
Abstract
Inflammation and oxidative stress contribute to liver injury. Amla (Emblica officinalis Gaertn.) is rich in vitamin C, gallic acid, flavonoids, and tannins, which may protect against hepatoxicity-induced liver injury. We elucidated the effects of supplementary Amla (100 mg/kg of body weight) on N-nitrosodiethylamine-induced injury by evaluating reactive oxygen species (ROS) responses in the liver and bile, the degree of accumulated leukocytes and Kupffer cell infiltration, 3-nitrotyrosine and 4-hydroxynonenal stains, apoptosis and autophagy, plasma aspartate aminotransferase (AST), alanine aminotransferase (ALT), and γ-glutamyl transpeptidase (γ-GT) levels, and antioxidant/oxidant enzymes in rats. Amla was more potent than vitamin C in scavenging O2 −·, hydrogen peroxide, and nitric oxide. N-Nitrosodiethylamine increased ROS production in liver and bile, hepatic Kupffer cell and leukocyte infiltration, 3-nitrotyrosine and 4-hydroxynonenal accumulations, apoptosis and autophagy, and plasma ALT, AST, and γ-GT levels in the rats, decreased hepatic manganese superoxide dismutase (MnSOD) and catalase protein expressions, and enhanced inducible nitric oxide synthase (iNOS) and cytochrome P450 2E1 (CYP2E1) protein expressions. Amla significantly preserved MnSOD and catalase expressions and decreased iNOS and CYP2E1 protein expressions in N-nitrosodiethylamine-treated livers. Amla decreased N-nitrosodiethylamine-enhanced hepatic apoptosis and autophagy appearances via down-regulation of the Bax/Bcl-2 ratio and Beclin-1 expression. Thus Amla supplementation counteracts N-nitrosodiethylamine-induced liver injury via its antioxidant, anti-inflammation, anti-apoptosis, and anti-autophagy properties.
Introduction
H
Overproduction of reactive oxygen species (ROS), including O2 −·, hydrogen peroxide (H2O2), and nitric oxide (NO), can cause lipid peroxidation, protein oxidation, DNA damage, and mutagenesis associated with various stages of liver injury. 9 –12 The main sources of ROS in the liver may derive from the mitochondria of hepatocytes, the activated macrophages (Kupffer cells), and the infiltrating neutrophils. 9,11 Activations of cytochrome P450 2E1 (CYP2E1) and inducible NO synthase (iNOS) enhance oxidative stress in the damaged liver. 12 ROS also triggered translocation of nuclear factor-κB and activator protein-1 to the nucleus 9 and activated several inflammatory cytokines and adhesion molecules that contribute to ROS production and cell death. 9 The increased levels of ROS may enhance the Bax/Bcl-2 ratio, caspase 3 expression, and poly(ADP-ribose) polymerase (PARP) fragments, resulting in apoptotic cell death, 13,14 and enhance the expression of the autophagy-promoting protein Beclin-1, leading to autophagy. 15 –17 Evidence has shown that an increased ROS amount led to autophagy and apoptosis in HCC. 10 Therefore DEN may promote oxidative stress, inflammation, apoptosis, and autophagy in impaired livers.
Amla (Emblica officinalis Gaertn., Family Euphorbiaceae) is a major constituent of famous Indian polyherbal formulations like Triphala and Chyavanprash used to treat liver diseases. 18,19 The Amla fruits are rich in phenolics, flavonoids, tannins, and ascorbic acid and are known to be hypolipidemic and anti-atherosclerotic. 20 Additionally, Amla possesses antimutagenic and anticarcinogenic properties owing to the combined presence of β-carotene, ascorbic acid, and chlorophyllin and modulates several biochemical events associated with tumor promotion. 21 –24 Although Amla offered protection against DEN-, thioacetamide-, and CCl4-induced liver injury, 25 –27 its hepatoprotective mechanism against DEN-induced oxidative stress, inflammation, apoptosis, and autophagy has not been studied. In this study, we elucidated the effect of Amla on DEN-induced O2 −·, H2O2, and NO activities in the liver in vivo and bile in vitro and correlated them with several liver injury markers, including the degree of accumulated leukocytes and Kuppfer cell (ED-1) infiltration, 3-nitrotyrosine (3-NT) and 4-hydroxynonenal (4-NE) stain, apoptosis and autophagy, plasma aspartate aminotransferase (AST), alanine aminotransferase (ALT), and γ-glutamyl transpeptidase (γ-GT) level, and antioxidant/oxidant status in the rat livers.
Materials and Methods
Chemicals
We used a commercial enzymatic Amla fruit juice extract (SunAmla®, Taiyo Kagaku Co., Ltd., Yokkaichi, Mie, Japan) in this study that contained total polyphenols (29.4±2.2%) and vitamin C (2.0±0.2%) as described previously. 28 It was previously determined that our experimental Amla samples contained total phenol (321 mg of gallic acid/g), total flavonoid (18 mg of rutin/g), total tannin (129 mg of tannin/g), and vitamin C (20 mg of vitamin C/g). 22
The Amla extract was dissolved in water and given orally to rats at a dose of 100 mg/kg of body weight/day for 8 weeks. Control rats were given access to water alone. The reagents H2O2, ascorbic acid, and DEN were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Experimental model of liver injury
Male Wistar rats (weighing 200–250 g) were housed at the Experimental Animal Center, National Taiwan University, Taipei, Taiwan, at a constant temperature and with a consistent light cycle (light from 07:00 to 18:00 hours). Food and water were provided ad libitum. All surgical and experimental procedures were approved by the National Taiwan University College of Medicine and College of Public Health Institutional Animal Care and Use Committee (permission number 20040321) and were in accordance with the guidelines of the National Science Council of Republic of China (NSC 1997). Fifty-four animals were randomly divided into eight groups of six each: group 1, 2 weeks of control water treatment; group 2, 4 weeks of control water treatment; group 3, 8 weeks of control water; group 4, 2 weeks of DEN treatment; group 5, 4 weeks of DEN treatment; group 6, 8 weeks of DEN treatment; group 7, 2 weeks of Amla and DEN treatment; group 8, 4 weeks of Amla and DEN treatment; and group 9, 8 weeks of Amla and DEN treatment. DEN was given as an initiator of liver injury at 500 ppm in the drinking water throughout the entire experiment (8 weeks). Administering DEN in drinking water rather than intraperitoneally reduced the stress and risk of mortality and infection with multiple DEN injections. Amla at a dosage of 100 mg/kg of body weight has been reported to reduce oxidative stress and hepatic injury. 26,27 Therefore, this dosage was selected and was given by oral gavage for different time courses in this study.
Blood biochemical estimation
Whole blood (2 mL) was transferred to an EDTA collection tube and centrifuged at 12,000 rpm (∼13,400 g) for 3 minutes, and the plasma stored at −80°C until analysis. Plasma levels of AST, ALT, and γ-GT were measured using commercially available analytical kits (Sigma) and an automatic analyzer TEXTRON U-240 Plus (Japan Tectron Int. Co., Tokyo, Japan).
In vitro and in vivo chemiluminescence determinations of ROS activity
To compare the antioxidant potential between Amla and vitamin C, we evaluated the inhibitory effect of Amla or vitamin C on O2 −·, H2O2, and NO activity at the dosage of 1, 10, 25, 50, and 100 μg/mL. We used 0.2 mL of Amla or vitamin C samples with 0.1 mL of xanthine (final concentration, 100 μM) plus 0.1 mL of xanthine oxidase (final concentration, 2.5 mU/mL) to measure induced O2 −· activity in 0.5 mL of 0.1 mmol/L lucigenin solution. We evaluated 0.2 mL of Amla or vitamin C samples on 0.2 mL of 0.003% H2O2 in 0.2 mmol/L luminol solution. We also determined NO activity with 0.2 mL of Amla or vitamin C samples by using the NO chemiluminescent probe containing 0.018 mmol/L luminol (Sigma), 10 mmol/L H2O2, 0.15 mmol/L desferrioxamine (Sigma), and 2 mmol/L K2CO3 (Sigma). 11 All O2 −·, H2O2, and NO activity was detected by a chemiluminescence analyzing system (model CLD-110, Tohoku Electronic Inc. Co., Sendai, Japan). 9,11
The ROS generation in response to DEN injury was measured from the liver surface by intravenous infusion (0.2 mg/mL/hour) of a superoxide anion probe, 2-methyl-6-(4-methoxyphenyl)-3,7-dihydroimidazo-[1,2-a]-pyrazin-3-one hydrochloride (MCLA) (TCI-Ace, Tokyo Kasei Kogyo Co. Ltd., Tokyo, Japan), and by the use of a chemiluminescence analyzing system (model CLD-110, Tohoku Electronic Inc. Co.). 9 The rat was maintained on a respirator with a circulating water pad at 37°C in a dark box with a shielded plate. Only the liver window was left unshielded and was positioned under a reflector, which reflects the photons from the exposed liver surface onto the detector area. The real-time–displayed chemiluminescence signal indicated the ROS level on the liver surface.
Bile ROS including O2 −·, H2O2, and NO counts were detected by lucigenin-, luminol-, and modified luminol-amplified methods, respectively. 9,11 In brief, a 0.2-mL bile sample mixed with 0.5 mL of 0.1 mmol/L lucigenin for O2 −· measurement, mixed with 0.2 mmol/L luminol for H2O2 measurement, or mixed with 0.2 mL of an NO chemiluminescent probe for measurement NO as described above was analyzed by a chemiluminescence analyzing system (model CLD-110, Tohoku Electronic Inc. Co.). 9,11 Each assay was performed six times and was expressed as the chemiluminescence count per 10 seconds. 13
In situ demonstration of leukocytes, ED-1 (Kupffer cells), 3-NT, 4-HNE, and autophagy and apoptosis formation in livers
High ROS levels might promote hepatic accumulation of leukocytes, nitrated protein, and lipid peroxides. Accordingly, we immunostained for 3-NT, 4-HNE, 17 ED-1, 9 apoptosis, 9 and autophagy 17 in the paraffin-embedded sections of liver tissues. For these oxidative injury measurements, the rats (n=6 in each group) were sacrificed at the end of the experiment. An 18-gauge needle connected to an infusion pump (model CH-4103, Infors AG, Bottmingen, Switzerland) was inserted into the portal vein. The livers were perfused (flow rate, 10 mL/minute) with Hanks' balanced salt solution (pH 7.4) at 37°C, and the perfusate was allowed to drain from the inferior vena cava. The livers were removed and fixed in 10% formalin for immunostaining.
Liver sections were deparaffinized, rehydrated, and stained immunohistochemically for markers of lipid peroxidation (4-HNE–protein adducts) by incubation with a polyclonal antibody (Alpha Diagnostic International, San Antonio, TX, USA) and with rabbit polyclonal anti-3-NT antibody (Alpha Diagnostic International) and a autophagy marker of Beclin-1 (AnaSpec, Inc., San Jose, CA, USA) diluted at 1:50. The percentage of 3-NT, 4-HNE, or Beclin-1 expression was calculated as (3-NT-, 4-HNE-, or Beclin-1-stained area/total area)×100% and analyzed by Adobe (San Jose) Photoshop version 7.0.1 image software analysis.
For hepatic macrophage (ED-1) staining, the tissue sections were incubated overnight at 4°C with a mouse anti-rat antibody to ED-1 (CD68, diluted 1:200; Serotec, Sydney, NSW, Australia). A biotinylated secondary antibody (Dako, Botany, NSW, Australia) was then applied followed by streptavidin conjugated to horseradish peroxidase (Dako). The chromogen used was Dako liquid diaminobenzene. Twenty high-power (×400) fields were randomly selected for each liver section, and the value of ED-1-positive cells was counted.
The method for the terminal deoxynucleotidyl transferase-mediated nick-end labeling method was performed as described previously. 9,13
Western blotting for manganese superoxide dismutase, catalase, iNOS, CYP2E1, Bax, Bcl-2, PARP, and Beclin-1 in livers
The liver protein concentration was determined by a protein assay from Bio-Rad Laboratories (Hercules, CA, USA). Ten micrograms of protein was electrophoresed as described below. The expression of manganese superoxide dismutase (MnSOD), catalase, iNOS, CYP2E1, Bax, Bcl-2, PARP, and Beclin-1 in liver tissue was evaluated by western immunoblotting and densitometry as described previously. 11,13,14,17 In brief, the total proteins were homogenized with a prechilled mortar and pestle in extraction buffer, which consisted of 10 mM Tris-HCl (pH 7.6), 140 mM NaCl, 1 mM phenylmethylsulfonyl fluoride, 1% Nonidet P-40, 0.5% deoxycholate, 2% β-mercaptoethanol, 10 μg/mL pepstatin A, and 10 μg/mL aprotinin. The mixtures were homogenized completely by vortex-mixing and kept at 4°C for 30 minutes. The homogenate was centrifuged at 12,000 g for 12 minutes at 4°C, the supernatant was collected, and the protein concentrations were determined with the Bio-Rad protein assay.
Polyclonal antibodies raised against MnSOD (Stressgen Bioreagents Ltd., Victoria, Canada), catalase (Chemicon International Inc., Temecula, CA, USA), rat anti-rat CYP2E1 (Chemicon International Inc.), monoclonal mouse anti-human PARP (Promega, Madison, WI, USA), polyclonal rabbit anti-rat Bax (Chemicon), polyclonal rabbit anti-rat Bcl-2 (BioVision, Mountain View, CA, USA), polyclonal rabbit anti-human Beclin-1 (Upstate Biotechnology, Lake Placid, NY, USA), and monoclonal mouse antimouse β-actin (Sigma) were used at 1:400 dilution. All of these antibodies cross-react with the respective rat antigens. Proteins on sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels were transferred to nitrocellulose filters and stained as described. The density of the band with the appropriate molecular mass was determined semiquantitatively by densitometry using an image analyzing system (Alpha Innotech, San Leandro, CA, USA).
Statistical analysis
All values were expressed as mean±SEM. Differences within groups were evaluated by paired t test. One-way analysis of variance was used for establishing differences among groups. Intergroup comparisons were made by Duncan's multiple-range test. Differences were regarded as significant if P<.05 was obtained.
Results
O2 −·, H2O2, and NO activity
We evaluated the antioxidant O2 −·, H2O2, and NO activities of Amla at 1, 10, 25, 50, and 100 μg/mL. As shown in Figure 1, Amla dose-dependently and significantly decreased O2 −·, H2O2, and NO amounts at 25 μg/mL, with near-maximal inhibition of O2 −· and H2O2 activity at 25 μg/mL and a maximal inhibition of NO activity at 1 μg/mL. Vitamin C (from 1 μg/mL to 100 μg/mL) also significantly inhibited O2 −·, H2O2, and NO amounts in a dose-dependent manner. However, Amla is more efficient than vitamin C for inhibiting O2 −·, H2O2, and NO production at dosages of 1 μg/mL and 25 μg/mL.

Dose effect (1, 10, 25, 50, and 100 μg) of Amla and vitamin C (VC) on O2 −·, hydrogen peroxide (H2O2), and nitric oxide (NO) scavenging. Data are mean±SEM values. *P<.05 for comparison of Amla versus VC.
DEN-enhanced ROS in liver and bile
Intravenous infusion of MCLA measured from a baseline level of O2 −·-dependent ROS counts in a range of 1100–1300 counts. After DEN toxicity, the level of O2 −·-dependent ROS from the liver surface was significantly and time-dependently increased from 2 weeks to 8 weeks (Fig. 2). Supplementary Amla (100 mg/kg) administration significantly decreased DEN-enhanced liver ROS production.
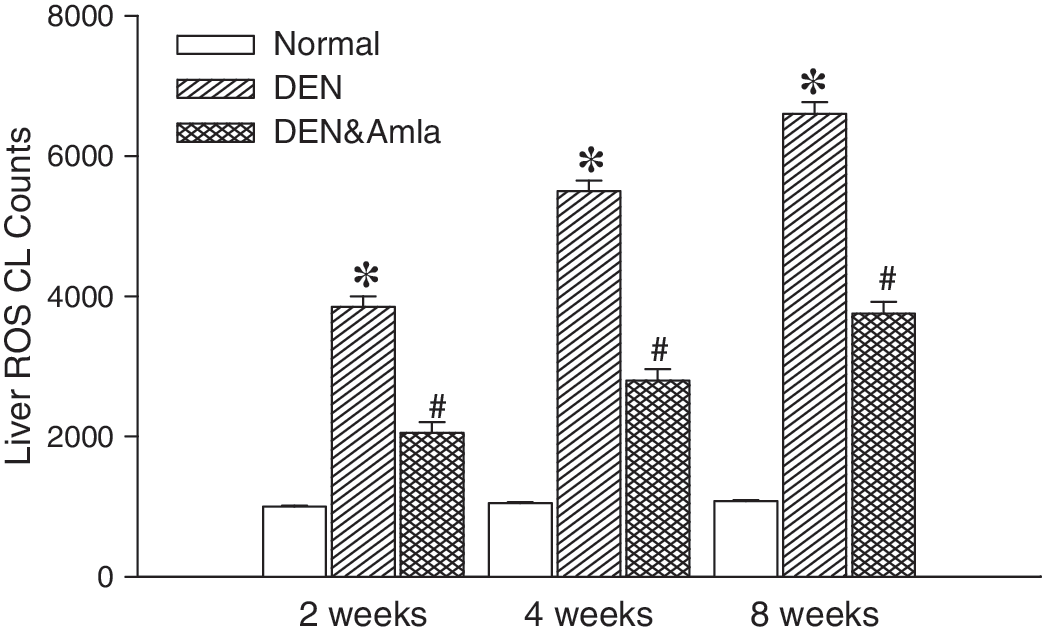
Effects of Amla on N-nitrosodiethylamine (DEN)-induced liver reactive oxygen species (ROS) chemiluminescence (CL) counts in vivo. The liver ROS level was time-dependently (from 2 weeks to 8 weeks) increased after daily DEN treatment. Oral Amla by gavage significantly reduced liver ROS CL counts during the 2–8-week period. Data are mean±SEM values (n=6). *P<.05 compared with the normal group; # P<.05 compared with the respective DEN group.
The increased hepatic ROS production was accompanied by the increased biliary O2 −·, H2O2, and NO levels, and the increased level was consistently and time-dependently elevated (Fig. 3). The increased level of hepatic and biliary ROS was significantly reduced by the Amla treatment.

Effect of Amla on DEN-enhanced bile O2 −·, H2O2, and NO production. DEN time-dependently enhanced bile O2 −·, H2O2, and NO production. Pretreatment with Amla significantly decreased DEN-enhanced bile O2 −·, H2O2, and NO production after 2–8 weeks of treatment. Data are mean±SEM values (n=6). *P<.05 compared with the respective normal value; # P<.05 compared with the respective DEN value.
DEN-enhanced plasma biochemical parameters
Supplementary Amla did not affect plasma ALT, AST, and γ-GT levels in the rats without DEN treatment. DEN-induced liver injury is significantly associated with time-dependent increases in plasma ALT, AST, and γ-GT levels (Fig. 4). The plasma ALT, AST, and γ-GT levels increased by DEN treatment were significantly suppressed by Amla treatment.

Effect of Amla on plasma alanine aminotransferase (ALT), aspartate aminotransferase (AST), and γ-glutamyl transferase (γ-GT) concentration in DEN-treated rats. DEN time-dependently increased plasma ALT, AST, and γ-GT levels in the rats. Intragastric Amla significantly decreased DEN-enhanced plasma ALT, AST, and γ-GT concentrations. Data are mean±SEM values (n=6). *P<.05 compared with the respective normal value; # P<.05 compared with the respective DEN value.
DEN-induced leukocyte infiltration, Kupffer cells, 3-NT, and 4-HNE
Histological examination of DEN-treated liver sections found increased leukocyte infiltration in the damaged liver parenchyma (Fig. 5B–D) in a time-dependent manner. In contrast, the histological appearance of liver sections in the control (Fig. 5A) and Amla-treated (Fig. 5E) groups were normal. Amla treatment significantly protected rats from DEN-induced liver damage and decreased DEN-induced leukocyte accumulation (Fig. 5F–H). ED-1 stain for Kupffer cells was less evident in the control (Fig. 5I) and Amla-treated (Fig. 5M) livers but was significantly enhanced in DEN-treated liver in a time-dependent manner (Fig. 5J–L). There was a significant decrease after Amla treatment in DEN-treated liver (Fig. 5N–P).
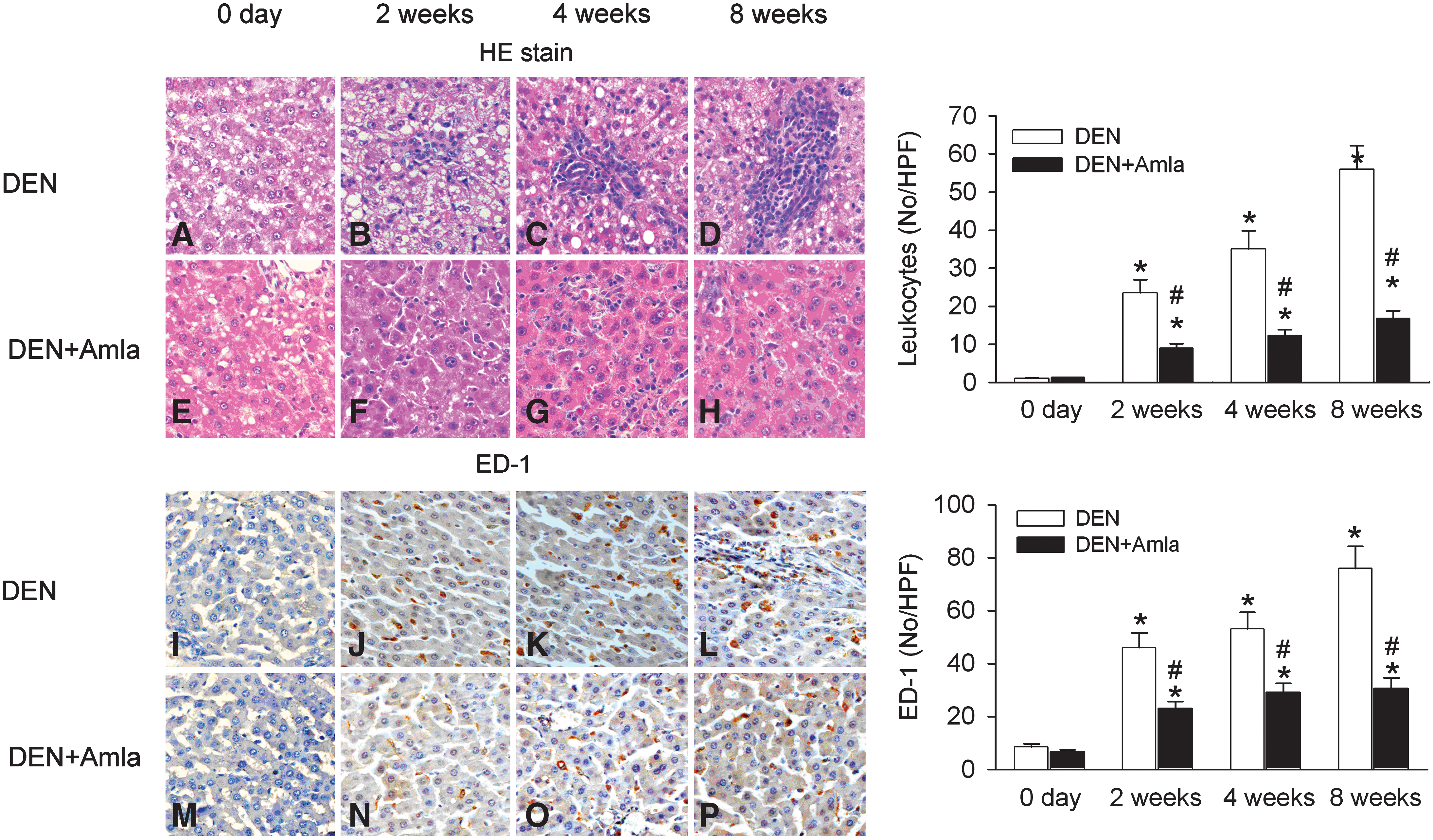
Effect of Amla on inflammation indicated by infiltrated leukocytes (blue spots,
Significant time-dependent 4-HNE lipid accumulation was detected by 4-HNE staining in DEN-treated liver (Fig. 6B–D) compared with the control (Fig. 6A) and Amla-treated (Fig. 6E) livers. Amla treatment decreased the appearance of 4-HNE stain (Fig. 6F–H).
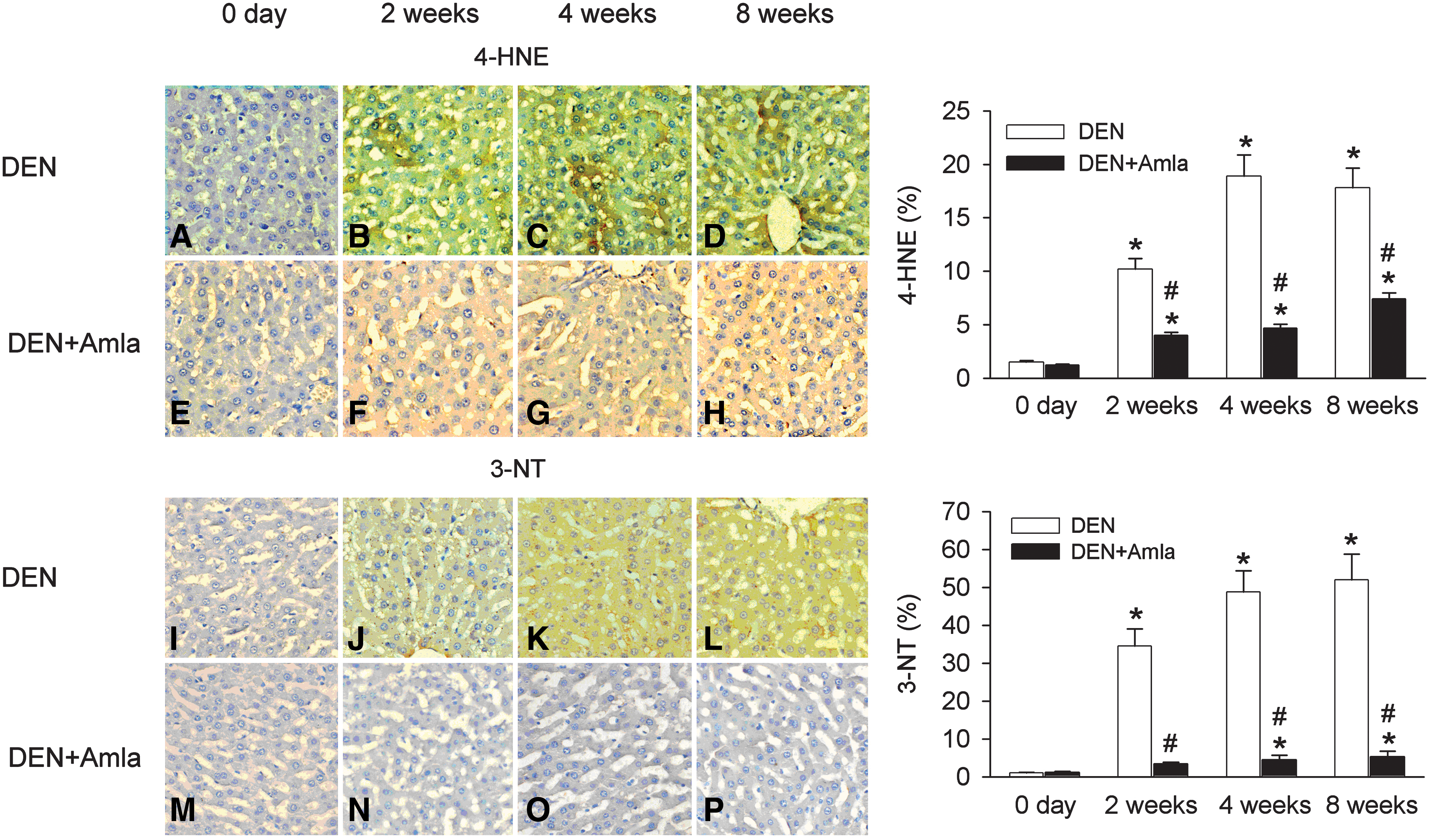
Effect of Amla on oxidative stress indicated by 4-hydroxynonenal (4-HNE) (brown stain,
Significant time-dependent 3-NT accumulation was detected in DEN-treated liver by 3-NT staining (Fig. 6F–H) compared with the control (Fig. 6A) or Amla-treated (Fig. 6E) liver. Amla treatment decreased the appearance of 4-HNE stain (Fig. 6N–P).
Antioxidant, inflammatory, and oxidant protein expression
DEN treatment decreased MnSOD and catalase expression in the livers and time-dependently enhanced hepatic iNOS and CYP2E1 protein expression (Fig. 7). Amla partially preserved MnSOD and catalase expression and decreased iNOS and CYP2E1 protein expression in the DEN-treated livers.
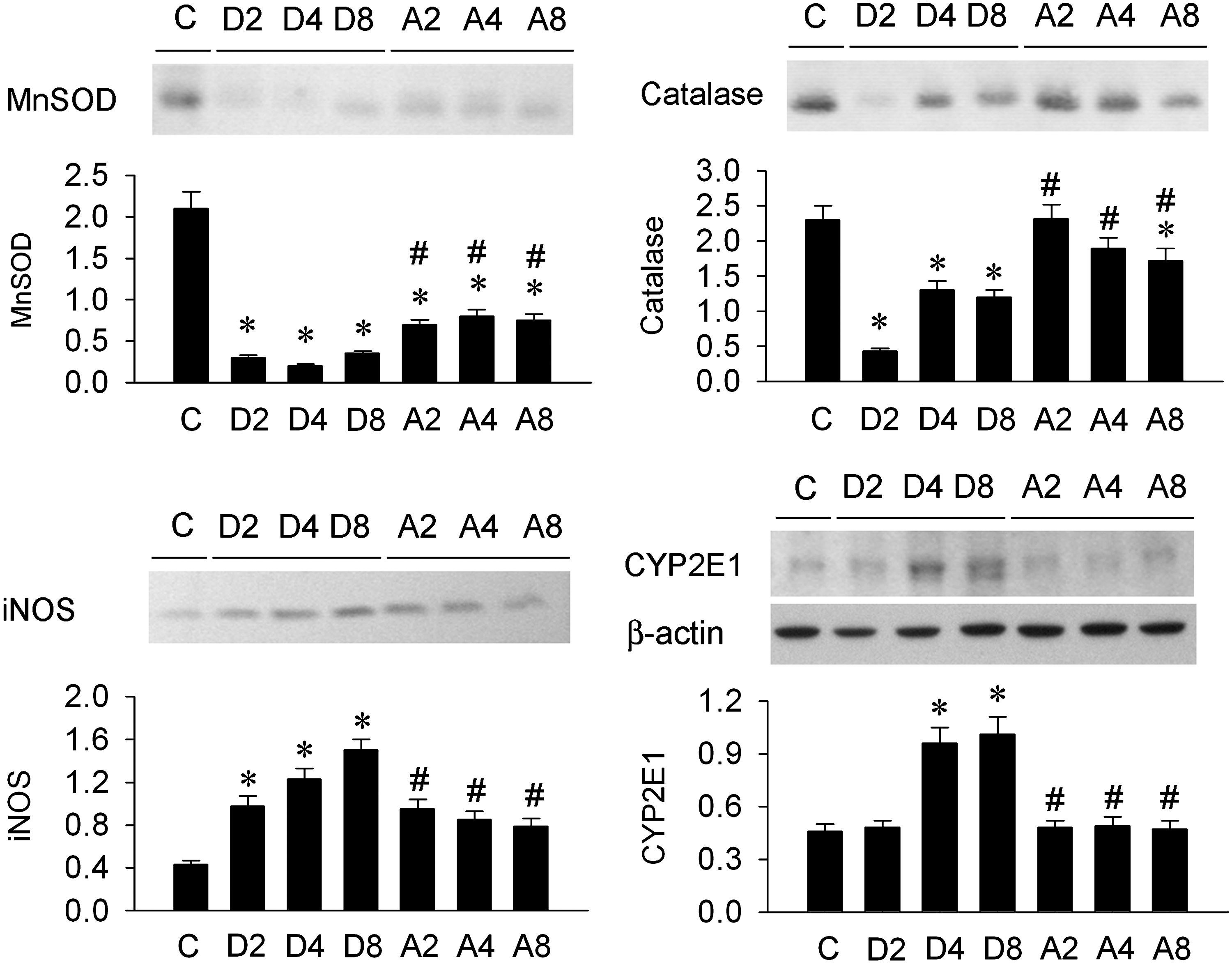
Representative photomicrographs and quantitative evaluation of manganese superoxide dismutase (MnSOD), catalase, inducible NO synthase (iNOS), and cytochrome P450 2E1 (CYPE2E1) protein levels in the rat livers under control conditions (C), after 2 weeks (D2), 4 weeks (D4), and 8 weeks (D8) of DEN treatment, or after Amla pretreatment and 2 weeks (A2), 4 weeks (A4), and 8 weeks (A8) of DEN treatment. Data are mean±SEM values (n=6). *P<.05 compared with the respective C value; # P<.05 compared with the respective DEN value.
Apoptosis- and autophagy-related proteins and production
We explored Bcl-2, Bax, PARP, and Beclin-1 expression in DEN-treated livers with or without Amla pretreatment by western blotting (Fig. 8A). Bax and Bcl-2 were detected in livers, and levels were enhanced post-DEN treatment. However, Bcl-2 expression was more enhanced by Amla treatment. The enhanced Bax/Bcl-2 ratio was depressed by Amla treatment. PARP expression was enhanced by DEN treatment but was depressed by Amla treatment. Beclin-1 was rarely detected in the normal liver but was greatly increased after DEN treatment. Amla depressed DEN-enhanced Beclin-1 expression. DEN significantly and time-dependently increased the percentage of apoptosis (Fig. 8B) and autophagy (Fig. 8C) appearance in the liver, whereas Amla significantly decreased the DEN-enhanced apoptosis and autophagy percentage.
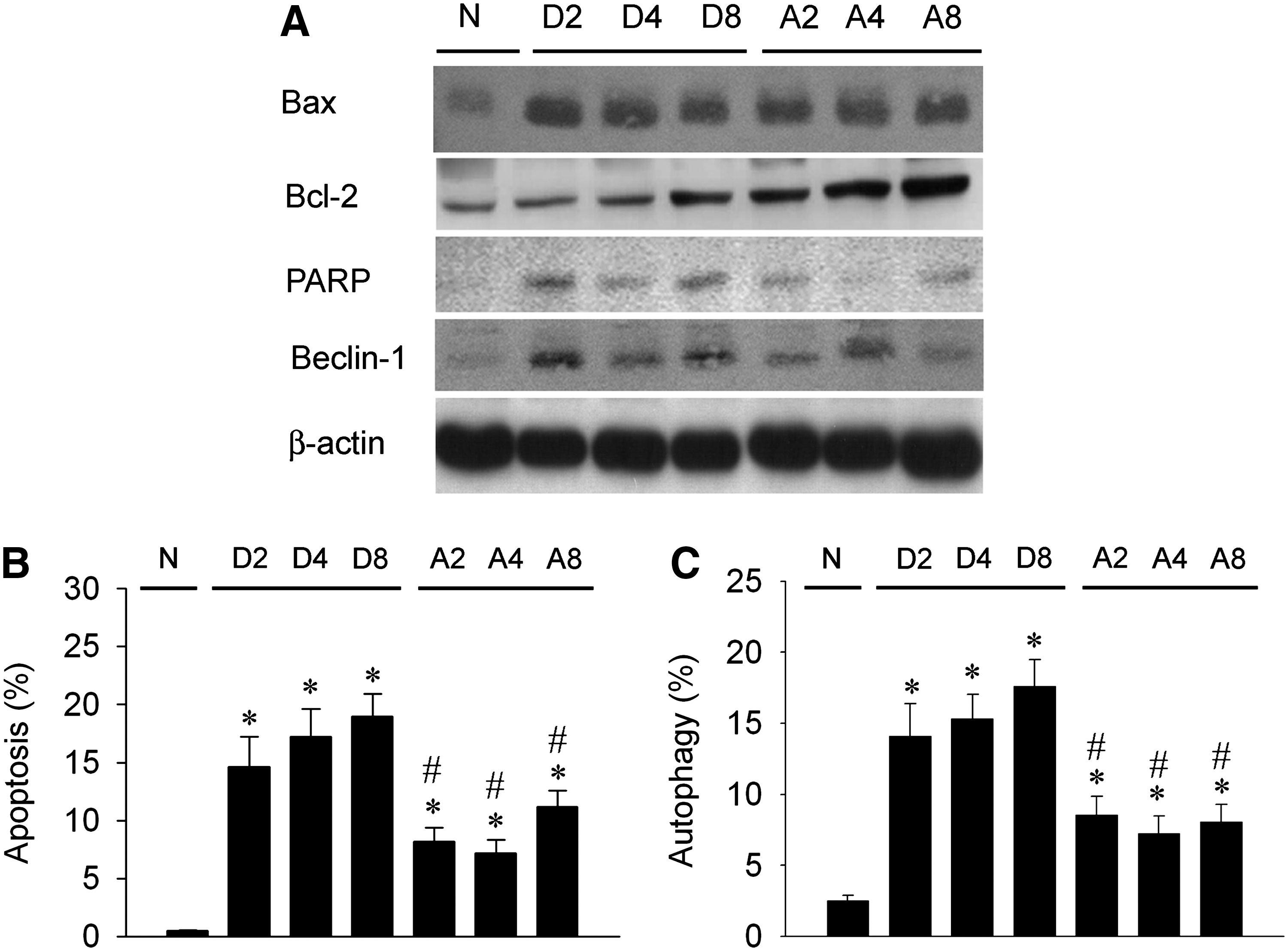
(
Discussion and Conclusions
Amla was stronger than vitamin C in reduction of O2 −·, H2O2, and NO activity in our in vitro experiment. In the in vivo study, we demonstrated DEN-induced liver injury by the increased ROS levels in liver and bile, increased neutrophil and ED-1 cell (monocyte) infiltration, 3-NT and 4-HNE accumulation, increased hepatocyte apoptosis and autophagy formation, and elevated plasma ALT, AST, and γ-GT levels. We also found a decreased MnSOD and catalase protein expression and a higher CYP2E1 and iNOS protein expression after DEN treatment, indicating increased oxidative stress. Supplementary Amla significantly attenuated the DEN-enhanced ROS levels in liver and bile, neutrophil and ED-1 cell infiltration, 3-NT and 4-HNE accumulation, CYP2E1 and iNOS protein expression, and plasma ALT, AST, and γ-GT levels. Amla also partially preserved MnSOD and catalase protein expression and depressed CYP2E1 and iNOS expression in the DEN-treated livers. Our results suggest that Amla may counteract DEN-induced hepatotoxicity, possibly via an enhancement of antioxidant defense mechanisms and a down-regulation of inflammation, apoptosis, and autophagy signaling pathways.
Previous studies showed that the active ROS scavenging components of Amla include tannoids, vitamin C-like materials, and tannins.
29
–31
In aqueous extracts of dried Amla fruits, vitamin C accounts for approximately 45–70% of the antioxidant activity.
31
Therefore, it is conceivable that the antioxidant activity of Amla was not only due to the high content of vitamin C, but also partly attributable to tannins, such as emblicanin A and emblicanin B.
29,31
Amla used in this study containing total polyphenols (29.4±2.2%) and vitamin C (2.0±0.2%) as previously described.
32
Components of polyphenols in the Amla extracts have been identified as follows: 3.458% gallic acid, 0.635% corilagin, 0.880% geraniin, 0.659% elaeocarpusin, 1.240% chebulagic acid, and 0.220% ellagic acid.
32
Other components like mucic acid 1,4-lactone 3-O-gallate, mucic acid 1,4-lactone 2-O-gallate, 1,6-di-O-galloyl-β-
We applied an enhanced chemiluminescence method to measure ROS, including O2 −·, H2O2, and NO production in the liver, blood, and bile. 9,11 This method has been well established for the determination of ROS production in cultured cells, whole blood, and urinary bladder, kidney, and liver in vivo. 9,13,14,33 Exacerbated production of ROS has been reported in various forms of liver injury. 9 –11,34 ROS are regularly produced as a result of carcinogen treatment causing oxidative stress that leads to damage of nucleic acids, proteins, and lipids, resulting in chromosomal instability, mutations, loss of organelle function, and membrane damage that play important mechanistic roles in the development of cancer. 10 In this study, the higher level of chemiluminescence was detected from the liver surface and bile secretion after DEN treatment. More specifically, a multifold increase in levels of O2 −·, H2O2, and NO was found in the bile. Western blotting revealed higher CYP2E1 and iNOS protein expression and lower MnSOD and catalase protein expression in the DEN-treated livers. These changes may result in the increased O2 −·, H2O2, and NO level. By hematoxylin and eosin stain and immunocytochemistry, we further found that the cellular source of hepatic ROS may come from affected hepatocytes, neutrophils, and Kupffer cells in DEN-treated liver. The increased 3-NT and 4-HNE oxidative product accumulation also evidenced DEN-induced oxidative stress in the livers. Our data also showed that higher ROS levels and several oxidative stress biomarkers were continuously maintained during 8 weeks of drinking DEN-containing water. These data suggest that chronic oxidative stress and inflammation occurred in the DEN-treated model.
Increased numbers of and activated Kupffer cells play a critical role in the pathogenesis of several inflammatory diseases, 35 such as alcoholic liver disease 9,35,36 and liver ischemia/reperfusion injury. 37,38 Kupffer cells can be activated to produce O2 −·, NO, and tumor necrosis factor-α release by hypoxia, toxicity, or tissue reperfusion. 35,38 –40 The increased oxidative stress and inflammatory mediators may lead to secondary vasoconstriction and disturbance in hepatic microcirculation and further aggravate liver injury. 41,42 Increased ROS are usually associated with the elevation of several inflammatory markers like interleukin-6, tumor necrosis factor-α, and prostaglandins. 33,43,44 Therefore, the use of antioxidants may depress inflammatory cytokines and chemokines. Oral administration of berberine (50 mg/kg) inhibited the iNOS expression and CYP2E1 activity in DEN plus phenobarbital–treated livers. 12 Polyphenolic compounds found in various plants down-regulate inflammatory responses through inhibiting iNOS, cyclooxygenase-2, and CYP2E1 via their inhibitory effects on nuclear factor-κB or activator protein-1, thereby exerting antioxidant, anti-inflammatory, and anti-aging effects. 9,43,44 Several lines of evidence suggest that supplementation with Amla extracts reduces expression of inflammatory biomarkers such as 8-iso-prostaglandin, the neutrophil chemokines interleukin-8, GRO-α, and GRO-γ, adhesion molecule intercellular adhesion molecule-1, and pro-inflammatory cytokine interleukin-6 and interleukin-1β. 44 –47 A possible anti-inflammatory mechanism of Amla extracts is selective competition with the binding nuclear factor-κB to its specific target DNA sequences and therefore decreasing the expression levels of interleukin-6. 47 In our results, Amla significantly attenuated DEN-induced ROS production and iNOS and CYP2E1 expression and decreased the infiltration of neutrophils and Kupffer cells in the livers, suggesting that Amla is an effective anti-inflammatory agent.
Overt ROS contribute to apoptosis and autophagy cell death. 13,15 –17,48,49 Exacerbated ROS trigger mitochondrial permeability transition pore opening by shifting its gating potential, 50 impairing ATP synthesis and subsequently leading to autophagy, apoptosis, and necrosis. 51 Sometimes, cellular features of necrosis, apoptosis, and autophagy occur together after death signals and toxic stresses. 51 Our data showed that increased Beclin-1-dependent autophagy and Bax/Bcl-2-dependent apoptosis production concomitantly occurred in DEN-treated liver. These increases in autophagic and apoptotic cell death in the hepatocytes may destroy the integrity of the hepatocyte plasma membrane, mainly in the canalicular domain, and consequently liberate ALT, AST, and γ-GT into plasma. Amla treatment significantly decreased DEN-induced apoptosis and autophagy and reduced plasma ALT, AST, and γ-GT levels in liver, demonstrating that Amla treatment preserved the functional and morphological integrity of the DEN-treated livers, possibly up-regulating Bcl-2 in the livers. The Drosophila spin gene product (HSpin1) resulted in a caspase-independent autophagic cell death without inducing a release of cytochrome c from mitochondria. 52 HSpin1 bound to Bcl-2 and Bcl-xL, and overexpression of Bcl-2 inhibited the HSpin1-induced autophagic cell death. 52 Our results showed that Bcl-2 was up-regulated after Amla supplementation and decreased the DEN-enhanced Bax/Bcl-2 ratio, PARP fragment-related apoptosis, and Beclin-1-induced autophagy in the rat livers.
In summary, Amla with high contents of polyphenols and vitamin C decreases O2 −·, H2O2, and NO activity. Amla supplementation significantly attenuates DEN-induced liver injury by antioxidative, anti-inflammatory, anti-apoptotic, and anti-autophagic mechanisms. We suggest that Amla is promising as an effective drug or healthy food for cancer prevention and cellular protection of the liver.
Footnotes
Acknowledgments
This work was supported in part by the Department of Medical Research at the National Taiwan University Hospital and from grants NTUH98-S1141 and NTUH98-M1216, and partly by the National Science Council of the Republic of China (grants NSC96-2320-B-002-007 and NSC96-2221-E-002-256-MY3).
Author Disclosure Statement
No competing financial interests exist.