
Abstract
The hepatoprotective effect of aged black garlic (ABG) against ethanol-induced oxidative liver damage was investigated in adult male Sprague–Dawley rats for 4 weeks. Rats were divided into three groups: a saline (WT) group, an ethanol (ET) group (15 mL/kg of body weight 20% [wt/vol] ethanol), and an ethanol + ABG (ET+ABG) group (ethanol + 100 mg/kg of body weight ABG). ABG administration led to decreased epididymal and total fat pad (P<.05) and liver weights, ameliorated prominent fatty changes around the portal triad, and reduced fat accumulation in liver. ABG caused a significant decrease of the alcohol-induced increases in hepatic activities of aspartate aminotransferase, alanine aminotransferase, alkaline phosphatase, and lactate dehydrogenase. Cytochrome P450 2E1 activity was reduced by 55%, whereas the activities of glutathione S-transferase and quinine reductase were increased by 1.5-fold (P<.05) and fourfold (P<.05), respectively, in the ET+ABG group compared with the ET group. ABG treatment significantly decreased the thiobarbituric acid–reactive substances level in liver, heart, and plasma. Glutathione content and the activities of antioxidant enzymes such as glutathione peroxidase, glutathione reductase, and catalase in liver were significantly enhanced. Furthermore, the oxidative damage to blood lymphocyte DNA caused by chronic alcohol ingestion was significantly decreased in the ET+ABG group. In conclusion, ABG has strong antioxidative properties and may be a promising agent for protecting against chronic alcohol-induced liver damage.
Introduction
G
Aged garlic extract is an odorless product obtained from prolonged extraction of fresh garlic at room temperature with highly available biological activity in vitro. Aged garlic extract contains water-soluble allyl amino acid derivatives, which account for most of its organosulfur content, stable lipid-soluble allyl sulfides, flavonoids, saponins, and essential macro- and micronutrients. 5 Aged black garlic (ABG) recently became available in the Korean market and is a new form of garlic preparation produced by aging garlic at high temperature and humidity. ABG, like other aged garlic products, is rich in bioactive organic sulfur compounds such as S-allylcysteine, S-allylmercaptocysteine, and tetrahydro-β-carboline. 6 It is also higher in superoxide dismutase–like activity, hydrogen peroxide (H2O2) scavenging activity, and polyphenols than normal garlic. 6 Therefore, ABG has stronger in vitro and in vivo antioxidant activity than raw garlic and may have greater efficacy for preventing metabolic diseases and alcoholic hepatotoxicity. 7,8
Alcohol-induced oxidative stress is caused by each of the three ethanol metabolic pathways involving alcohol dehydrogenase, the microsomal ethanol oxidation system, and catalase. 9 Alcohol is catalyzed by alcohol dehydrogenase to form acetaldehyde, which results in the formation of free radicals. In the mitochondria, microsomal ethanol oxidation via catalysis by the cytochrome P450 2E1 (CYP2E1) isoenzymes form reactive oxygen species (ROS) and large amounts of H2O2. 10 Peroxisomal activity also contributes to ethanol oxidation in the liver and might be more prominent in heavy ethanol consumers, who accumulate fatty acids in the liver because of increased fatty acid peroxisomal oxidation. The oxidative stresses caused by ethanol metabolism contribute to these hepatotoxic effects. 11
This study evaluated capacity of ABG to mitigate chronic alcohol-induced hepatotoxicity through antioxidant activity in rats.
Materials and Methods
Materials
Garlic was fermented in a thermo-hygrostat (model TH-500 [patent pending], Hanyoung nux Co., Incheon, Korea) for 9 days maintaining proper temperature and humidity. ABG was rendered black by fermentation. Glutathione reductase (GR), reduced glutathione, oxidized glutathione, tetramethoxypropane, 5,5′-dithiobis(2-nitrobenzoic acid), NADP, NADPH, thiobarbituric acid, and bovine serum albumin were products of Sigma-Aldrich Chemical Co. (St. Louis, MO, USA). All reagents used were of analytical grade.
Animals and experimental design
The animal experiment complied with the guidelines for care and use of laboratory animals of the U.S. National Institutes of Health. Four-week-old male Sprague–Dawley rats (Damul Science, Daejeon, Korea) were housed in individual polycarbonate cages in an animal room with controlled temperature (23±2°C) and humidity (55±10%) under a 12-hour/12-hour light–dark cycle with free access to a normal diet (AIN 93G, Samyang Co., Seoul, Korea) (Table 1) and water. To determine the proper dosage of ABG for rats, we performed a dose–response pretest in which commercial alcohol detoxication beverage (15 mL/kg of body weight) and ABG (0, 50, 100, or 400 mg/kg) were orally administered to rats, and then 20% ethanol (15 mL/kg) was given at 30-minute intervals. Plasma alcohol and acetaldehyde contents were measured up to 7 hours. Doses of ABG at 100 and 400 mg/kg of body weight resulted in significantly lower concentrations of plasma alcohol and acetaldehyde than in the control (0 mg/kg ABG), and no significant difference was found among treatment groups receiving the commercial beverage or 100 and 400 mg/kg ABG (data not shown). Therefore, we chose 100 mg/kg ABG as the dose for this study.
AIN 93G Mineral Mixture (in g/kg): calcium carbonate, 357.00; potassium phosphate (monobasic), 196.00; potassium citrate·H2O, 70.78; sodium chloride, 74.00; potassium sulfate, 46.60; magnesium oxide, 24.00; ferric citrate (USP), 6.06; zinc carbonate, 1.65; manganous carbonate, 0.63; cupric carbonate, 0.30; potassium iodate, 0.01; sodium selenate, 0.01025; ammonium paramolybdate·4H2O, 0.00795; sodium metasilicate·9H2O, 1.45; chromium potassium sulfate·12H2O, 0.275; lithium chloride, 0.0174; boric acid, 0.0815; sodium fluoride, 0.0635; nickel carbonate, 0.0318; ammonium vanadate, 0.0066; and sucrose (finely powdered), 221.026.
AIN 93G Vitamin Mixture (in g/kg): niacin, 3.00; calcium pantothenate, 1.60; pyridoxine HCl, 0.70; thiamine HCl, 0.60; riboflavin, 0.60; folic acid, 0.20; biotin, 0.02; vitamin E acetate (500 IU/g), 15.00; vitamin B12 (0.1%), 2.50; vitamin A palmitate (500,000 IU/g), 0.80; vitamin D3 (400,000 IU/g), 0.25; vitamin K1/dextrose mix (10 mg/g), 7.50; and sucrose, 967.23.
Seven-week-old male Sprague–Dawley rats were fed a normal diet for a 1-week acclimation period and then divided into three groups: saline (WT) group (15 mL/kg of body weight saline, n=5), ethanol (ET) group (15 mL/kg of body weight 20% [wt/vol] ethanol, n=7), and ethanol + ABG (ET+ABG) group (ethanol + 100 mg/kg of body weight ABG, n=7). The ET+ABG group was orally administered 100 mg/5 mL of distilled water/kg of body weight ABG followed by 15 mL/kg of body weight 20% (wt/vol) ethanol 30 minutes later each day for 4 weeks. The body weights of the rats were measured two times per week, and food intakes were measured daily. The feed efficiency ratio (FER) was calculated throughout the experiment using these data. After 4 weeks of administration of the test compound, 12-hour-fasted rats were anesthetized during the postabsorptive period. Blood was drawn from the vena cava into heparin tubes and centrifuged at 1,400 g and 4°C; the plasma was stored in a freezer for later assays.
Plasma lipid profile and biochemical parameters
Plasma triglyceride (TG), total cholesterol (TC), and high-density lipoprotein-cholesterol (HDL-C) were enzymatically determined using immunoassay kits (Asia Pharmaceuticals, Seoul) and an enzyme-linked immunosorbent assay reader (Pharmacia Biotech, Cambridge, United Kingdom) according to the manufacturer's protocol. Plasma low-density lipoprotien-cholesterol (LDL-C) was calculated using the Friedewald equation. 12
Parameters of hepatotoxicity, such as aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), and lactate dehydrogenase (LDH) were determined using an enzyme assay kit (Asia Pharmaceuticals).
Histopathology
Liver tissue slices were embedded by the paraffin slice technique, stained with hematoxylin and eosin, and photographed; later the microscopic slides of the liver sections were photographed.
Activities of quinone reductase, CYP2E1, and glutathione S-transferase
The quinone reductase (QR) activity in liver was determined using 2,6-dichloroindophenol and expressed as nanomoles of 2,6-dichloroindophenol reduced per minute per milligram of cellular protein. Liver CYP2E1 activity was determined by the modified method of Patten et al, 13 and liver glutathione S-transferase (GST) activity was determined using 1-chloro-2,4-dinitrobenzene as a substrate in the presence of 0.1 mM reduced glutathione according to the method of Habig et al. 14
Thiobarbituric acid–reactive substances and total glutathione content
Lipid peroxidation in rat plasma, liver, heart, and kidney was determined by the thiobarbituric acid–reactive substances (TBARS) value obtained colorimetrically at 533 nm as previously described. 15 Total glutathione (GSH) level was determined enzymatically according to a previous procedure by Habig et al. 14
Activities of the antioxidative enzymes glutathione peroxidase, GR, and catalase
Glutathione peroxidase (GSH-Px) activity was measured by the methods described by Tappel et al. 16 GR activity was determined using the method described by Pinto et al. 17 CAT activity was determined using the method of Beers and Sizer. 18
DNA damage of lymphocytes
Peripheral blood lymphocytes were isolated by a Ficoll-Paque™ PREMIUM (GE Healthcare, Little Chalfont, United Kingdom) density gradient of centrifugation. Endogenous DNA damage in the lymphocytes was analyzed by alkaline single-cell gel electrophoresis (comet assay) according to the method of Martinet et al. 19
Statistical analysis
Statistical analyses were performed using SAS software (release 8.2, 2000, SAS Institute, Cary, NC). Differences between means were assessed by analysis of variance and Duncan's multiple range test, and statistical significance was defined at P<.05.
Results
Effects of ABG on body weight, food intake, and FER and weights of liver and adipose tissues
The weight gain was not significantly different among the three groups, but the weight gains of the WT and ET+ABG groups were lower than that of the ET group (Table 2). Food intakes and FER were not significantly different between the alcohol-administered ET and ET+ABG groups.
Values with different letters within the row are significantly different by analysis of variance and Duncan's multiple range test (P<.05).
ET group, group in which rats were given a daily dose of 15 mL/kg of 20% ethanol orally (n=7); ET+ABG, group in which rats were given orally a daily dose of 100 mg/5 mL of distilled water/kg aged black garlic (ABG) and 15 mL/kg 20% ethanol in 30-minute intervals (n=7) for 4 weeks; FER, food efficiency ratio (%)=(body weight gain/food intake)×100; WT, group in which rats were given a daily dose of 15 mL/kg saline orally (n=5).
The weights of liver and four major fat pads were determined. Even though the weight of liver was not significantly different among the groups, the liver weight of the ET+ABG group was less than that of the ET group. Among the fat pads, values for the epididymal and total fat pad in the WT and ET+ABG groups were significantly lower than those in the ET group (P<.05) excluding the mesenteric, retroperitoneal, and inguinal fat pad (Table 2).
Effect of ABG on plasma lipid profiles and functional and histopathological changes in livers
The plasma lipid profile and atherogenic index (AI) were not statistically different between the groups administered ethanol for 4 weeks, but the levels of TG, TC, HDL-C, and LDL-C of the alcohol-treated rats were increased compared with those of the WT group (Table 3). Administration of ABG showed the trend of decreased TC, TG and LCD-C and increased HDL-C plasma levels induced by alcohol exposure. Therefore, the AI was reduced in the ET+ABG group compared with that in the chronic alcoholic (ET) group (Table 3).
Values with different letters within the row are significantly different by analysis of variance and Duncan's multiple range test (P<.05).
AI, atherogenic index=(total cholesterol [TC] – high-density lipoprotein-cholesterol [HDL-C])/HDL-C; ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; LDH, lactate dehydrogenase; LDL-C, low-density lipoprotein-cholesterol.
Plasma markers of liver function and injury, such as AST, ALT, ALP, and LDH, were significantly improved by ABG administration (P<.05). Chronic alcohol exposure increased the plasma levels of AST (1.7-fold), ALT (1.5-fold), ALP (1.4-fold), and LDH (1.2-fold) compared with the WT group. In contrast, the ABG-administered (ET+ABG) group showed significantly reduced levels of AST, ALT, ALP, and LDH by 38.7%, 27.3%, 20.4%, and 14.7%, respectively, compared with those of the ET group (P<.05) (Table 3). No significant differences were observed between the WT and ET+ABG groups.
The histopathological examination of livers revealed that ethanol treatment markedly induced prominent fatty changes (arrows) around the portal triad of the liver compared with the WT group (Fig. 1). The increased fat accumulation was found in the liver of the ET group; however, such hepatic fat accumulation was considerably reduced by ABG treatment in the ET+ABG group.

Histopathological changes in the liver: (
Effects of ABG on hepatic CYP2E1, GST, and QR activities
Alcohol-induced metabolic changes or early liver injuries such as steatosis and steatohepatitis 20 result in oxidative stress and increased CYP2E1 expression. In this study, alcohol exposure induced CYP2E1 activity by 2.5-fold in the ET group compared with the WT group. However, ABG treatment significantly decreased alcohol-induced CYP2E1 activity by 45% (P<.05) (Fig. 2). In addition, the activities of drug-metabolizing phase II enzymes such as GST and QR in the liver were significantly improved in the ET+ABG group (P<.05). Activities of hepatic GST and QR in the ET group were 52% and 78% lower than those in the WT rats, and ABG treatment remarkably elevated those activities by 1.5-fold (P<.05) and fourfold (P<.05), respectively, compared with the ET group (Fig. 2).
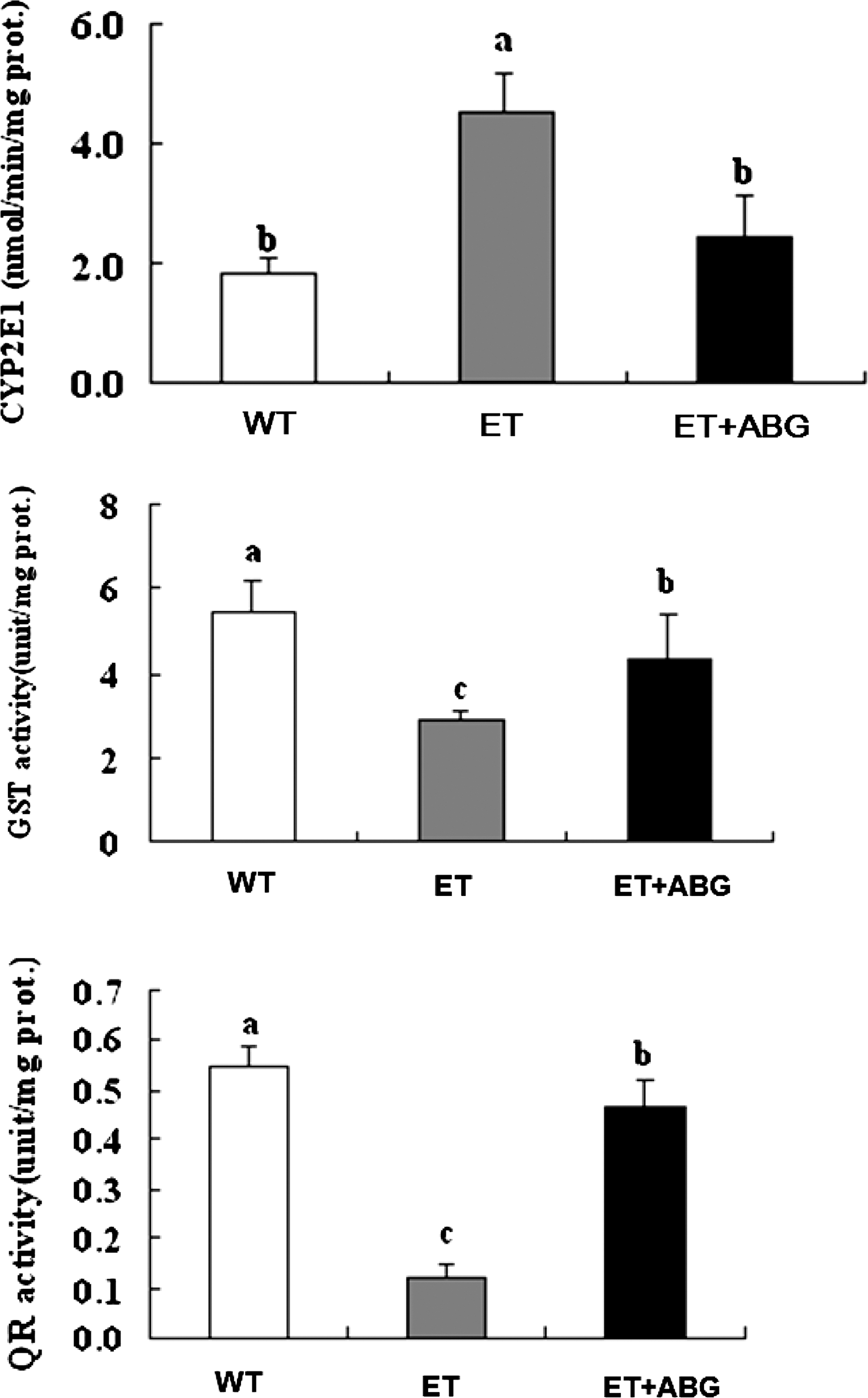
Effects of ABG on hepatic cytochrome P450 2E1 (CYP2E1), glutathione S-transferase (GST), and quinine reductase (QR) activities in rats with chronic alcohol-induced liver injury. abcDifferent letters indicate values are significantly different by analysis of variance and Duncan's multiple range test (P<.05).
Effect of ABG on lipid peroxidation, antioxidants, and activities of antioxidative enzymes in liver and red blood cells
Alcohol as an oxidant can be metabolized to the reactive products acetaldehyde and acetate, leading to a change in the redox state. Such an oxidative state results in the promotion of toxicities such as protein oxidation, lipid peroxidation, oxidative damage of DNA, and fat accumulation in the liver. 21 Lipid peroxidation was determined by measurement of TBARS in the plasma, liver, and heart. Chronic ethanol exposure (ET group) induced a significant increase in TBARS level (P<.05) compared with the control (WT group). However, the increased TBARS levels induced by ethanol treatment were decreased by ABG treatment, showing a significant reduction by 19.8% in liver, 22.9% in plasma, and 23.7% in heart of the ET+ABG group (P<.05) (Fig. 3).

Effect of ABG on lipid peroxidation expressed as thiobarbituric acid–reactive substances (TBARS) values in plasma, liver, and heart. abDifferent letters indicate that values are significantly different by analysis of variance and Duncan's multiple range test (P<.05).
GSH levels in the liver and red blood cells (RBCs) after chronic ethanol exposure were, respectively, 46.22% and 61.53% lower than levels in the WT group (P<.05). ABG dramatically restored GSH levels by 59.9% in liver and 6.4-fold in RBCs compared with the ET group (P<.05). The activities of liver and RBC GSH-Px were also enhanced remarkably in the ET+ABG group. GSH-Px activities increased by 64% in liver and by 73.1% in RBCs in the ET+ABG group compared with the ET group (P<.05). In addition, the activities of liver GR and CAT, which were reduced to 72% and 27% in the ET group compared with the WT group (P<.05), were also restored to control levels in the ET+ABG group. Among the parameters examined, the most sensitive parameters were liver GR activity and RBC GSH level (Table 4).
Values with different letters within a row are significantly different by analysis of variance and Duncan's multiple range test (P<.05).
CAT, catalase; GR, glutathione reductase; GSH, total glutathione; GSH-Px, glutathione peroxidase; RBC, red blood cell.
Effects of ABG on oxidative damage to lymphocyte DNA
The extent of oxidative damage to blood lymphocyte DNA caused by 4-week chronic alcohol digestion was observed by the comet assay. Comet parameters including tail DNA, tail extent moment, olive tail moment, and tail length of the ABG-treated rats significantly decreased by 85.2%, 95.1%, 83.0%, and 74.3%, respectively, compared with those of the alcohol-treated rats (P<.05) (Table 5). Additionally, head DNA of the ABG-treated rats significantly increased by 20.6% compared with that of the alcohol-treated rats (P<.05), meaning that ABG treatment decreased susceptibility of lymphocyte DNA to chronic alcohol-induced oxidative damage.
Values with different letters within the row are significantly different by analysis of variance and Duncan's multiple range test (P<.05).
Discussion
We investigated the antioxidative and hepatoprotective effects of ABG on rats with chronic alcohol-induced liver injury. Administration of ABG significantly decreased epididymal and total fat pads compared with the ET group (P<.05). Moreover, it was confirmed in the histological data that prominent fatty changes around the portal triad and fat accumulation in the liver induced by chronic alcohol treatment were ameliorated by ABG treatment. Long-term alcohol consumption promotes synthesis of fatty acids and TG in the liver by inducing lipid synthetic enzymes such as glycerol-3-phosphate acyltransferase, fatty acid synthase, and malic enzyme as well as by reducing AMP-activated protein kinase activity and β-oxidation through reduction of the cytosolic [NAD+]/[NADH] ratio. 22 The finding that ABG administration inhibits fatty acid and TG synthesis in adipose tissues and livers of rats suggests that ABG may exert an anti-obesity effect as well as hepatoprotective effect against chronic alcohol-induced fatty liver and steatosis.
To investigate the effect of ABG on the liver function in chronic alcoholic rats, analyses of plasma AST, ALT, ALP, and LDH were performed. AST can be found in the liver, cardiac muscle, skeletal muscle, kidney, brain, pancreas, lungs, leukocytes, and erythrocytes, whereas ALT is present in the liver. 23 Increased levels of serum AST and ALT indicate increased permeability and damage to and/or necrosis of hepatocytes 23 and liver injury or inflammation. The membrane-bound enzymes ALP, ALT, and LDH are released unequally into the bloodstream depending on the pathological phenomenon. 24 In this study, ABG administration caused a significant decrease in the activities of AST, ALT, ALP, and LDH in rats with alcohol-induced liver injury, demonstrating that ABG administration protects the liver from chronic alcohol-induced histological and functional injury.
In the mitochondria and microsomes, ethanol is oxidized by CYP2E1 isoenzymes, causing ROS production. 25 The phase I detoxification system, composed mainly of the cytochrome P450 family of enzymes, is the first enzymatic defense against exogenous toxins like alcohol. It is known that the organic sulfur compounds in garlic exert chemoprotective effects against this chemical (alcohol)-induced toxicity in rats and mice. 26 This chemoprotective effect of garlic is explained partly by its ability to decrease the activity of phase I detoxification like CYP2E1 and to some part by inducing phase II detoxifying enzymes such as GST and QR. 27 Consistent with this, our results showed that ABG treatment of chronic alcoholic rats decreased CYP2E1 activity by 55% and increased GST and QR activities by 1.5-fold and fourfold, respectively, compared with those of ethanol-treated rats. These results suggest that the hepatoprotective action of ABG may be due to its suppressive effect on CYP2E1 activity and its inductive effect on GST and QR activities.
Experimental and clinical studies have shown that ethanol ingestion induces oxidative stress in organisms. 28 Increased lipid peroxidation is associated with pathological liver injury in experimental alcoholic liver disease. 29 Likewise, in our study, chronic alcohol ingestion elevated lipid peroxidation and reduced antioxidant content and antioxidative enzyme activities in rats. Of note is that ABG administration remarkably decreased lipid peroxidation in the liver, heart, and plasma, whereas it increased GSH content in the liver and RBCs and the activities of GSH-Px, GR, and CAT in the liver compared with the ET group. Previously, anti-fatty liver effects of garlic oil were observed in mice subjected to 3-day acute ethanol toxicity. 30 Treatment with 100 mg/kg ABG decreased the TBARS level and enhanced the GSH and GR activities in the liver. Accordingly, this suggests that garlic has hepatoprotective effects against both chronic and acute ethanol-induced liver injury of animals through an antioxidative mechanism.
Our results also demonstrated that ABG significantly mitigated alcohol-induced oxidative damage to blood lymphocyte DNA. Antioxidative compounds in ABG might inhibit the oxidative attack of endogenously produced ROS after ethanol exposure. This result is comparable to that of an earlier study 31 in which an antioxidant-rich vegetable and fruit diet containing vitamin C, vitamin E, and β-carotene significantly reduced the DNA tail moment and induced a significant decrease in oxidative DNA damage, finally leading to the resistance of lymphocyte DNA to oxidative damage.
Although garlic has several bioactive activities, its use remains limited because of its pungent smell and flavor. Through the processing of garlic, its pungent smell and flavor can be decreased, and its physiological and functional activities can be enhanced, increasing its bioactive sulfur compound content. Therefore, processed garlic may have efficacy for the prevention and treatment of metabolic diseases such as cancer, atherosclerosis, diabetes, and obesity. Moreover, practical use of processed ABG, which is devoid of adverse effects but tastes good, is not problematic.
This study demonstrated that ABG exerts hepatoprotective effects against chronic alcohol digestion-induced liver injury through antioxidative activities. Consequently, ABG may be a promising agent for protecting against hepatic damage caused by alcohol-induced liver injury.
Footnotes
Acknowledgment
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (NRF 2009-0077171).
Author Disclosure Statement
No competing financial interests exist.