide were from Sigma (St. Louis, MO, USA). Antibodies specific to cyclin A, cyclin E, and hyperphosphorylated Rb (p-pRb) were purchased from BD Biosciences (San Diego, CA, USA). Antibodies specific to poly(ADP-ribose) polymerase (PARP), caspase-3, caspase-9, Bax, Bcl-2, cytochrome c, phosphatase and tensin homolog deleted on chromosome 10 (PTEN), and anti-mouse immunoglobulin G–horseradish peroxidase were purchased from Cell Signaling Technology (Beverly, MA, USA). Antibodies specific to nuclear factor-κB (NF-κB) p50, p21, p27, glyceraldehyde 3-phosphate dehydrogenase (GAPDH), phospho-Akt1/2/3 (Ser-473), Akt1, PPARγ, and anti-goat immunoglobulin G–horseradish peroxidase were from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-p53 was from Oncogene (Cambridge, MA, USA), and anti-rabbit immunoglobulin G–horseradish peroxidase was from Assay Designs (Ann Arbor, MI, USA).
Extraction and isolation of amentoflavone
The whole plant of S. tamariscina (600 g) was extracted with methanol at room temperature, yielding 50.54 g of residue. The methanol extract was resuspended in water and partitioned sequentially with dichloromethane, ethyl acetate, and n-butanol. Based on their thin-layer chromatography pattern, the fractions were combined to yield subfractions, which were designated E1–E10. Subfraction E7 (296.33 mg) was finally purified by repeated column chromatography, yielding amentoflavone (82.23 mg). The ultraviolet, infrared, and 1H and 13C nuclear magnetic resonance (NMR) data for amentoflavone were identical to those reported in the literature: 16 1H-NMR (CD3OD, 500 MHz), δ 6.50 (1H, s, H-3), 6.15 (1H, d, J=2.0 Hz, H-6), 6.38 (1H, d, J=2.0 Hz, H-8), 7.91 (1H, d, J=2.0 Hz, H-2′), 7.03 (1H, d, J=8.0 Hz, H-5′), 7.75 (1H, d, J=8.0 Hz, H-6′), 6.50 (1H, s, H-3′′), 6.31 (1H, s, H-6′′), 7.43 (2H, d, J=8.5 Hz, H-2′′′, H-6′′′), 6.66 (2H, d, J=8.5 Hz, H-3′′′, H-5′′′); 13C-NMR (pyridine-d5, 125 MHz), δ 165.01 (C-2), 104.44 (C-3), 183.41 (C-4), 163.43 (C-5), 100.34 (C-6), 164.95 (C-7), 95.19 (C-8), 158.85 (C-9), 105.38 (C-10), 122.08 (C-1′), 128.66 (C-2′), 122.83 (C-3′), 161.66 (C-4′), 117.60 (C-5′), 132.92 (C-6′), 166.15 (C-2′′), 103.92 (C-3′′), 183.06 (C-4′′), 162.77 (C-5′′), 100.14 (C-6′′), 164.06 (C-7′′), 105.61 (C-8′′), 156.19 (C-9′′), 105.27 (C-10′′), 122.76 (C-1′′′), 129.16 (C-2′′′), 117.18 (C-3′′′), 162.98 (C-4′′′), 117.18 (C-5′′′), 129.16 (C-6′′′); high-resolution fast atom bombardment–mass spectrometry, m/z 539.1603 (calculated for C30H19O10 [M+H]+, 539.1622); infrared νmax (KBr) cm−1, 3200, 1660, 1605, 1350, 1160, 830.
Cell culture
High-risk HPV-16 genome containing human cervical cell lines (SiHa and CaSki) and HPV-free C33A cervical cancer cells with mutated p53, obtained from the American Type Culture Collection (Rockville, MD, USA), were cultured in Dulbecco's modified Eagle's medium (Hyclone Laboratories, Logan, UT, USA) supplemented with 2 mM
Cell viability assay and cell morphological assessment
In order to test the effect of amentoflavone on cell growth, approximately 0.5×104 cells per well were plated onto 96-well microtiter plates, grown overnight, and treated with amentoflavone dissolved in dimethyl sulfoxide (final concentration 0.1%) for 24 and 48 hours. Cell viabilities were analyzed by MTS assay according to the manufacturer's instruction. The optical absorbance was measured at 492 nm using a microplate reader (Apollo LB 9110, Berthold Technologies GmbH, Bad Wildbad, Germany). Morphology of the cells was observed under the inverted phase-contrast microscope. Apoptotic morphological changes were detected by Hoechst staining. Cells plated on coverslips were treated with amentoflavone and incubated for 24 hours. The coverslips were washed with phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde for 1 hour at room temperature. After washing with PBS, the fixed cells were stained for 20 minutes with Hoechst staining solution at 37°C. After washing with PBS, the coverslips were completely dried and mounted on microscope slides. The slides were then observed by fluorescent microscopy.
Reverse transcription–polymerase chain reaction and real-time quantitative polymerase chain reaction analysis
Total RNAs were isolated using the easy-BLUE™ total RNA extraction kit (iNtRon Biotechnology, Seoul, Korea), and the cDNA products were prepared with M-MuLV reverse transcriptase (New England Biolabs, Beverly). Reverse transcription (RT)–polymerase chain reaction (PCR) analysis was conducted using a Dice PCR thermal cycler (TaKaRa, Shiga, Japan) with the following primer sets: HPV-E7, 5′-ATGCATGGAGATACACCTACATTGC-3′ (forward) and 5′-TTATGGTTTCTGAGAACAGATGGGGC-3′ (reverse); IL-32, 5′-ATGTGCTTCCCGAAGGTCCTC-3′ (forward) and 5′-TCATTTTGAGGATTGGGGTTC-3′ (reverse); COX-2, 5′-GAAACCCACTCCAAACACAG-3′ (forward) and 5′- CCCTCGCTTATGATCTGTCT-3′ (reverse); and GAPDH as an internal control. Real-time quantitative PCR (qPCR) was performed with a relative quantification protocol on a Chromo 4 Real-Time PCR system (Bio-Rad, Hercules, CA, USA), using iQ™ SYBR® Green Supermix (Bio-Rad). All target genes were normalized to the housekeeping gene, GAPDH. Each sample was assayed in triplicate with the following primer set: PPARγ, 5′-CCTATTGACCCAGAAAGCGA-3′ (forward) and 5′-CCATTACGGAGAGATCCACG-3′ (reverse). Fold changes represent the ratio of PPARγ expression in SiHa and CaSki cells treated with amentoflavone to the control.
Western blot analysis
The cells were lysed in a lysis buffer containing 0.1% sodium dodecyl sulfate, 0.1% sodium deoxycholate, 1% Triton X-100, 1 mM EDTA, 0.5 mM EGTA, 140 mM NaCl, 10 mM Tris-HCl (pH 8.0), 10 μg/mL aprotinin, and 0.5 mM phenylmethylsulfonyl fluoride. Cytosolic fractions were obtained with cytosolic lysis buffer containing 10 mM HEPES-KOH (pH 7.9), 10 mM KCl, 2 mM NaCl2, 0.1 mM EDTA, 0.2 mM NaF, 0.4 mM phenylmethylsulfonyl fluoride, 0.1 mM Na3VO4, 1 mM dithiothreitol, 10 μg/mL aprotinin, and 0.5 mM phenylmethylsulfonyl fluoride. Protein concentrations were determined using a Bio-Rad protein assay kit; proteins were subjected to 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred to polyvinylidene difluoride membranes (Millipore, Billerica, MA, USA). The membranes were blocked with 5% nonfat dry milk dissolved in 140 mM NaCl, 27 mM KCl, 10 mM Na2HPO4·12H2O, 1.8 mM KH2PO4, and 0.05% Tween-20. Primary antibodies specific to COX-2, Akt, phospho-Akt (Ser-473), GAPDH, p21, p27, PARP, caspase-3, -8, and -9, Bax, Bcl-2, cytochrome c, p-pRb, cyclin E, cyclin A, p53, and IL-32 (KU32-52) 17 were applied to the membranes. Horseradish peroxidase–conjugated secondary antibodies were used to visualize the bound primary antibodies with the Westzol® Plus Western Blot detection system (iNtRON Biotechnology).
Cell cycle analysis by flow cytometry analysis
Approximately 3×105 cells per well were plated onto six-well plates and incubated overnight to adhere. The cells were then treated with amentoflavone. After 24 hours, the cells were harvested and fixed with 70% ethanol at −20°C. Then, the fixed cells were washed with PBS prior to staining. The cells were stained for 30 minutes with PBS containing 50 μg/mL propidium iodide and 100 μg/mL RNase A. The proportion of apoptotic cells was analyzed by FACSCalibur™ and CellQuest™ software (BD Biosciences).
Statistical analysis
The data were expressed as mean±SD values. Statistical analysis was performed using Student's t test, with the following significance levels: **P<.001.
Results
Antiproliferative and apoptotic effects of amentoflavone in human cervical cancer cells
The amentoflavone was isolated from S. tamariscina (Fig. 1A). We first examined the antiproliferative effects of amentoflavone on HPV-positive human cervical cancer cell lines, SiHa and CaSki cells. To maximize the specificity and efficiency of the amentoflavone effect, we chose the CaSki cells, which contain 500–600 copies of integrated HPV-16 genome, compared with SiHa cells, containing only one or two copies of HPV-16. 18 Ametoflavone was more cytotoxic to CaSki cells (Fig. 1B), suggesting that CaSki cells were more sensitive to amentoflavone than SiHa cells because CaSki cells contain more copies of integrated HPV-16 genome compared with SiHa cells. The cells were exposed to different concentrations of amentoflavone for 24 and 48 hours, after which cell viability was measured by MTS assay. We did not investigate the DNA ladder and nuclear changes regarding the normal cell lines because amentoflavone did not show any significant cytotoxic effects on the growth of normal keratinocytes, HaCaT cells (data not shown), and HPV–non-harboring C33A cervical cancer cells (Fig. 1C). Therefore, we focused on the cell proliferation and on nuclear modifications such as distinct condensed chromatin and fragmentation by nuclear staining with Hoechst in amentoflavone-treated SiHa and CaSki cervical cancer cells. The growths of both SiHa and CaSki cells were significantly inhibited in a time- and dose-dependent manner, and morphological changes were apparent under inverted phase-contrast microscopy (Fig. 1B and D). It was then ascertained whether nuclear morphology would be altered by treatment with amentoflavone. Nuclear staining with Hoechst revealed morphological changes of SiHa and CaSki cells treated with amentoflavone, and nuclear modifications such as distinct condensed chromatin and fragmentation were noted at 40 μM and 80 μM (Fig. 1D). These results indicate that amentoflavone inhibits cell proliferation and induces the apoptotic death in the human cervical cancer cells SiHa and CaSki.
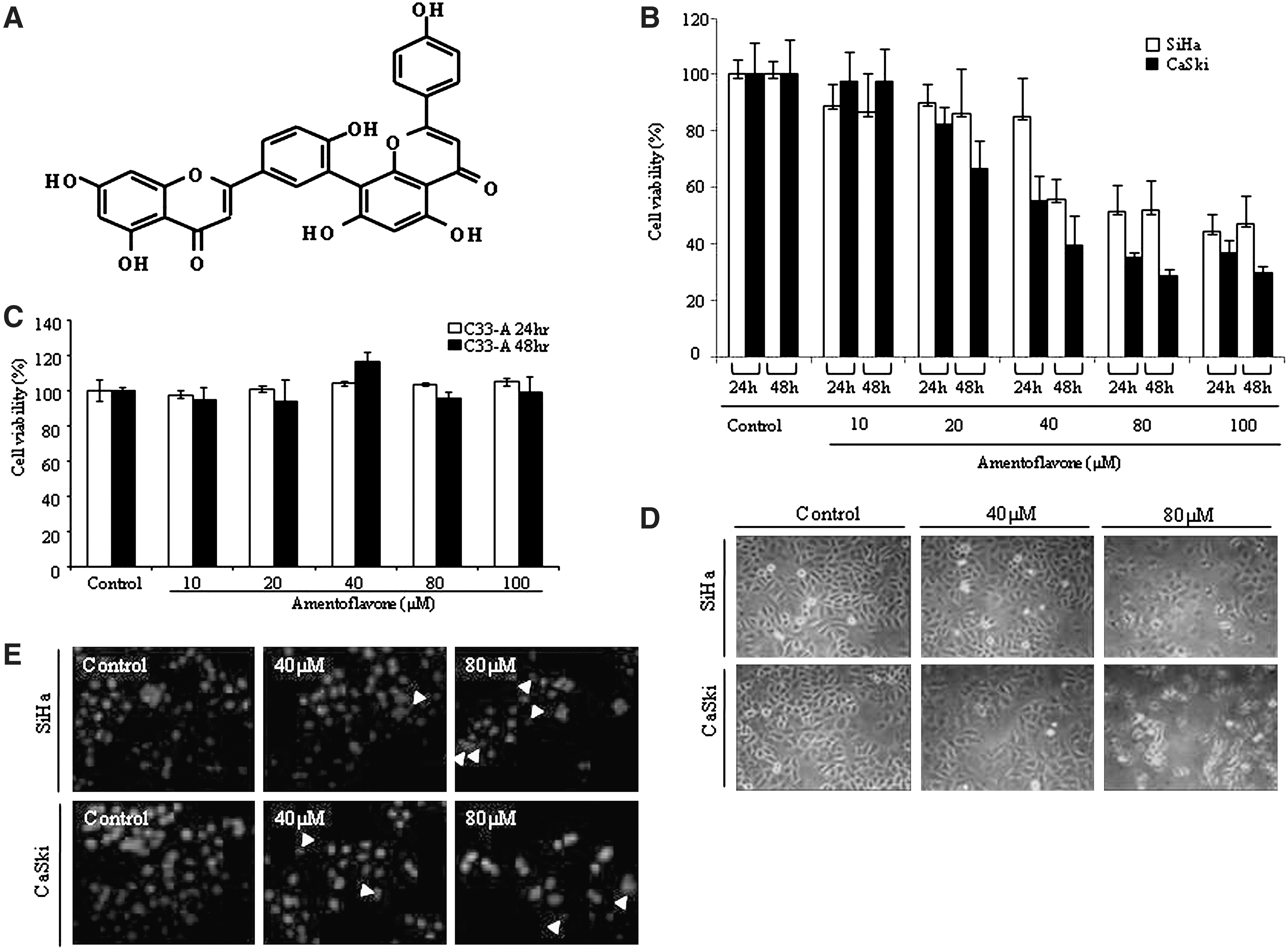
Amentoflavone-induced antiproliferative and apoptotic effects in SiHa and CaSki human cervical cancer cells. (
Amentoflavone-induced sub-G1 phase cell cycle arrest in SiHa and CaSki cells
In order to examine whether the antiproliferative effect of amentoflavone is related to cell cycle arrest, SiHa and CaSki cells were treated with or without amentoflavone for 24 hours, and then cell cycle distribution was measured by flow cytometry. The SiHa cells that accumulated at sub-G1 phase were increased from 7.02% to 49.33%, and the proportion of CaSki cells was increased from 8.14% to 51.89%. However, the populations of G0/G1 cell were decreased from 59.60% to 26.75% in SiHa cells and from 58.30% to 27.99% in CaSki cells, upon amentoflavone treatment for 24 hours (Fig. 2A). However, amentoflavone increased the fraction at G2/M at 40 μM in SiHa cells but not in CaSki cells, whereas the fractions at G2/M at 80 μM were slightly decreased in SiHa and CaSki cells (Fig. 2A). These results led to the postulation that amentoflavone induces apoptotic cell death through cell cycle arrest mostly at the sub-G1 phase in SiHa and CaSki cells. Based on the cell cycle dynamics, the regulation of G1 phase-associated molecules was then analyzed by western blot. As shown in Figure 2B, the expression levels of cyclin E and cyclin A were reduced in amentoflavone-treated cells. The expression level of p-pRb protein, which is regulated by E7 and G1 cyclins (cyclin-dependent kinases), was also reduced. Next the upstream regulators of cell cycle, p21 and p27, which are involved in the regulation of G1/S cyclins and p53 tumor suppressor, were examined. Amentoflavone treatment clearly elevated p21, p27, and p53 levels in SiHa and CaSki cells. In order to investigate whether amentoflavone might induce p21 through a p53-independent pathway, C33A cervical cancer cells with mutated p53 were used. Amentoflavone did not inhibit the growth of C33A cells (Fig. 1C). Moreover, neither p53 nor p21 expressions were altered in C33A cells (Fig. 2B). These results suggest that amentoflavone induces the down-regulation of G1/S cyclins and p-pRb and the up-regulation of p27 and p21 via a p53-dependent pathway in SiHa and CaSki cells.
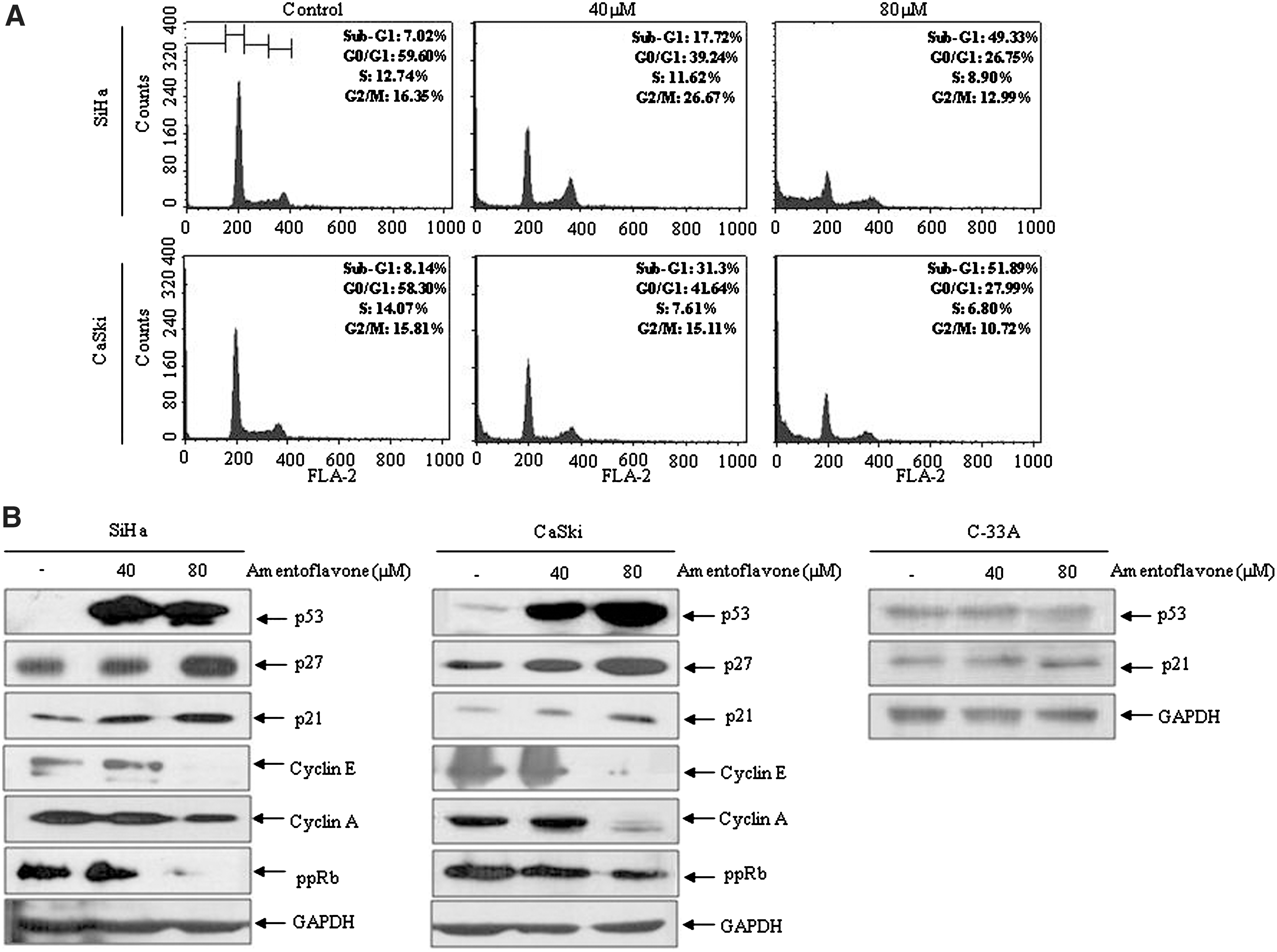
Effects of amentoflavone on cell cycle progression and expression of cell cycle regulators in cervical cancer cells. The cells were treated with different concentrations for 24 hours. (
Effects of amentoflavone on E7-mediated COX-2/IL-32 activation in SiHa and CaSki cells
RNA interference against HPV-16 E7 oncogene leads to viral E6 and E7 suppression in cervical cancer cells and apoptosis via up-regulation of Rb and p53. 19 Theoretically, the small interfering RNA targeting the E7 coding region can inhibit the expression of both E6 and E7 proteins simultaneously. 20 Based on the above-mentioned reports, we investigated whether amentoflavone would regulate E7 oncogene expression and performed RT-PCR using specific primers set for HPV-16 E7. As shown in Figure 3A, E7 transcript (300 bp) was down-regulated by amentoflavone in a dose-dependent manner. The inhibitory effect of amentoflavone on E7 expression was more evident in CaSki cells, which harbor more copies of HPV-16 type genome than SiHa cells. Because E7 oncogene is able to up-regulate COX-2, 21 we examined whether COX-2 could also be suppressed by amentoflavone-induced E7 down-regulation. As shown in Figure 3, both mRNA and protein expressions of COX-2 were significantly inhibited by amentoflavone. Additionally, we investigated whether IL-32 would be regulated by E7/COX-2 down-regulation. Recent reports showed that IL-32 depends on prior COX-2 activation 22 and E7-mediated COX-2 activation. 23 Our result also indicated that IL-32 was suppressed following by E7/COX-2 down-regulation in amentoflavone-treated SiHa and CaSki cells. These results suggest that amentoflavone induces apoptosis in SiHa and CaSki cervical cancer cells via blocking the E7-mediated COX-2/IL-32 activation.

Inhibition of human papillomavirus (HPV) E7-mediated cyclooxygenase-2 (COX-2)/interleukin-32 (IL-32) activation by amentoflavone in SiHa and CaSki cells. The cells were treated with different concentrations for 24 hours. (
Effect of amentoflavone as a PPARγ activator on an Akt-mediated signal pathway
To ascertain whether PPARγ is a target protein of amentoflavone, we analyzed the expression level of PPARγ by real-time qPCR and western blot. As shown in Figure 4, PPARγ expression was significantly increased at both transcriptional and post-transcriptional levels by amentoflavone treatment. Based on the previous report that the activation of PPARγ is associated with PTEN, which is a tumor suppressor and an upstream regulator of Akt, 24 we investigated the effects of amentoflavone on modulation of PTEN and found that the PTEN level was elevated in amentoflavone-treated SiHa and CaSki cells, thereby resulting in reduction of phosphorylated Akt content (Fig. 4B). These results suggest that amentoflavone might be a PPARγ activator and that the inhibition of Akt-mediated signaling is partially involved in the apoptosis induced by amentoflavone treatment in SiHa and CaSki cells.
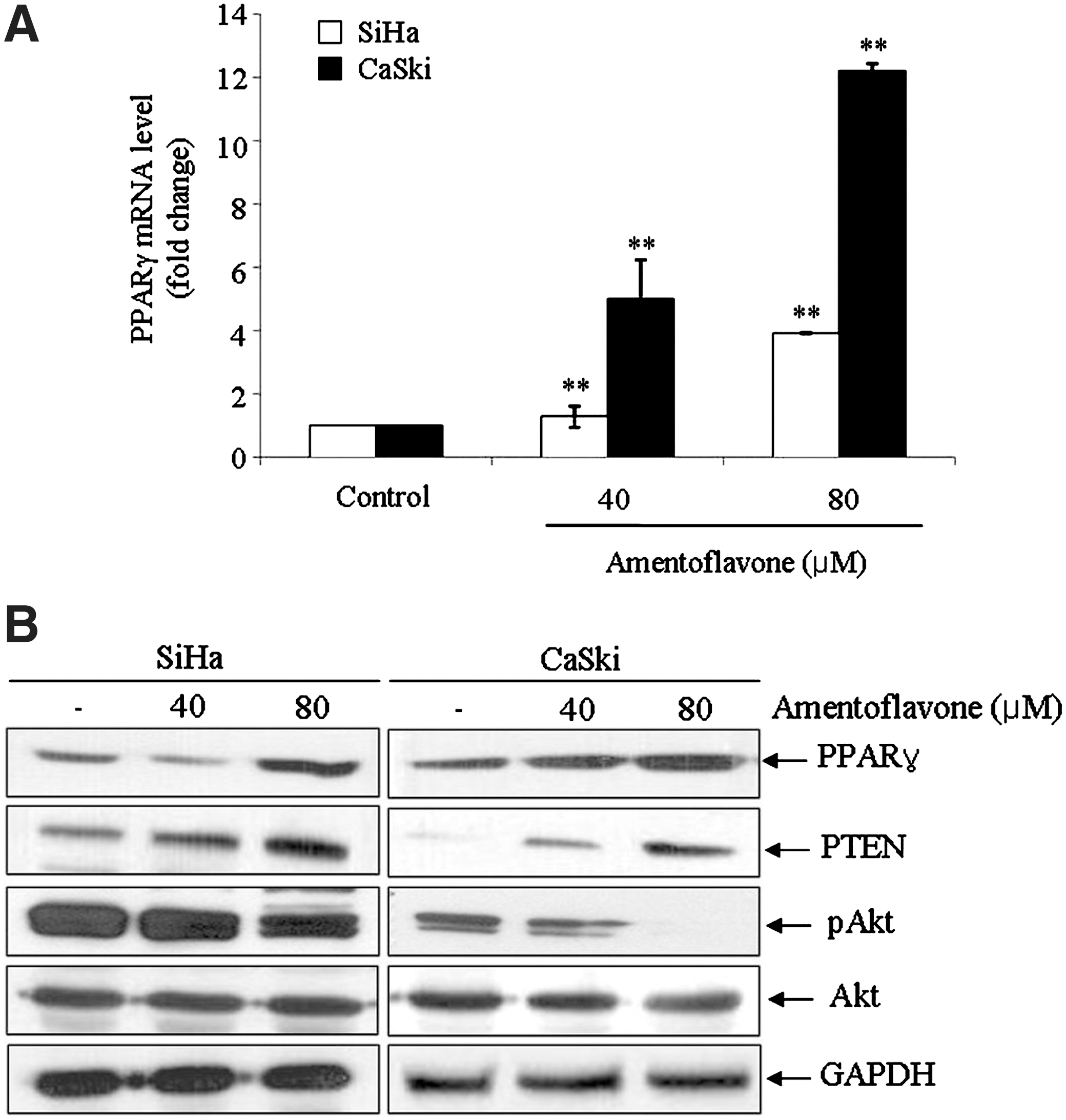
Effects of amentoflavone on expression of peroxisome proliferator-activated receptor γ (PPARγ) and on regulation of Akt-mediated signaling. SiHa and CaSki cells were treated with different concentrations of amentoflavone for 24 hours. (
Effects of amentoflavone on apoptotic signaling pathways
The activation of NF-κB plays a critical role in cancer cell survival. In addition, cysteine-aspartic proteases (caspases) and PARP are critical factors of apoptosis. As shown in Figure 5A, translocation of NF-κB p50 into the nucleus was significantly decreased by amentoflavone treatment, and cleaved PARP (89 kDa) was detected in amentoflavone-treated SiHa and CaSki cells. In particular, PARP was almost completely cleaved when CaSki cells were exposed to a high concentration of amentoflavone. We then examined whether the proteolytic cleavage of PARP would be induced by activation of caspases. The levels of cleaved caspase-9 (35 kDa and 37 kDa) were readily increased in a dose-dependent manner, whereas cleaved caspase-3 (17 kDa and 19 kDa) was detectable at high concentrations of amentoflavone. However, amentoflavone had no effect on caspase-8 activation (data not shown). Considering that amentoflavone-induced apoptosis may be via the intrinsic pathway involving activation of mitochondria, we subsequently attempted to evaluate the modulation of Bax/Bcl-2 proteins and cytochrome c release in amentoflavone-induced apoptosis by western blotting. The level of pro-apoptotic Bax was elevated in a dose-dependent manner, whereas the level of pro-survival Bcl-2 level was reduced, and then the release of cytochrome c into the cytosol was increased by amentoflavone treatment (Fig. 5B). Expression levels of extrinsic death receptors such as Fas (CD95) were not altered, and there was no cleavage of the downstream effector molecule caspase-8 by amentoflavone treatment (data not shown), suggesting that the Fas extrinsic death receptor would not be involved. These results suggest that amentoflavone induces apoptosis through intrinsic pathway accompanying the processing of caspase-9/-3 and PARP and the activation of the mitochondria in SiHa and CaSki cells.

Effects of amentoflavone on the processing of poly(ADP-ribose) polymerase (PARP) and caspases and on the expressions of mitochondrial-related apoptotic proteins in SiHa and CaSki cells. The cells were treated with different concentrations of amentoflavone for 24 hours. (
Discussion
Previous studies have shown that amentoflavone is associated with apoptosis, metastasis, and angiogenesis. 25,26 Therefore, amentoflavone may be considered to act as an antitumor agent. In this study, we demonstrated that amentoflavone exhibited anticancer activity against human cervical cancer cells. Several studies have reported apoptotic effects of amentoflavone. Amentoflavone induced apoptosis and antiproliferation via the inhibition of fatty acid synthase in human breast cancer cells. 27 Also, it produced apoptosis in B16F-10 melanoma cells by regulating the bcl-2, p53, and caspase-3 genes as well as those for nitric oxide and a pro-inflammatory cytokine. 28 However, the precise molecular mechanism has not been studied in amentoflavone-induced apoptosis in human cervical cancer cells. Our data showed that amentoflavone clearly inhibits the growth of SiHa and CaSki cells within 24 hours and induces the formation of apoptotic bodies. In the antiproliferation and apoptosis phenomena exhibited by amentoflavone in SiHa and CaSki cells, cell cycle arrest happened at the sub-G1 phase. Tumor suppressor protein p53 controls the G1 cell cycle checkpoints while promoting the induction of p21 and p27, which are cyclin-dependent kinase inhibitors, thereby regulating cell cycle–associated molecules such as G1 cyclins and pRb. 29 Our data demonstrated that cyclin E, cyclin A, and p-pRb were down-regulated, whereas p53, p21, and p27 were up-regulated, by amentoflavone treatment. The modulation of p-pRb is especially connected to a viral protein, HPV-E7, that binds to Rb and interferes with binding to E2F. 7 Several reports point out that the viral oncogene product E7 is related to the modulation of cellular proliferation and apoptosis. 30 Our results indicated that amentoflavone suppresses the expression of E7 oncogene in HPV-positive SiHa and CaSki cells, supporting the down-regulation of p-pRb in amentoflavone-induced cell cycle arrest. Previous reports showed that HPV-E7 oncogene regulates COX-2, which is overexpressed in cervical cancer cells and has been implicated in carcinogenic processes including cellular invasion, angiogenesis, and anti-apoptosis. 31,32 It is interesting that this current study supported our previous report indicating that E7 oncogene activates the expression of COX-2 and IL-32 and results in cancer development. 23 Our findings demonstrated that the inhibitory pathway of E7-mediated COX-2/IL-32 expression is involved in amentoflavone-induced apoptosis in SiHa and CaSki cells.
PPARγ regulates the proliferation, apoptosis, and differentiation in various human cancer cells, including lung, breast, colon, and prostate cancer cells. 13 Also, it has been known that activation of PPARγ induces the up-regulation of PTEN, which is a tumor suppressor protein, and subsequently inhibits Akt-mediated signaling, thereby resulting in apoptosis of cancer cells. 33 –35 Thus, PPARγ is a well-known target protein for anticancer therapy. Recent reports showed that amentoflavone can activate PPARγ in tumor necrosis factor-α–activated A549 cells. 36 Likewise, our results indicated that amentoflavone up-regulated the expression level of PPARγ in SiHa and CaSki cells and that the PTEN level was elevated and Akt phosphorylation was reduced in the amentoflavone-induced apoptotic process, suggesting that amentoflavone may be a PPARγ activator. Additionally, our data showed that the expression level of the anti-apoptotic factor Bcl-2 was decreased, whereas that of the well-known pro-apoptotic factor Bax was increased, thereby releasing cytochrome c into cytosol. Furthermore, amentoflavone treatment led to the activation of caspases-3/-9 and proteolytic cleavage of PARP in SiHa and CaSki cells. The expression level of an extrinsic death receptor such as Fas (CD95) was not altered by amentoflavone treatment, and amentoflavone did not cause the activation of the downstream effector molecule caspase-8 (data not shown). These findings suggest that amentoflavone induces apoptosis via suppressing E7 expression and mitochondria-emanated intrinsic pathways, which are known to be regulated by mitochondrial-related proteins composed of anti-apoptotic and pro-apoptotic members. 37 These results imply that amentoflavone may be developed as a therapeutic agent for human cervical cancer (Fig. 6).

Molecular mechanism by which amentoflavone induces apoptosis via suppressing E7 expression and in mitochondria-emanated intrinsic pathways in human cervical cancer cells. HPV E7 oncogene expression is suppressed by amentoflavone, followed by the inhibition of COX-2 and IL-32 expressions. Amentoflavone also activates PPARγ, and the PTEN level is increased, thereby resulting in inhibition of the Akt-mediated pathway. Moreover, amentoflavone triggers the intrinsic pathway following apoptosis via mitochondrial activation, inducing the growth inhibition of SiHa and CaSki cells and the ensuing sub-G1 cell cycle arrest. When these findings are taken together, the biflavonoid amentoflavone activates PPARγ/PTEN expressions and induces apoptosis via suppressing E7 expression, cell cycle arrest at the sub-G1 phase, and mitochondria-emanated intrinsic pathways in SiHa and CaSki human cervical cancer cells. Cyt c, cytochrome c; PI3K, phosphoinositide 3-kinase.
Footnotes
Acknowledgments
This work was supported by Korea Research Foundation grants (MEST 2010-0019306, 2009-0072028) funded by the Korean Government. D.-Y.Y is partially supported from the Priority Research Centers Program (2009-0093824) and a grant (0920080) from the National R&D Program for Cancer Control, Ministry for Health, Welfare and Family Affairs, Republic of Korea. J.H. and Y.Y are partially supported by grants R13-2008-001-00000-00 and 2009-0085906, respectively, through the Korea Research Foundation.
Author Disclosure Statement
No competing financial interests exist.