
Abstract
The beneficial effects of exercise on overall health make it desirable to identify the orally active agents that enhance the effects of exercise in an effort to cure metabolic diseases. Natural compounds such as resveratrol (RSV) are known to increase endurance by potentiating mitochondrial function. Korean mistletoe (Viscum album coloratum) extract (KME) has characteristics similar to those of RSV. In the present study, we determined whether KME could increase mitochondrial activity and exert an anti-fatigue effect. We found that KME treatment significantly increased the mitochondrial oxygen consumption rate (OCR) in L6 cells and increased the expression of peroxisome proliferator-activated receptor γ coactivator (PGC)-1α and silent mating type information regulation 2 homolog 1 (SIRT1), two major regulators of mitochondria function, in C2C12 cells. In the treadmill test, KME-treated mice could run 2.5-times longer than chow-fed control mice. Additionally, plasma lactate levels of exhausted mice were significantly lower in the KME-treated group. In addition, the swimming time to exhaustion of mice treated with KME was prolonged by as much as 212% in the forced-swim test. Liver and kidney histology was similar between the KME-treated and phosphate-buffered saline-treated animals, indicating that KME was nontoxic. Taken together, our data show that KME induces mitochondrial activity, possibly by activating PGC-1α and SIRT1, and improves the endurance of mice, strongly suggesting that KME has great potential as a novel mitochondria-activating agent.
Introduction
M
Mitochondrial dysfunction, and thus ATP production issues, is involved in several human diseases, including metabolic diseases, 7 cardiac dysfunction, 8 mental disorders, 9 and aging-related diseases. 10 RSV is one of the most potent activators of mitochondrial activity; RSV treatment increases mitochondrial oxygen consumption rate, resulting in an increase in exercise endurance and life span. 11,12
Peroxisome proliferator-activated receptor γ coactivator (PGC)-1α is a coactivator of nuclear receptors, in addition to other transcription factors. It controls several elements of energy metabolism. Alterations in mitochondrial function due to coordinated gene expression are induced by environmental stimuli. PGC-1α is a potent regulator of mitochondrial gene expression. PGC-1α transgenic animals also show physiological improvements in whole-body oxygen uptake and decreased fatigue in response to an exercise stimulus. 13 In addition, PGC-1β is a PGC-1 (Peroxisome proliferator-activated receptor-2 [PPAR2] coactivator-1) coactivator and is a homolog of PGC-1α; 14 it coactivates a variety of nuclear receptors and transcription factors 15,16 that are targeted by PGC-1α's target transcription factors. PGC-1β regulates mitochondrial metabolism 17,18 and promotes mitochondrial proliferation and muscle cell respiration.
Sirtuin 2 (Sir2) belongs to the silent information regulator family of proteins. These proteins are deacetylated in an NAD+-dependent manner, yielding nicotinamide (the deacetylated product) and a novel metabolite, O-acetyl-ADP-ribose. 19 In mammals, seven sirtuin genes have been identified (SIRT1–SIRT7). 19 Of the seven mammalian sirtuins, SIRT1 is the closest homolog to Sir2. SIRT1 interacts with PGC-1α, which regulates mitochondrial biogenesis and is sensitive to metabolic alteration. 20 In the skeletal muscle, exercise increases mitochondrial biogenesis and alters energy metabolism by increasing PGC-1α expression. 21,22 Recent evidence has demonstrated that selective PGC-1α deacetylation by SIRT1 can alter skeletal muscle metabolism and mitochondrial biogenesis. 11,23
In this study, we determined whether KME treatment increases mitochondrial activity in vivo and in vitro. Our data revealed that KME enhances mitochondrial activity, possibly by activating PGC-1α and SIRT1, increases mitochondrial function, as evidenced by increases in the running and swimming time of mice, and by increased resistance to muscle fatigue, possibly due to the increased expressions of several PGC-1α target genes.
Materials and Methods
Extraction of KM
KM growing on oak was harvested in February in Kangwondo, Korea. The mistletoe was 1 or 2 years old at the time of harvesting. The leaves, stems, and fruits were cut into two joints from the end of a branch. They were then washed with distilled water (DW) and dried. The vacuum-packed mistletoe was stored at−80°C until extraction. The freeze-dried leaves, fruits, and stems of mistletoe were mashed up and ground in ∼10 volumes of DW for 30 sec. After washing, they were ground in a mixer for 2 min and stirred for 16 h at 4°C. To obtain the mistletoe extract, mistletoe was centrifuged at 6500 g for 30 min at 4°C, and the suspension was filtered through 0.9 and 0.45 μm pores, successively. The mistletoe extract was freeze dried (yield 15%) and resuspended in phosphate-buffered saline (PBS) at an appropriate dilution factor.
Cell culture and KME treatment
The rat skeletal muscle cell line L6 (rat myoblast cell line) was provided by the Asan Institute for Life Science, and the mouse muscle cell line C2C12 (mouse myoblast cell line) was provided by the POSTECH. L6 and C2C12 cells were cultured in Dulbecco's Modified Eagle's medium (DMEM; Invitrogen) supplemented with complete medium, 10% fetal bovine serum (FBS; CAMBREX), and 1% penicillin/streptomycin (CAMBREX), at 37°C under a humidified 95%/5% (v/v) mixture of air and CO2. L6 cells were cultured in complete medium to ∼90% confluence. Before drug treatment, cells were washed with PBS and incubated with DMEM containing 0.5% FBS for 16 h. Cells were then treated for 48 h with KME (6 μg/mL) or PBS. C2C12 cells were maintained in DMEM containing 10% FBS until the cells were ∼80% confluent. To induce myotube differentiation, media were replaced with DMEM containing 1% FBS when the cells reached 80% confluence. The experimental procedure commenced after 7 days of culture in DMEM containing 1% FBS. C2C12 myotubes were treated with KME (6 μg/mL) or PBS for 24 h.
Animals care
Pathogen-free 12 week-old male ICR mice were purchased from the Hyochang Science (
Endurance tests
Endurance was measured on a treadmill (Columbus Instruments) enclosed in a plexiglass chamber that was outfitted with a shock grid at the rear of the belt to keep the animal running during the test and forced-swim test. The shock grid delivered a 0.2-mA shock, which was uncomfortable, but did not physically harm or injure the animals. The animals were habituated to the test conditions before experimentation. During the test procedure, the mice ran on a treadmill with a 5° incline at a speed of 16.2 m/min for 10 min. For the chow-fed mice, the speed was first set at 15 m/min with a 5° incline. The speed was then gradually increased from 15 to 24.6 m/min and was maintained at 24.6 m/min until exhaustion.
11
When the mice reached exhaustion, as defined by the inability to run for 10 s,
24
the electric shock was discontinued. Supplementary Figure S1 (Supplementary Data are available online at
Analysis of blood biochemical parameters
After the mice had been killed using CO2 gas, whole blood samples were collected in a vacutainer tube, prepared with sodium fluoride and potassium oxalate by heart puncture. Plasma was prepared by centrifugation at 2500 g at 4°C for 10 min and then stored at −80°C. The level of plasma lactate was analyzed with commercial kits (BioVision, Inc.); the levels of plasma glucose were analyzed using commercial kits (Roche Diagnostics).
Measurement of tissue glycogen contents of mice
Immediately after blood collection, the gastrocnemius muscle and livers were quickly dissected out and kept at −80°C until they were analyzed for glycogen content. The glycogen content was spectrophotometrically measured using the glucose oxidase method of Chun et al. 26 Briefly, after digestion of the gastrocnemius muscle and liver samples in 30% KOH at 100°C for 30 min, 1.5 mL of 95% alcohol was added to the vials. After centrifugation at 3000 rpm for 10 min, the supernatant was discarded, the residue was suspended in 0.5 mL of distilled water, and 1 mL of 0.2% anthrone in H2SO4 was added. The vials were placed in a boiling water bath for 30 min. The absorbances of the solutions were determined using a FLUOStar OPTIMA microtiter plate reader (BMG Labtech GmbH) at 620 nm wavelength.
RNA extraction, cDNA synthesis, and real-time reverse transcriptase–polymerase chain reaction
Total RNAs were isolated from C2C12 myotube cells using the TRIzol reagent (Invitrogen). Four micrograms of total RNA were used for first-strand cDNA synthesis using the Superscript II reverse transcriptase (Invitrogen). The materials and methods used for this analysis are described in detail in supplementary data. The primers used are listed in Supplementary Table S1.
Measurement of oxygen consumption
The O2 consumption rate was determined by high-resolution respirometry using an Oxygraph-2k instrument (Oroboros Instruments). The amplified signal from the oxygen sensor was recorded on a computer at sampling intervals of 1 s (DatLab acquisition software; Oroboros Instruments). The respiration medium was equilibrated with air in the oxygraph chambers at 37°C and stirred until a stable signal was obtained for calibration. After calibration, the medium was replaced with 2.0 mL aerated cell suspensions, and the chambers were sealed with a stopper. Typically, the respiration rate stabilized within 5–10 min of measurement, as indicated by a linear depletion of oxygen in the chamber. Once the respiration rate stabilized, the cells were trypsinized and suspended at 1×107 cells/mL in medium. Experimental data were corrected for the entire range of O2 concentrations using DatLab software (Oroboros Instruments), which allows for the subtraction of background oxygen consumption. Baseline cell respiration was determined, and FCCP (carbonyl cyanide p-[trifluoromethoxy]-phenyl-hydrazone) was used to estimate the maximum (uncoupled) respiratory rate.
Histologic analysis
In preparation for hematoxylin and eosin (HE) staining, the tissues were fixed in 10% formalin and embedded in paraffin. Sections (3 μm in thickness) were affixed to slides, deparaffinized, dehydrated, and then stained with HE. Stained liver and kidney sections were observed under a light microscope (BX 50; Olympus). Each sample was observed at 400×magnification. The liver and kidney tissues from one mouse from the control, 400 mg/kg KME, and 1000 mg/kg KME groups were analyzed.
Statistics
Statistical analyses was performed using GraphPad Prism version 4.0 (GraphPad Software) and was conducted by using unpaired two-tailed Student's t-test or one-way analysis of variance and, subsequently, Tukey's test was applied (GraphPad Prism 5). A P value<.05 was considered statistically significant.
Results and Discussion
KME treatment increased the mitochondrial oxygen consumption rate in L6 myoblast cells
Cellular energy expenditure was investigated by measuring the oxygen consumption rate (OCR) using a Clark-type electrode. After treatment with KME for 48 h, we observed a 37% acceleration of OCR (Fig. 1). The effects of mitochondrial uncoupling were measured by treatment with the uncoupling agent, FCCP. In mitochondria from KME-treated cells, FCCP noticeably accelerated oxygen consumption (∼35%), indicating that the maximal capacity of the respiratory chain was enhanced (Fig. 1). It is an arduous task to identify molecules that increase oxygen consumption which are not direct mitochondrial uncouplers. However, xenobiotic molecules found in dietary sources, such as kaempferol (KPF), are small polyphenolic molecules that increase cellular energy expenditure. 27 Treatment with KPF increases oxygen consumption by ∼30% in skeletal myocytes, which is comparable to the ∼35%–37% increase induced by KME treatment. It will, therefore, be of great interest to investigate the expressions of various genes involved in this activity, including GLUT4, PGC-1α, UCP-3, and Dio2 (type 2 deiodinase), to gain an insight into the molecular mechanisms triggered by KME.

Oxygen consumption rates of control and KME-treated cells. Oxygen consumption rates (OCR; pmol oxygen/[s·mg protein]) were measured in L6 cells. Cells were treated for 48 h with KME (6 μg/mL). FCCP (5 μmol/L) was used to uncouple mitochondrial respiration. Values represent the mean±SEM. (*P<.05, n=4). KME, Korean mistletoe extract. FCCP, carbonyl cyanide p-[trifluoromethoxy]-phenyl-hydrazone.
KME increased endurance capacity in mice
Since KME treatment increased OCR (Fig. 1) in vitro, we evaluated the effects of KME administration in vivo using the treadmill endurance test. We treated ICR mice with KME at 400 or 1000 mg/(kg·d) for 1 week and then performed a treadmill endurance test. Surprisingly, KME-treated (1000 mg/kg) mice ran a distance that was 2.5-times longer than the distance run by the PBS-fed mice (Fig. 2B). Thus, KME treatment significantly increased the resistance to muscle fatigue. This result is consistent with the observed increase in OCR. Recently, Narkar et al. 28 reported that AMP-activated protein kinase and PPARδ agonists, such as 5-aminoimidozole-4-carboxamide ribotide (AICAR) and GW1516, enhanced running endurance by 44% and suggested that these agonists might be used to enhance training adaptation or even to increase endurance without exercise. Seemingly, KME is more effective than AICAR and GW1516.
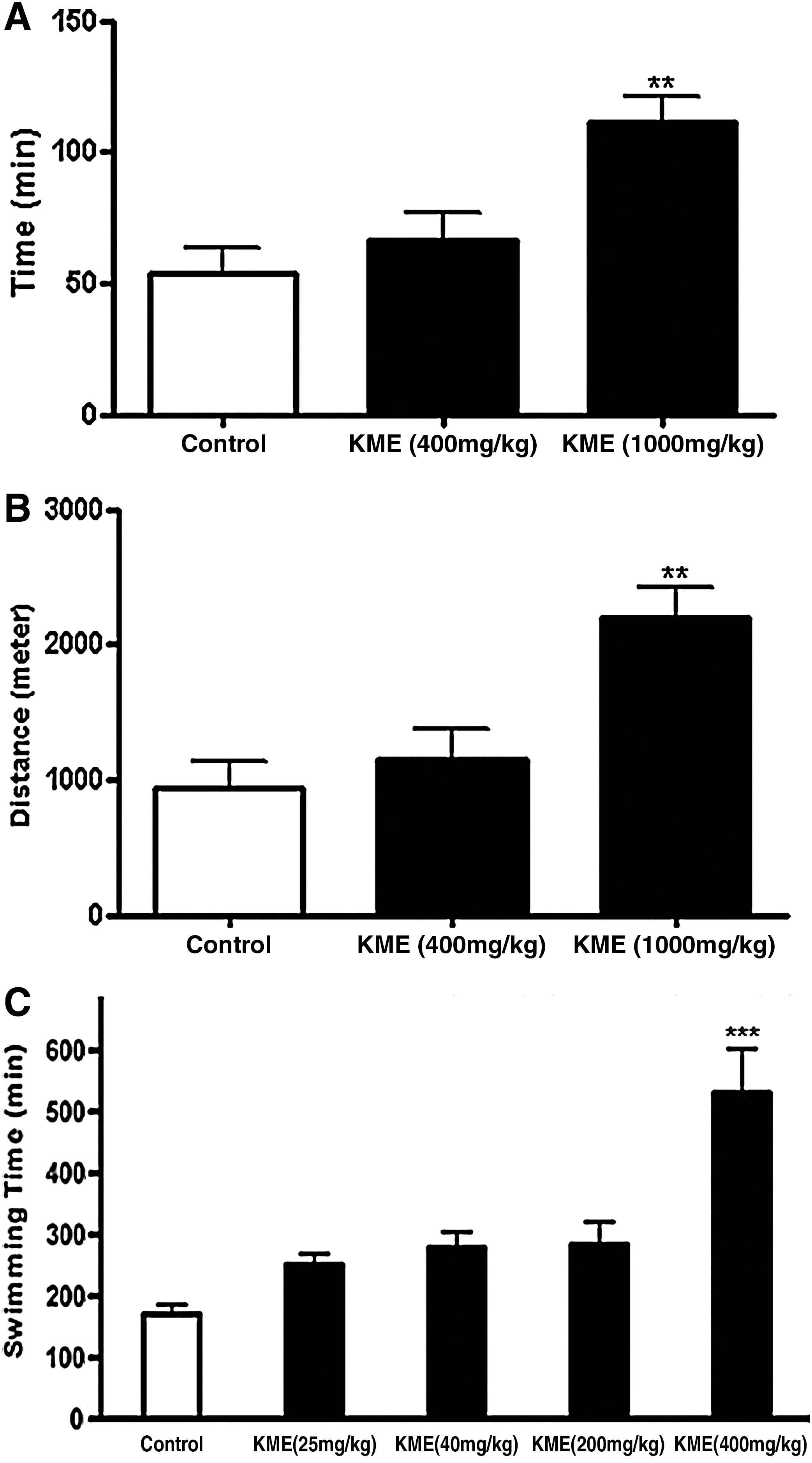
Enhancement of endurance capacity with KME treatment. The effect of KME on endurance was assessed using a treadmill and forced-swim test. The average time
Swimming capacity of mice was increased by oral administration of KME
The tonic effects of KME were measured using the forced-swim test, an accepted method that measures exercise endurance. 29 To further consolidate our hypothesis that KME enhances mitochondrial function, various concentrations of KME at 25, 40, 200, and 400 mg/kg were administered daily to ICR mice for 1 week. As shown in Figure 2C, there were dose-dependent responses in the swimming time to exhaustion between the control group and each treatment group. The swimming time to exhaustion of the control, 25, 40, 200, and 400 mg/kg KME groups were 169.8±16.1 min (100%), 250.5±18 min (148%), 277.8±26.7 min (164%), 283.5±27.0 min (167%), and 522.9±72.4 min (312%), respectively. No differences were observed in average food intake between the control-treated and KME (400 mg/kg)-treated mice (data not shown). Thus, the swimming data and the relevant molecular parameters, such as PGC-1α expression (Fig. 3A), indicate that KME treatment enhanced the forced-swimming capacity of mice by increasing mitochondrial activity and biogenesis. Jung et al. 30 examined the effect of medicinal plant extracts, including Panax ginseng, Grifola frondosa, and Paecilomyces japonica, on the forced-swimming capacity of mice and showed that G. frondosa had the greatest efficacy with a 30% increase in the swimming time to exhaustion compared with the control group. To our knowledge, KME increases the forced-swimming capacity with the greatest potency of any reported agent. Furthermore, the whole-body temperature of mice rapidly declines to the water temperature during swimming, requiring heat production. KME-treated mice might have enhanced thermogenesis, because they were able to swim 2.5× longer than the PBS-fed mice in a glass container, including 20°C to 22°C water. These results were consistent with those in Figure 1, which showed that KME enhanced OCR in conditions of FCCP-induced uncoupled respiration. Therefore, we conclude that the increase in swimming time was caused not only by enhancement of endurance exercise capacity, but also by increased heat production through enhanced mitochondrial function.
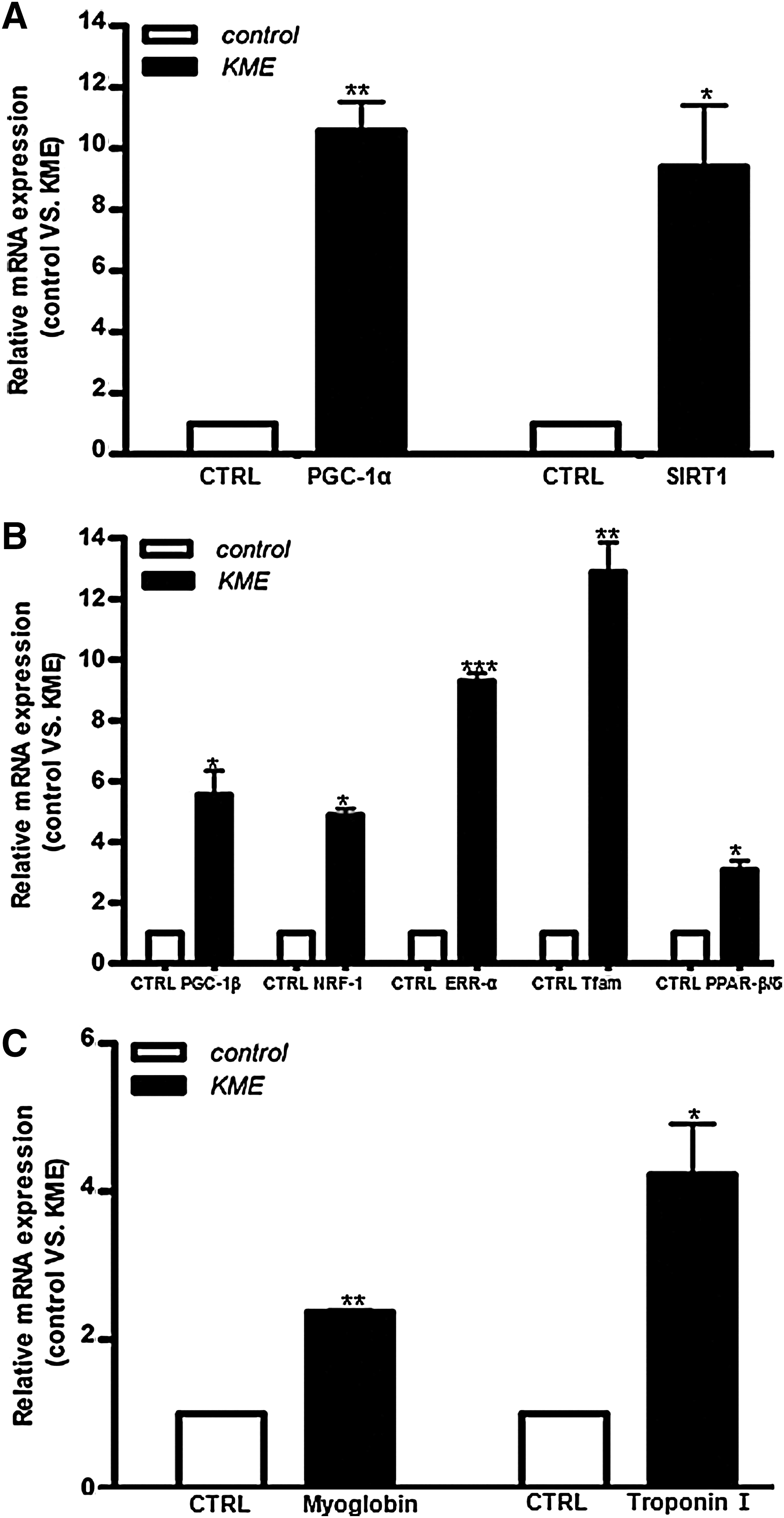
Effects of KME treatment on mRNA expression levels of genes related to mitochondrial biogenesis and function. KME treatment increased SIRT1 and PGC-1α expression
Effects of KME on the level of lactate and glucose in blood after treadmill test
As shown in Table 1, the plasma lactate levels after the treadmill test were significantly lower in the KME 1000 mg/kg group than in the control group (63.2±10.3 vs. 18.1±9.1 mg/dL, P<.001). This result was expected, because lactate levels generally tend to be elevated after exercise, and but lower if KME plays a role in reducing lactate level similar to that observed for silk amino acids (SAAs). 31 Shin et al. 31 showed that swimming exercise increases lactate levels, which was substantially inhibited by SAA treatment. Unfortunately, since the resting level of lactate was not measured before exercise in this study, it is difficult to conclude that the increased level of lactate in the blood was due to exercise and that the depletion was due to KME treatment. However, we believe that the lower level in the KME-treated group compared with the control group is likely due to the KME treatment, because the only difference between the two groups is the presence of KME. It is quite natural to expect that the level of lactate in KME-treated mice should be higher than in the control group, because treated mice ran longer than the untreated control group by 2.5 times. However, the opposite result was obtained. This finding is consistent with that of SAA. 31 It has been shown that the accumulation of lactate leads to a reduction of pH in muscle tissue and blood and may cause muscle fatigue. However, the role of lactic acid looks more complicated, because it is now being recognized as more of a positive player in metabolism. Therefore, we do not favor the interpretation that a low level of lactate induced less muscle fatigue, resulting in longer running in the treadmill test. However, there might be some relationship among the level of lactate, KME treatment, and exercise endurance capacity. To investigate the mechanism related with these findings, more careful studies are required with single leading compounds isolated from KME such as RSV, a natural compound found in grapes and red wine. 32 The lower blood glucose levels could be expected, because there is much evidence that mistletoe extracts have anti-diabetic activity. 5 Recently, it was demonstrated that pentacyclic triterphenoids such as betulinic acid and oleanolic acid, which are major components of KME for anti-cancer activity, 33,34 have anti-diabetic activity via AMPK activation. 35 More interestingly, both betulinic acid and oleanolic acid have been demonstrated as enhancing exercise endurance capacity and extension of the life span of Bombyx mori. 36
Data are means±SD values.
P<.001, b P<.01, significantly different from the control group by one-way analysis of variance and subsequently applying Tukey's test.
KME, Korean mistletoe extract.
Level of glycogen in gastrocnemius muscle and liver
The glycogen content of the liver and gastrocnemius muscle is shown in Table 1. The glycogen content of the liver tended to be lower in KME 1000 mg/kg groups compared with the control group, but not significantly. However, the glycogen content of gastrocnemius muscle was significantly lower in KME 1000 mg/kg groups compared with the control group. These results may be due to the fact that KME-treated mice utilized more glycogen for energy, because they ran longer than the control groups. These results are consistent with the lactate and glucose results. To determine how much the exercise changed the biochemical parameters and how much the KME supplementation reversed the exercise-dependent changes, parameter levels should be measured in a control-sendentary group at rest before exercise and during treatment, a subject currently under investigation. 12
KME treatment increased expression of key genes related to mitochondrial function in KME-treated C2C12 myotube cells
To investigate the molecular basis for the KME-dependent increase in mitochondrial activity, we determined the expression of PGC-1α mRNA, an important factor in mitochondrial biogenesis, 37 and several of its transcriptional targets in C2C12 myotube cells by reverse transcriptase–polymerase chain reaction. We found that PGC-1α mRNA expression significantly increased (10.5-fold) in KME-treated C2C12 myoblast cells when compared with vehicle-treated cells (Fig. 3A). It is well documented that RSV improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1α. 11 Similar to PGC-1α, SIRT1 was shown to increase mitochondrial activity and life span. Therefore, we determined whether KME treatment also affects SIRT1 expression. As shown in Fig. 3A, KME treatment significantly increased SIRT1 expression by 9.3-fold. Our results suggest that KME and RSV may activate the mitochondrial activity by similar mechanisms. SIRT1 is not only involved in mitochondrial activity associated with endurance capacity, but it also has anti-diabetic activity; 38 thus, the effects of KME on SIRT1 expression may underlie its anti-diabetic activity. 38 Recently, we demonstrated that KME treatment extended the life span of the silkworm moth, Bombyx mori, by 30% 36 and increased Sir2 gene expression. These results are consistent with the findings of the present study and further support similar functional roles of KME and RSV. Furthermore, an increase in PGC-1β expression was observed after KME treatment in C2C12 cells (Fig. 3B). PGC-1β functions overlap with those of PGC-1α and can increase the expression of genes related to respiration. 14 In addition to being a target of PGC-1α, the estrogen-related receptor α (ERRα) mediates many of the mitochondrial effects of PGC-1α. 39 KME treatment significantly increased the expression of ERRα (Fig. 3B) and nuclear respiratory factor-1 (NRF-1), a transcriptional target of ERRα/PGC-1. 40 Mitochondrial transcription factor A (Tfam), a nuclear-encoded mitochondrial transcription factor that is essential for the expression of important mitochondrial-encoded genes, 40 is a target of NRF-1, and its expression was increased after KME treatment (Fig. 3B). In addition to these transcription factors, KME affected the expression of an array of PGC-1α transcriptional target genes, 41 including the fiber-type markers myoglobin (MB) and troponin I (TNNI2; Fig. 3C). Research with muscle-specific PGC-1α overexpressing mice has demonstrated the ability of the coactivator to enhance exercise performance, as demonstrated by longer times to exhaustion. 13 Although we did not determine the expression of PGC-1α after excise training compared with a resting sedentary state, there are some reports that PGC-1α mRNA increases several folds after endurance excise training. 13,42 Our results shown in Figure 3 were consistent with the possibility that KME enhances endurance capacity (Fig. 2).
KME does not show any toxicity on the liver and kidney
To determine KME's toxicity, we performed a histological analysis from the liver and kidney of treated mice. As shown in Supplementary Figure S2, the administration of KME did not have any deleterious effects on liver and kidney histology. The serum levels of glutamic oxaloacetic transaminase and glutamic pyruvic transaminase (data not shown) in 1000 mg/kg KME-treated mice were similar to those of control-treated mice, indicating that KME treatment did not induce hepatic toxicity and that KME was well tolerated.
Taken together, our data showed that KME improved mouse mitochondrial activity, resulting in enhanced forced-swimming capacity, forced running time, increased OCR, and expression of key genes related to mitochondrial function. KME supplementation may be beneficial to athletes for improving performance by increasing endurance. Further studies are needed to identify bioactive compound(s) in KME with mitochondrial activating activity.
Footnotes
Acknowledgments
The authors are grateful to Dr. Youngmi Kim Park (Kyung Hee University College of Medicine) for generously allowing them to use the OROBOROS. They are also grateful to Dr. Kyung-Ah Jung (Chuncheon National University of Education) for her kind help. This work was supported by a project on the development of regional industrial technology from the Ministry of Knowledge Economy, Korea.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.