
Abstract
The purpose of the current study was to determine the anti-obesity and anti-inflammatory effects of an extract of purple sweet potatoes (PSPs) on 3T3-L1 adipocytes. For this purpose, differentiated 3T3-L1 adipocytes were treated with a PSP extract at concentrations of 1,000, 2,000, and 3,000 μg/mL for 24 hours. Then, we measured the changes in the sizes of the adipocytes, the secretion of leptin, and the mRNA/protein expression of lipogenic, inflammatory, and lipolytic factors after the treatment with the PSP extract. The PSP extract diminished leptin secretion, indicating that growth of fat droplets was suppressed. The extract also suppressed the expression of mRNAs of lipogenic and inflammatory factors and promoted lipolytic action. The antioxidative activity of the PSP extract was also measured using three different in vitro methods: 1,1-diphenyl-2-picrylhydrazyl free radical scavenging activity, ferric reducing ability potential assay, and chelating activity of transition metal ions. Taken together, our study shows that PSP extract has antilipogenic, anti-inflammatory, and lipolytic effects on adipocytes and has radical scavenging and reducing activity.
Introduction
O
Purple sweet potatoes (PSPs) are grown and developed in many countries, including Japan, to meet the growing demand for functional foods. PSPs are also used in various processed commercial products, including natural food colorants, juices, bread, noodles, jams, confectionaries, and fermented beverages. PSPs have an attractive reddish–purple color and contain high levels of anthocyanins and other phenolic compounds with known antioxidant activities. 1,2 Recent studies have demonstrated that anthocyanins from PSP extracts exhibit strong free radical scavenging activity, had antimutagenic activity, and significantly alleviated hypertension and liver injury in rats. 3 –6 Other physiological functions attributed to anthocyanins include anti-inflammatory activity, antimicrobial activity, protection from ultraviolet light, a reduction in memory impairment, and protection of the liver against oxidative stress. 7,8 The main PSP anthocyanins are acylated cyanidin and peonidin, 9 and these anthocyanins were reported to represent 1% and 11% of the total anthocyanins in PSPs, respectively. 10 In spite of acylated cyanidin's relatively low concentration in PSPs, an animal experiment revealed that it has a preventive effect against obesity and improves the hyperglycemia. 11 In particular, anthocyanins from black soybean were reported to reverse the effects of high-fat diet on body weight and serum lipids in an in vivo experiment. 12
However, reports of anti-obesity and anti-differentiation effects of PSPs have been scarce. The current study examined the anti-obesity effects of a PSP extract, specifically its effect on lipogenetic, lipolytic, and inflammatory mechanisms, which have not been extensively examined with PSP previously. Additionally, we evaluated the antioxidant effects of PSP extract using three in vitro methods based on different mechanisms: 1,1-diphenyl-2-picrylhydrazyl (DPPH) scavenging activity, ferric reducing ability potential (FRAP) assay, and Fe2+-chelating activity. 2,3,13
Materials and Methods
Materials
PSPs were provided by Dongwon Dairy Food (Suwon, Gyeonggi-do, Korea). High-glucose Dulbecco's modified Eagle's medium (DMEM), bovine calf serum, and fetal bovine serum were purchased from Hyclone (Logan, UT, USA). DPPH, FeCl2·4H2O, FeCl3·6H2O, 3-2-pyridyl-5,6-diphenyl-1,2,4-triazine-4′,4″-disulfonic acid sodium salt (ferrozine), HCl, 2,4,6-tris-2-pyridyl-s-triazine chloride, trypsin/EDTA, insulin, dexamethasone, 3-isobutyl-1-methylxanthine, and methyl thiazol tetrazolium (MTT), which were used in cell culture and for sample treatments, were purchased from Sigma (St. Louis, MO, USA). Phosphate-buffered saline (PBS) was purchased from Lonza (Walkersville, MD, USA). easyBLUE for RNA extraction and PRO-PREP for protein extraction were purchased from iNtRON Biotechnology (Seongnam, Gyeonggi-do, Korea). ImProm reverse transcriptase was purchased from ProMega (Madison, WI, USA). HotStart-IT® SYBR® Green qPCR Master Mix was purchased from USB Corp. (Cleveland, OH, USA). All primers and oligo(dT) primers were purchased from Bioneer (Daejeon, Korea). Antibodies against β-actin and phospho–hormone-sensitive lipase (HSL) were purchased from Cell Signaling (Danvers, MA, USA).
Extraction of crude PSP anthocyanin and toxicity measurement
Fifty grams of peeled PSPs was chopped and soaked in 1 L of 20% ethanol (with 0.1% citric acid added) and extracted by the method of Kim et al., 14 with some modifications. Filtration was followed by evaporation of the liquid and then by lyophilization. The extract had a yield of 7.8%.
Toxicity of the PSP extract was measured using an MTT assay. Cells were seeded at 104 cells per well in 96-well plates and incubated in culture medium. Then, the cells were treated with designated concentrations of the PSP extract, followed by a 24-hour incubation. After the treatment, MTT was added to the cells and incubated for 4 hours to observe formazan, which reflects the activity of living cells via mitochondrial dehydrogenase activity that reduces MTT to purple formazan. The formazan was solubilized with dimethyl sulfoxide, and its absorbance at 595 nm was measured.
Measurement of cyanidin concentration
Lyophilized PSP extract was dissolved in deionized distilled water to make a 1,000 mg/mL solution, and the concentration of cyanidin-3-glycoside was measured using a pH-differential method. 15 The appropriate dilution factor for the sample was determined with a spectrophotometer by diluting with a potassium chloride buffer (pH 1.0) until the absorbance of the sample at 510 nm reached an optical density of 1.2. Two dilutions of the sample, one with a potassium chloride buffer (pH 1.0) and the other with a sodium acetate buffer (pH 4.5), were each diluted by a predetermined dilution factor (×300). The absorbance of each dilution was measured at 510 and 700 nm (to correct for haze). One gram of the extract contained 4.28 mg of cyanidin.
3T3-L1 adipocyte culture
Mouse 3T3-L1 pre-adipocytes were grown to confluence in DMEM with 10% bovine calf serum at 37°C in a humidified atmosphere of 5% CO2. At day 1 post-confluency (designated “day 0”), cell differentiation was induced with a mixture of 3-methyl-1-isobutylxanthine (0.5 mM), dexamethasone (1.25 μM), and insulin (4 μg/mL) in DMEM containing 10% fetal bovine serum. On day 2 and thereafter, DMEM containing only 10% fetal bovine serum and insulin (4 μg/mL) was subsequently replaced every 2 days.
The PSP extract was dissolved in PBS, divided into small quantities, and stored at −20°C. At day 8, after induction of differentiation of the adipocytes, the extract was dissolved in DMEM at different concentrations (control, 1,000, 2,000, or 3,000 μg/mL) and added to the cultured adipocytes for 24 hours.
Oil Red O staining and size measurement of adipocytes
Accumulation of TG was measured by Oil Red O staining. Cultured adipocytes were washed with PBS three times prior to fixation with 3.8% formaldehyde (Sigma) for at least 4 hours. They were rinsed again with PBS three times and then treated with filtered Oil Red O staining solution (Sigma) for 1 hour. Stained cells were observed with a microscope, and the dye was then eluted with isopropanol. The eluted dye was collected, and the optical density at 540 nm was measured. 16 During the fixation of the adipocytes with formaldehyde, photographs were taken to measure cell size. The AlphaImager™ 2200 Series program (Alpha Innotech, San Leandro, CA, USA) was used to measure the sizes from the images of the adipocytes.
Leptin enzyme-linked immunosorbent assay
Concentrations of leptin secreted by the 3T3-L1 adipocytes were determined using an enzyme-linked immunosorbent assay from R&D Systems (Minneapolis, MN, USA), with some modifications. The absolute concentration of leptin was calculated using a standard leptin calibration curve.
RNA extraction, cDNA synthesis, and real-time polymerase chain reaction
Total RNA isolation, first-strand cDNA synthesis, and real-time polymerase chain reaction analysis were performed using the method of Do et al. 17 with modifications. For real-time polymerase chain reaction analysis, samples were incubated in the polymerase chain reaction system for an initial denaturation at 95°C for 2 minutes, followed by 50 polymerase chain reaction cycles; each cycle consisted of 95°C for 1 second and 60°C for 1 minute. To confirm amplification of specific transcripts, melting curve profiles (phase 1, slowly heating the sample to 95°C for 15 seconds and cooling to 60°C for 1 minute; phase 2, heating the sample to 95°C for 30 seconds and cooling to 60°C for 15 seconds, with continuous fluorescence measurement during both phases) were produced at the end of each polymerase chain reaction. The threshold cycles (C t) for the target genes, in comparison with the cycles of β-actin, were determined in triplicate experiments, and the relative RNA quantity was calculated by the comparative C t method. The oligonucleotide primers for the experiment are indicated in Table 1.
ACD, acyl-coenzyme A dehydrogenase; ACO, acyl-coenzyme A oxidase; ACS, acetyl-coenzyme A synthetase; COX-2, cyclooxygenase 2; CPT-1, carnitine palmitoyltransferase 1; FAS, fatty acid synthetase; HSL, hormone-sensitive lipase; IL-6, interleukin-6; LPL, lipoprotein lipase; MCP1, monocyte chemotactic protein 1; SREBP-1c, sterol regulatory element binding protein-1.
Western blot analysis
After experimental treatment, cells were washed twice with ice-cold PBS and lysed with an extraction buffer (PRO-PREP). Insoluble materials were removed by centrifugation at 15,000 g for 10 minutes at 4°C. The total concentration of extracted proteins was determined using the method of Bradford. 18 The proteins in the supernatants were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes. After blocking with 10 mM Tris, 150 mM NaCl, and 0.05% Tween-20 (pH 7.6) containing 5% nonfat milk for 1 hour at room temperature, the membranes were incubated with the appropriate primary antibodies. To detect the antigen-bound antibodies, the blots were treated with secondary antibodies conjugated with horseradish peroxidase. Immunoreactivity was detected using an enhanced chemiluminescence western blotting detection system.
DPPH radical scavenging activity
The free radical scavenging activity of PSPs was analyzed using the DPPH method. 19 Two hundred microliters of PSP extract and 800 μL of DPPH solution (200 μM in methanol) were mixed, and the absorbance was measured at 517 nm for 30 minutes at room temperature using a spectrophotometer. Ascorbic acid was used as a positive control.
DPPH radical scavenging activity was calculated using the following equation: DPPH radical scavenging activity (%)=(1−[absorbance of sample at 517 nm/absorbance of control at 517 nm])×100.
FRAP assay
The ferric reducing ability was measured by the modified method of Benzie and Strain. 20 The FRAP solution contained 25 mL of 0.2 M acetate buffer (pH 3.6), 2.5 mL of 10 mM 2,4,6-tris-2-pyridyl-s-triazine chloride in 40 mM HCl, and 2.5 mL of 20 mM FeCl3·6H2O. For the reactions, 100 μL of diluted PSP extracts was added to 900 μL of FRAP solution. Absorbance of the mixture was measured at 593 nm.
Fe2+-chelating activity
The Fe2+-chelating activity of the PSP extract was measured using a method modified from that of Ozsoy et al. 21 Five hundred microliters of PSP extracts was mixed with 50 μL of 2 mM FeCl2·4H2O and 1.85 mL of methanol. The reaction was started by adding 150 μL of 5 mM ferrozine, and the mixture was incubated at room temperature for 10 minutes. The absorbance was measured at 562 nm and compared with that of the positive control (EDTA).
Statistical analysis
The data were analyzed with Student's t test and/or analysis of variance test at the P<.05 level.
Results
PSP extract altered sizes of adipocytes and suppressed levels of TG and leptin
3T3-L1 adipocytes were treated with a range of concentrations of the PSP extract (0, 10, 100, 1,000, 2,000, 3,000, and 5,000 (μg/mL), none of which had any toxic effects (Fig. 1a). To show the anti-obesity effect of the PSP extract on differentiated 3T3-L1 cells, we measured the size of PSP-treated adipocytes. Microscopic images of adipocytes were taken during fixation with formaldehyde for Oil Red O staining. Two hundred adipocytes were randomly chosen from the image files and analyzed with AlphaImager software. The average cell size of adipocytes treated with PSP extract was decreased compared with that of the control (Fig. 1b). Microscopic images of adipocytes taken during Oil Red O staining showed that oil droplets in adipocytes were smaller compared with control cells (Fig. 1c). TG accumulation was lower with PSP extract treatment compared with the control (92.6% with 3,000 μg/mL PSP extract, P<.01) (Fig. 1d).

Effects of a purple sweet potato (PSP) extract on (
Leptin levels were significantly lower following PSP extract treatment compared with the control in a dosage-dependent manner: 76.7% (1,000 μg/mL), 71.3% (2,000 μg/mL), and 62.4% (3,000 μg/mL) of control (Fig. 1e).
PSP extract suppressed the expression of adipogenic enzymes and a transcription factor
In this study, we measured the changes in the expression of a transcription factor and adipogenic enzymes by PSP extract treatment. The PSP extract dose-dependently suppressed the expression level of acetyl-coenzyme A (CoA) synthetase by up to 34.8% (Fig. 2a). Expression of fatty acid synthetase was also diminished by the PSP extract in a dose-dependent manner; with expression as low as 11.3% of control at 3,000 μg/mL (Fig. 2b). Expression of lipoprotein lipase was also significantly reduced up to 52.9% that of the control at 3,000 μg/mL (Fig. 2c). The expression of sterol regulatory element binding protein-1c was significantly and dose-dependently down-regulated by the PSP extract to as low as 16.7% of the control with 3,000 μg/mL PSP extract (Fig. 3).
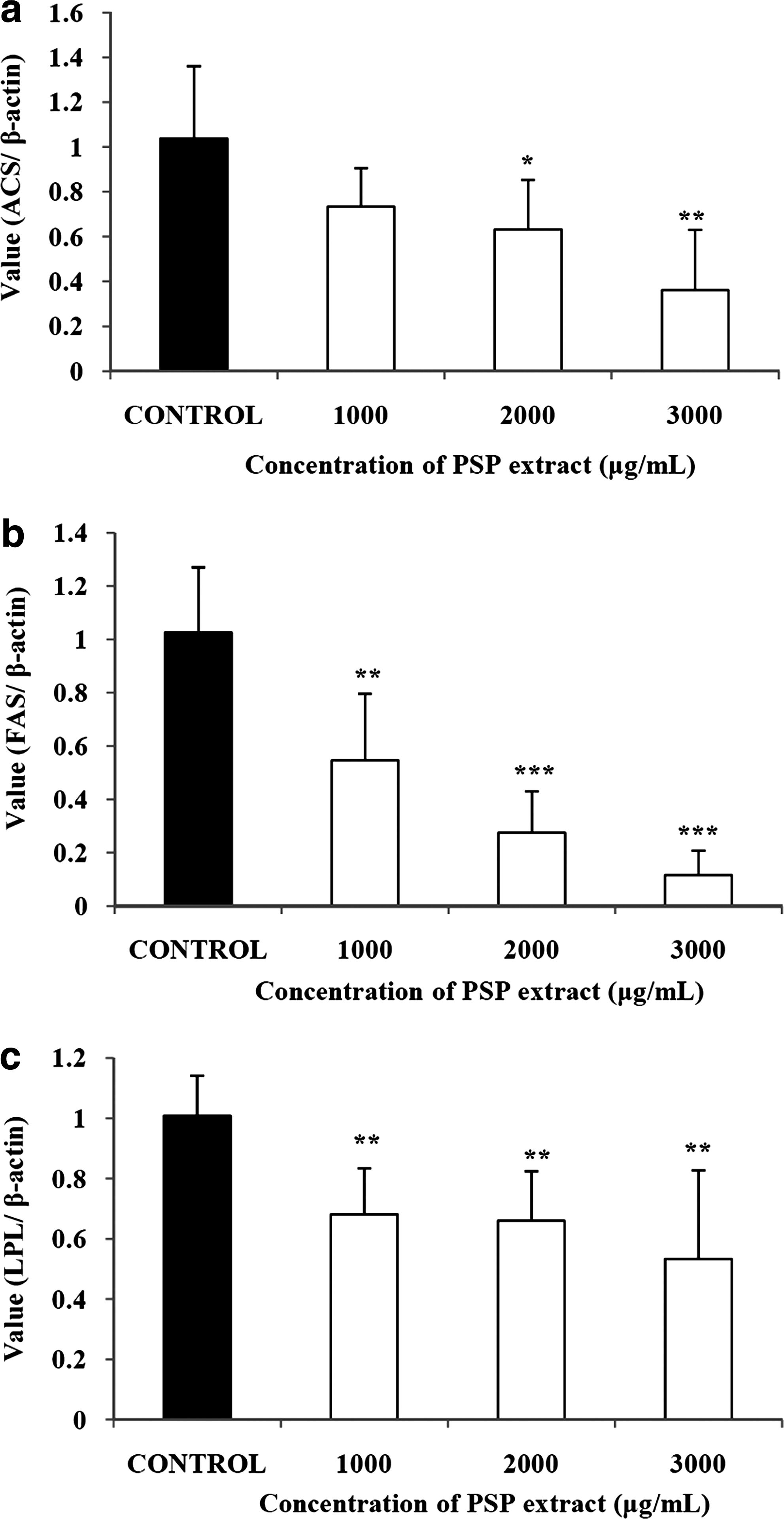
Effects of a PSP extract on (
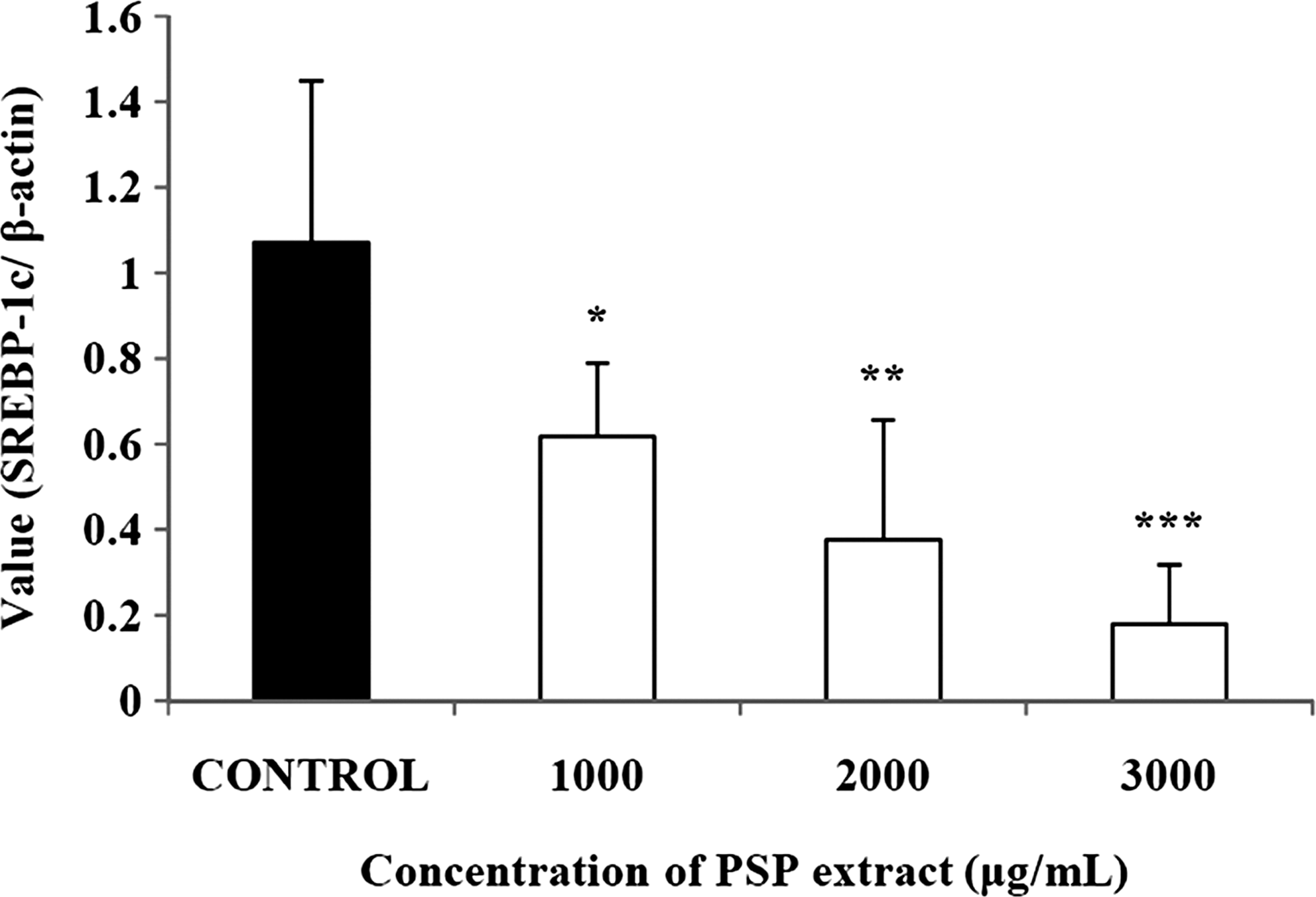
Effects of a PSP extract on SREBP-1c in 3T3-L1 adipocytes. *P<.05, **P<.01, ***P<.001 compared with control (expressed by Student's t-test). Data are mean±SD values from triplicate experiments.
PSP extract induced lipolytic factors
The expressions of HSL and perilipin were measured to determine the induction of lipolytic factors. Compared with the control, the mRNA expression level of perilipin was up-regulated by up to 144.2% of the control in a dose-dependent manner (Fig. 4a). In addition, the mRNA expression level of HSL was also significantly altered on treatment with the PSP extract by up to 129.7% (Fig. 4b) of the control. We also measured the change in the expression level of phosphorylated HSL levels relative to that of unphosphorylated HSL induced by the PSP extract, and the expression was up-regulated in a dose-dependent manner (Fig. 4c). Protein expression of perilipin was generally up-regulated, even though such regulation was less significant (Fig. 4d).
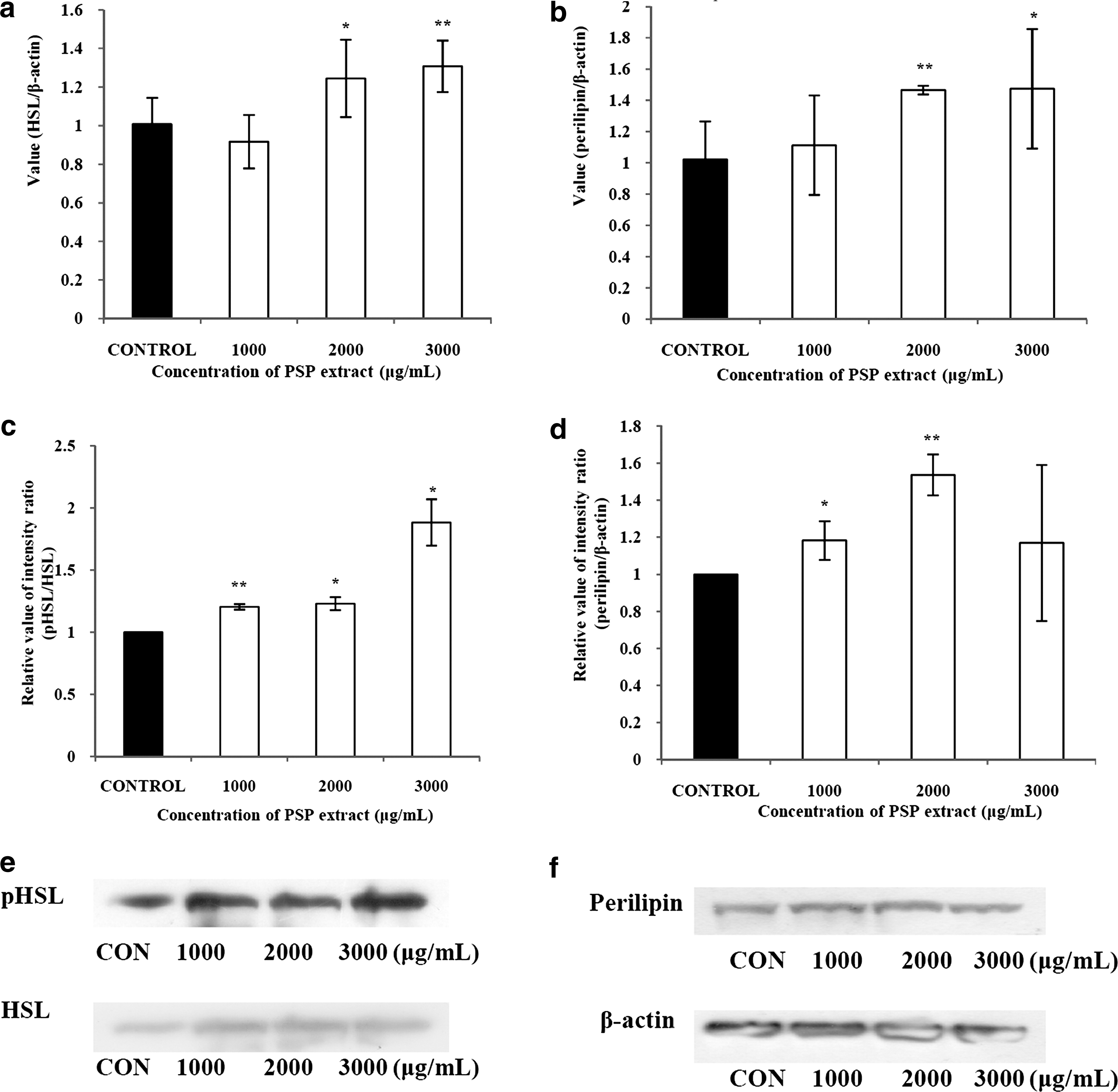
Effects of a PSP extract on mRNA expression of (
PSP extract promoted the expression of β-oxidation factors
We measured the changes in the expression of factors closely related to β-oxidation. The PSP extract significantly promoted the expression of acyl-CoA oxidase by up to 176% (Fig. 5a). Carnitine palmitoyltransferase-1 was also up-regulated by treatment with the extract by up to 194% (Fig. 5b). Furthermore, expression levels of acyl-CoA dehydrogenase were significantly increased by up to 193% (Fig. 5c).
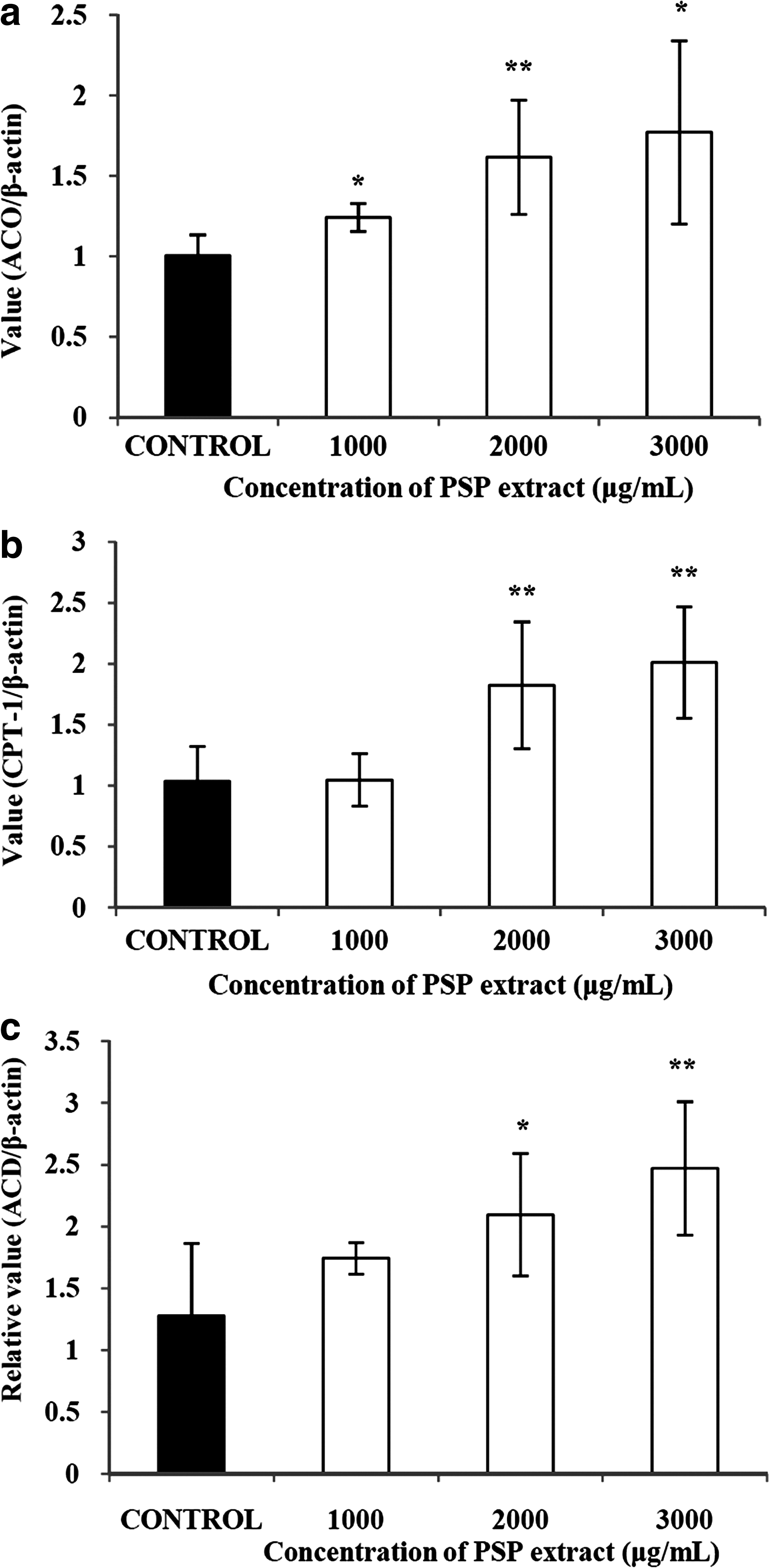
Effects of a PSP extract on (
PSP extract inhibited the expression of inflammatory factors
We also examined the effect of the PSP extract on the expression of inflammatory factors. The PSP extract significantly suppressed the expression of cyclooxygenase 2 by up to 20.1% (Fig. 6a). The mRNA expression of interleukin-6 was also significantly lowered by the PSP extract to 55.0% of the control with the PSP extract at 3,000 μg/mL (Fig. 6b). The PSP extract also significantly suppressed the expression of haptoglobin at all concentrations tested (P<.001) (Fig. 6c). The expression of monocyte chemotactic protein-1, even though it was less proportional to the dose of the PSP extract, was also significantly diminished by the PSP extract (Fig. 6d).
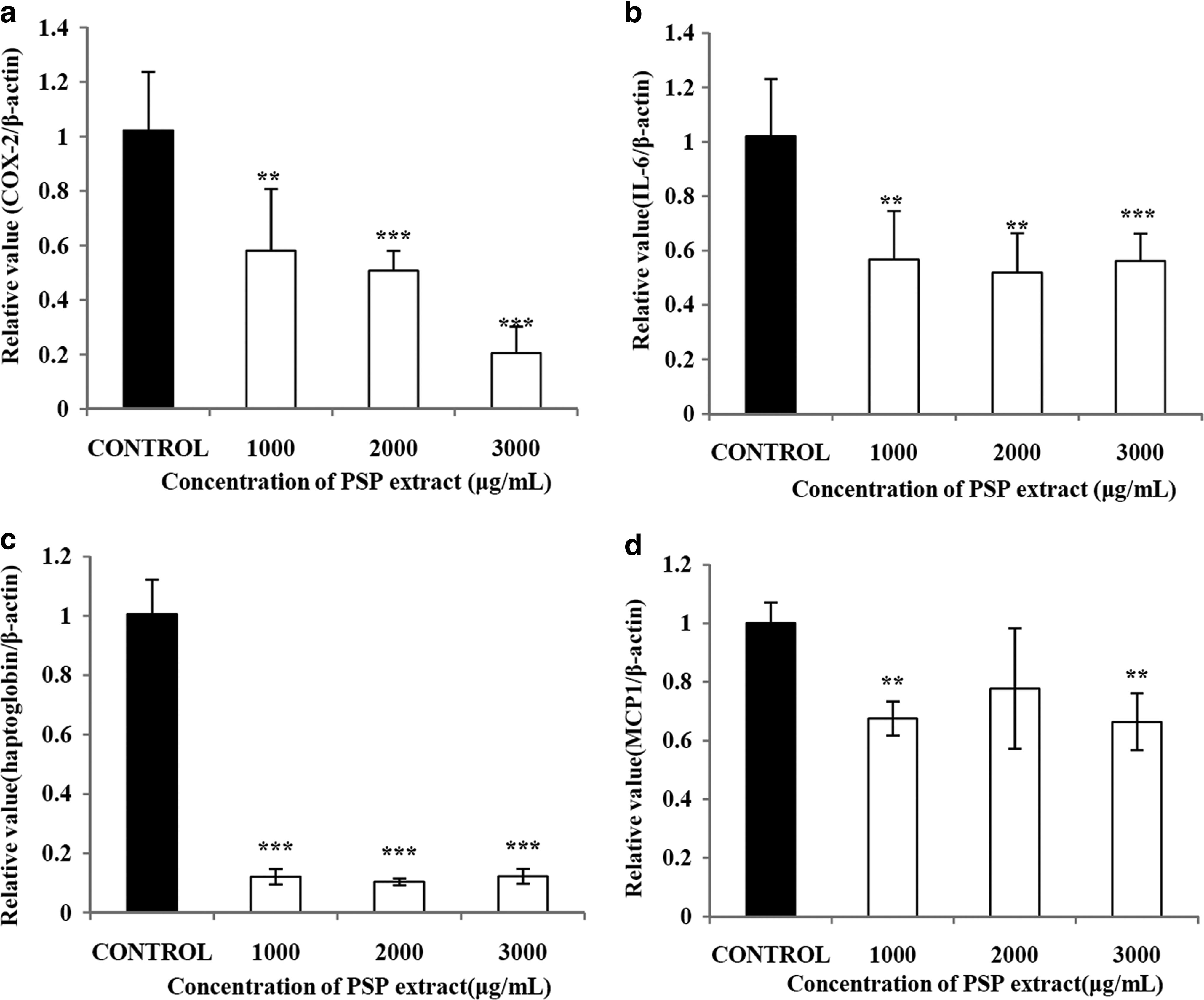
Effects of a PSP extract on (
Antioxidant effects of PSP extract
We examined the antioxidant effects of the PSP extract using three different methods in vitro: DPPH free radical scavenging activity assay, FRAP assay, and chelating activity assay of transition metal ions. The results of the DPPH and FRAP experiments were expressed in ascorbic acid equivalents (Table 2). In the DPPH method, the ascorbic acid equivalent values of the PSP extract were 559.46 and 844.77 μM with 2 and 4 mg/mL concentrations, respectively. The FRAP assay also exhibited similar values of DPPH activity. This indicated that the extract exhibited significant dose-dependent antioxidative capactiy. However, the Fe2+-chelating activity of the PSP extract was not changed by concentrations ranging from 0.1 mg/mL to 2 mg/mL (Fig. 7). In contrast, EDTA, the positive control in this assay system, showed a high chelating ability of about 99% at concentrations as low as 0.1 mg/mL (data not shown).
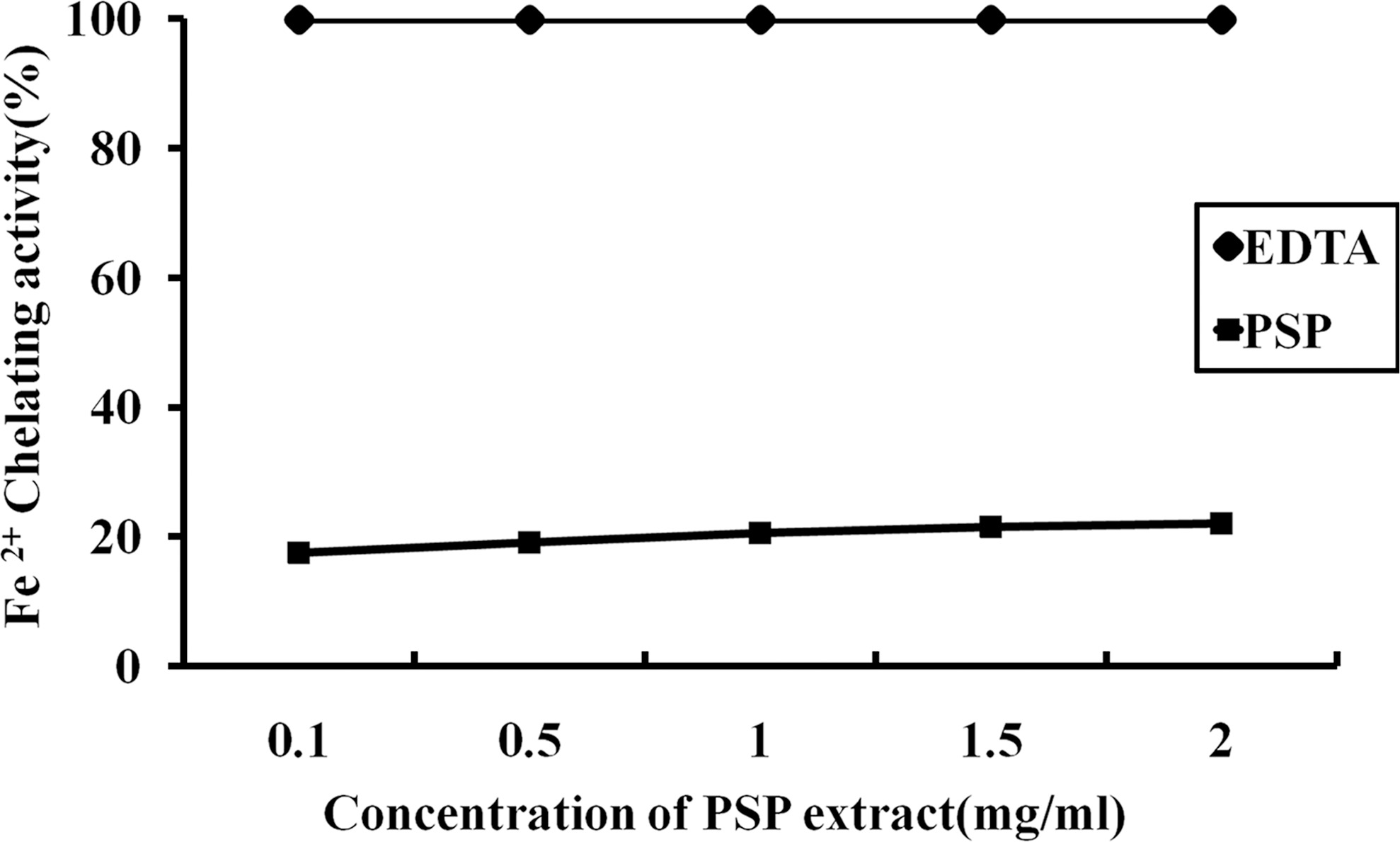
Fe2+-chelating activity of the PSP extract. EDTA was used as a positive control. Data are mean±SD values from triplicate experiments.
Data are mean±SD values (n=3).
Means with the same letters are significantly different (P<.05): 1,1-diphenyl-2-picrylhydrazyl (DPPH), P=1.57×10−4; ferric reducing ability potential (FRAP), P=5.72×10−16.
Discussion
This study elucidated the anti-obesity potential of a PSP extract that suppressed the expression of lipogenic factors including fatty acid synthetase, lipoprotein lipase, and acetyl-CoA synthetase as well as the transcription factor sterol regulatory element binding protein-1c and up-regulated lipolytic enzymes such as phospho-HSL. In addition, this study showed that the PSP extract suppresses the expression of inflammatory factors interleukin-6, haptoglobin, and cyclooxygenase 2. In addition, we found that the PSP extract produced antioxidative actions through the ferric chelation test and the DPPH assay.
The size of adipocytes is a major modulator of their endocrine function. Hypertrophic adipocytes, for instance, secrete greater amounts of free fatty acids than normal-sized adipocytes. 22 Treatment with the PSP extract significantly reduced the size of adipocytes, which suggests that such a treatment may diminish the accumulation of free fatty acids in adipocytes.
Leptin is a cytokine exclusively secreted by adipocytes in proportion to the amount of accumulated TG. 23 This study showed that treatment with the PSP extract significantly reduced leptin levels, and it can be inferred that the PSP extract may have at least suppressed de novo fatty acid synthesis, which may lead to decreased accumulation of TGs and eventually in the secretion of leptin.
Adipogenesis is induced by several enzymes, including fatty acid synthetase and acetyl-CoA synthetase. The latter catalyzes the formation of acetyl-CoA from acetate, coenzyme A, and ATP 24 so that acetyl-CoA can be fed into lipid synthesis or the tricarboxylic acid cycle. Fatty acid synthetase is a complex of multifunctional polypeptides that catalyzes the synthesis of long-chain saturated fatty acids, including palmitate, in the presence of acetyl-CoA as a primer and with malonyl-CoA as the two-carbon donor. 25 Lipoprotein lipase is an enzyme that hydrolyzes lipid in lipoproteins, such as very low-density lipoprotein or chylomicrons, into two free fatty acids and one monoacylglycerol to allow fatty acid entry into adipose tissue. 26 We observed a significant decrease in the expression of such factors upon treatment with the PSP extract, which strongly suggests that the PSP extract suppresses lipogenesis in 3T3-L1 cells.
Adipocytes undergo differentiation by the activation of transcription factors. Genes to encode enzymes such as fatty acid synthetase or acetyl-CoA are expressed under the control of transcription factors 27 such as sterol regulatory element binding protein-1c. Treatment with the PSP extract significantly decreased the expression of sterol regulatory element binding protein-1c in differentiated 3T3-L1 adipocytes in a dose-dependent manner, and the expressions of fatty acid synthetase, lipoprotein lipase, and acetyl-CoA synthetase were also significantly diminished in a similar manner.
When ATP is converted to cyclic AMP by adenylyl cyclase in response to a hormone such as glucagon, cyclic AMP activates protein kinase A to phosphorylate HSL and perilipin, which leads to the degradation of TGs to free fatty acids and glycerol. 28,29 In this experiment, we examined the effect of the extract on the mRNA expression of HSL and perilipin. Additionally, protein expression of phospho-HSL was also observed along with mRNA expression. Such treatment significantly and dose-dependently promoted the expression of these enzymes. We observed a significant up-regulation in the expression of such factors, showing that the PSP extract boosts the lipolytic action of 3T3-L1 cells in addition to its lipogenic effects.
Mitochondrial β-oxidation of fatty acids provides energy, especially during fasting conditions. Fatty acid oxidation occurs in the mitochondria and consists of a repeating circuit of four sequential steps. Carnitine palmitoyltransferase-1 is the rate-limiting enzyme for fatty acid oxidation. This enzyme catalyzes the formation of acylcarnitine from acyl-CoA, which is the first step in the transport of long-chain fatty acids from the cytosol into the mitochondrial matrix for fatty acid oxidation. Acyl-CoA oxidase catalyzes the first step of the β-oxidation cycle in which a fatty acyl-CoA is oxidized to 2-trans-enoyl-CoA. Acyl-CoA dehydrogenase, specifically medium-chain acyl-CoA dehydrogenase, is responsible for catalyzing the dehydrogenation of medium-chain length (C6–C12) fatty acid thioesters. 30,31 In this experiment, we examined the effect of the PSP extract on the mRNA expression of acyl-CoA oxidase, carnitine palmitoyltransferase-1, and acyl-CoA dehydrogenase. Treatment with the PSP extract significantly promoted the expression of these enzymes in a dose-dependent manner, showing that PSP extract promotes β-oxidation in 3T3-L1 adipocytes.
Lipogenesis is also correlated with inflammation. Adipocytes express certain inflammatory factors, including cyclooxygenase 2 and interleukin-6, in proportion to their size, which is associated with the accumulation of free fatty acids as previously described. Inflammatory adipokines such as interleukin-6 and cyclooxygenase 2 are closely connected to complications from obesity, including type 2 diabetes, hypertension, and arthrosclerosis. 31 Some natural materials such as berberin were found to suppress the expression of inflammatory factors such as interleukin-6, C-reactive protein, and haptoglobin. 17 In particular, anthocyanins are found to lessen expression of inflammatory factors, including monocyte chemotactic protein-1, not to mention that they also elucidate improvement in lipid parameters in vivo. 32 PSP extract decreased the mRNA expression of cyclooxygenase 2, monocyte chemotactic protein-1, haptoglobin, and interleukin-6, demonstrating a dose-dependent anti-inflammatory effect. Taken together, treatment with the PSP extract diminished the synthesis and accumulation of free fatty acids and up-regulated lipolysis, which may lead to a reduction in the secretion of inflammatory adipokines.
The PSP extract showed a significant antioxidative effect in two different assays, DPPH and FRAP, but the antioxidative properties could not be measured using a PSP extract concentrations of 5 mg/mL and higher because of color interference from the extract. Anthocyanins with ortho-dihydroxy groups can inhibit hydroxyl radical generation by chelating transition metals. 33,34 However, PSP had no Fe2+-chelating activity, suggesting that it is not a metal ion chelator. This result indicates the possibility that there are no ortho-dihydroxy groups in the anthocyanin molecules that were extracted from the PSPs and would explain the lack of Fe2+-chelating activities.
In conclusion, the PSP extract examined here had the capacity to suppress lipogenesis and inflammation and to induce lipolysis and β-oxidation. The PSP extract had an antioxidative effect of radical scavenging ability and reducing capacity but was not a metal ion chelator.
Footnotes
Acknowledgment
This study was supported by the Technology Development Program for Agriculture and Forestry, Ministry for Food, Agriculture, Forestry, and Fisheries, Republic of Korea (project number 109114-03-1-HD120).
Author Disclosure Statement
J.-O.Y. and M.-S.S. are employees of Dongwon Dairy Food. All other authors declare no competing financial interests exist.