
Abstract
We investigated the anti-obesity effect of abietic acid in mice fed a high-fat diet with emphasis on changes in adipogenesis in epididymal adipose tissues. Male C57BL/6J mice were divided into four groups and fed a normal diet, a high-fat diet (HFD), or HFD plus oral administration of abietic acid (20 mg/kg of body weight/day [LA] or 40 mg/kg of body weight/day [HA]) for 8 weeks. Compared with the HFD group, mice orally administered 40 mg of abietic acid/kg of body weight/day exhibited significantly decreased body weight and adipose tissue weights. Serum triglyceride concentrations in the HA group were significantly lower than those in the HFD group, as were the levels of serum insulin and leptin. Hematoxylin and eosin staining revealed that epididymal adipose tissue mass was decreased by abietic acid administration. Abietic acid also inhibited the protein expression of sterol regulatory element-binding protein-1c, CCAAT/enhancer-binding protein α, and CD36 in epididymal adipose tissues, which are up-regulated by HFDs. These data demonstrate that abietic acid has an anti-obesity effect in mice mediated by the regulation of adipogenesis.
Introduction
O
Adipogenesis is a multifaceted process that results in the development of mature adipocytes from preadipocytes. Sterol regulatory element-binding protein-1c (SREBP-1c) is a transcriptional factor that principally regulates lipid synthesis, and it has also been implicated in adipocyte differentiation. 3 CCAAT/enhancer-binding protein α (C/EBPα) plays an important role in the differentiation of preadipocytes into mature adipocytes via its interaction with peroxisome proliferator-activated receptor γ (PPARγ). Indeed, C/EBPα is associated with lipid metabolism and insulin signaling. 4
Many drugs, including sibutramine and fibrates, are used to reduce body weight gain, but these drugs are known to cause serious side effects. Recently, many plants have been tested to reveal novel treatments for obesity. Abietic acid, a terpenoid, is a phytochemical compound isolated from rosin produced by conifer species. 5,6 Many dietary plants contains terpenoids, which exhibit various beneficial effects in biological systems. 7 A previous study reported that abietic acid inhibited lipoxygenase activity. 8 It has also been reported that abietic acid has anti-inflammatory effects by inhibiting expression of genes involved in inflammation such as those for tumor necrosis factor-α and cyclooxygenase 2 in lipopolysaccharide-induced macrophages. 9 Takahashi et al. 10 demonstrated that abietic acid regulates PPARγ target gene expression in RAW 264.7 macrophages and 3T3L adipocytes. They also suggested that abietic acid can be used for regulating lipid metabolism and atherosclelosis. However, there are no studies regarding the anti-obesity effect of abietic acid in animal models.
The primary aim of the present study was to test whether abietic acid has an anti-obesity effect in vivo. For this purpose, we examined the effect of abietic acid on phenotypic properties of obesity and protein expression involved in adipogenesis in the well-characterized C57BL/6J model of high-fat diet (HFD)-induced obesity.
Materials and Methods
Materials
Abietic acid was purchased from Sigma-Aldrich (St. Louis, MO, USA). An HFD (45% of kcal are fat) was purchased from Research Diets (New Brunswick, NJ, USA). All other chemicals were of analytical grade or purer.
Animals and diets
Four-week-old male mice (C57BL/6J) were purchased from Orient Bio Inc. (Gyeonggi, Korea) and given free access to commercial chow for 7 days. After acclimation, the mice were randomly divided into four groups (n=10) and fed a normal diet (Nor), HFD, HFD + a low dose of abietic acid (LA), or HFD + a high dose of abietic acid (HA) for 8 weeks. The LA and HA groups were orally administered daily a dose of 20 mg or 40 mg of abietic acid/kg of body weight/day, respectively, dissolved in 150 μL of 0.1% hydroxypropyl methylcellulose using oral gavage. The Nor and HFD groups received daily 150 μL of vehicle (0.1% hydroxypropyl methylcellulose) using oral gavage. The composition of the normal diet was based on the 1980 AIN-76 diet of the American Institute of Nutrition. Diets and water were provided ad libitum. The animals were maintained at 21–25°C and a humidity of 50–60% under a 12-hour:12-hour light:dark cycle (lights on from 06:30 hours to 18:30 hours). All animal procedures were conducted in accordance with the Guidelines for Institutional Animal Care and Use Committee of the Korea Food Research Institute (Seongnam, Korea).
Sample preparation
At the end of experimental period, the mice were fasted for 12 hours. After the animal was anesthetized with diethyl ether, blood was obtained from the ophthalmic vein and centrifuged at 1,500 g for 20 minutes to separate the serum, which was stored at −70°C until analysis. The epididymal and perirenal adipose tissues were removed, weighed, snap-frozen in liquid nitrogen, and stored at −70°C until use.
TG, leptin, and insulin analysis
TG levels were measured using a commercial enzyme kit (Shinyang Chemical Co., Busan, Korea). Leptin was analyzed using a mouse leptin immunoassay kit (R&D Systems, Minneapolis, MN, USA). Insulin was analyzed using an insulin enzyme-linked immunosorbent assay kit (Shibayaki, Tokyo, Japan).
Histological analysis of adipose tissue
Epididymal adipose tissue removed from the same part of each mouse was fixed in buffered 4% formaldehyde in phosphate-buffered solution, embedded in paraffin, sliced, and stained with hematoxylin and eosin (Sigma-Aldrich). Morphological changes were assessed and photographed with an Olympus (Tokyo, Japan) BX-51 microscope at ×200 magnification.
Western blot analysis
Epididymal adipose tissue was homogenized in PRO-PREP (Invitrogen, Seoul, Korea) and centrifuged, and then the protein in the supernatant was assayed using a Bradford assay (Bio-Rad, Hercules, CA, USA). Quantified proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a polyvinylidine fluoride membrane (Millipore, Billerica, MA, USA). After proteins were transferred, the membrane was washed in 0.1% Tween containing 10 mM Tris-HCl (pH 7.6) and 150 mM NaCl (TBST) and blocked with 5% skim milk in TBST. Blots were then incubated with primary antibody (anti-SREBP-1c, anti-C/EBPα, anti-CD36, or anti-β-actin [Santa Cruz Biotechnology, Santa Cruz, CA, USA]) overnight at 4°C and then washed with TBST. Blots were incubated with horseradish peroxidase–linked secondary antibody for 2 hours, washed with TBST, and developed by enhanced chemiluminescence.
Statistical analysis
Data are mean±SE values. All statistical analyses were performed by analysis of varaiance and Duncan's multiple-range test using SAS software (SAS Institute, Cary, NC, USA). A value of P<.05 was considered statistically significant.
Results
Body weight, adipose tissue weight, and serum TGs, leptin, and insulin
As shown in Figure 1, body weight in the HFD group was significantly increased compared with that in the Nor group. The LA group showed no significant difference in body weight compared with the HFD group. However, the mice fed HFD supplemented with 40 mg of abietic acid/kg of body weight (HA group) exhibited approximately 27% lower body weight compared with that of the HFD group. No significant difference was observed in daily food intake among the HFD-fed groups. These data indicate that 40 mg of abietic acid exerted anti-obesity effects without affecting food intake in HFD-induced mice. Obesity is characterized by excess body fat and the hypertrophy of adipocytes. Therefore, to evaluate whether the lower body weight in HA mice is accompanied by lessened fat mass, we measured the weight of white adipose tissue and brown adipose tissue. As shown in Table 1, epididymal, perirenal, and brown adipose tissues weights in the HFD group were significantly increased compared with those in the Nor group. However, weights of these adipose tissues were decreased in the HA group by approximately 22%, 22%, and 50%, respectively, compared with those in the HFD group.

Effects of abietic acid on body weight of mice fed experimental diets. Data are mean±SEM values for 10 mice per group. abcValues not sharing a common superscript letter were significantly different at P<.05 between groups. Nor, normal diet; HFD, high-fat diet; LA, HFD-fed mice orally administered low-dose abietic acid (20 mg/kg of body weight/day); HA, HFD-fed mice orally administered high-dose abietic acid (40 mg/kg of body weight/day).
Values for a parameter not sharing a common letter are significantly different (P<.05) between groups.
TGs, triglycerides.
The HFD group exhibited significantly higher serum TG concentrations than the Nor group. However, serum TG concentrations were significantly decreased by the oral administration of abietic acid. Serum leptin and insulin levels in the HA group were significantly lower by approximately 22% and 34%, respectively, compared with those of the HFD group (Table 1).
Adipocyte size in epididymal adipose tissues
Enlarged adipose tissues are one of the general characteristics of obesity. To clarify whether adipocyte size was decreased by HA supplementation, epididymal adipose tissues were stained with hematoxylin and eosin. As shown in Figure 2, the HFD group showed larger size of adipose cells compared with the Nor group. When the mice were administered low-dose abietic acid with HFD (LA group), there was not much difference in cell size compared with the HFD group. However, cell size in adipose tissue in the HA group was dramatically smaller than that of the HFD group and almost the same of the Nor group. These results clearly demonstrate that supplementation of 40 mg of abietic acid with HFD exerts anti-obesity effects by reducing fat mass and adipocyte size in mice.
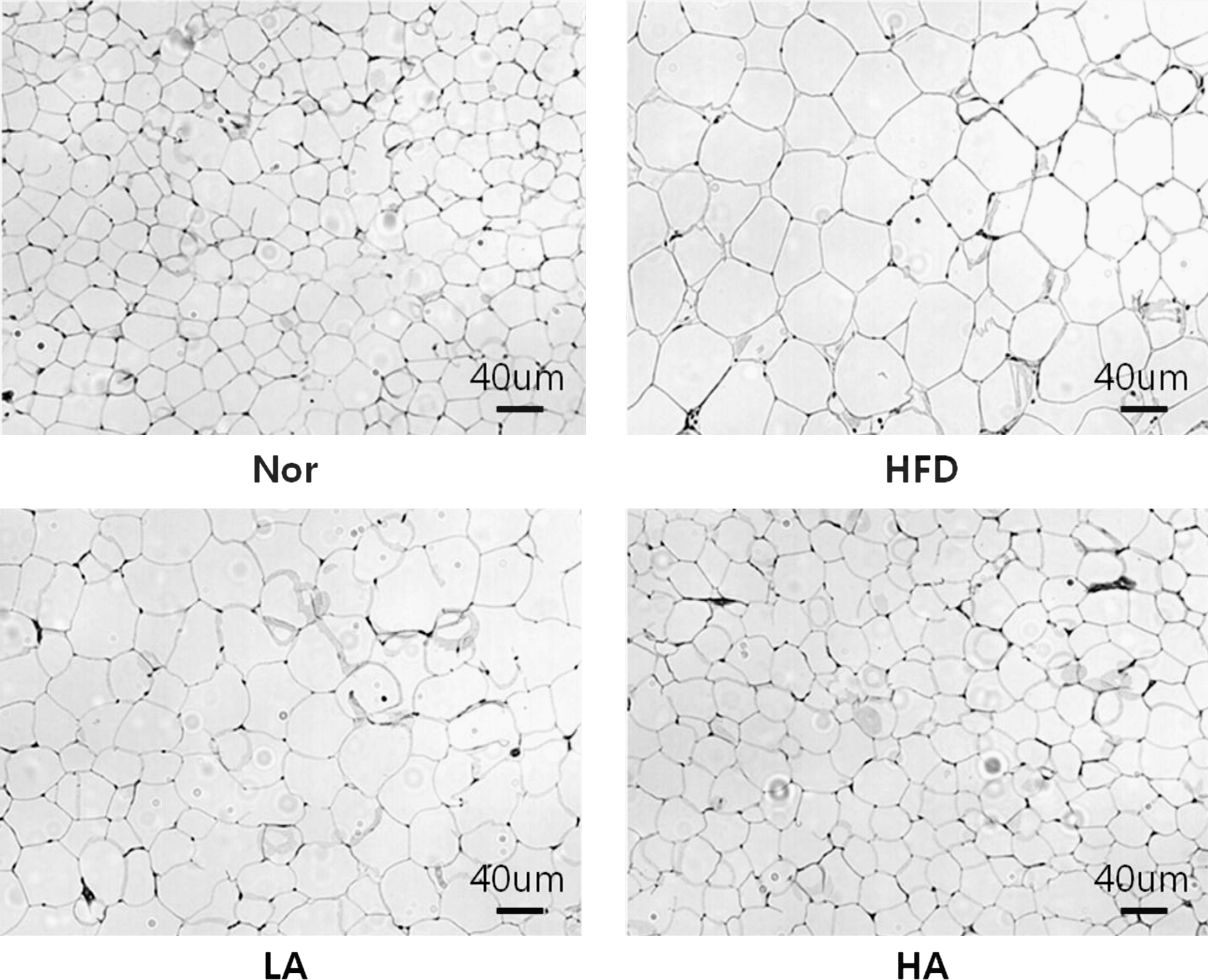
Representative hematoxylin and eosin images of epididymal adipose tissues in experimental animals.
Changes in adipogenic transcriptional factors in epididymal adipose tissues
To clarify the mechanism of action underlying the anti-obesity effects of abietic acid, we investigated changes in the expression of proteins involved in adipogenesis in adipose tissues by western blot analysis. As shown in Figure 3, the protein expression of SREBP-1c and C/EBPα, which are involved in fatty acid synthesis and adipogenesis, in epididymal adipose tissues in the HFD group was higher than that in the Nor group. However, the increased protein expression of SREBP-1c and C/EBPα in the HFD group was attenuated in the HA mice. The protein expression of CD36, which is related in lipid accumulation, was also decreased in the HA group compared with that of the HFD group.

Effects of abietic acid on protein expression of adipogenic markers in epididymal adipose tissues of experimental mice. C/EBPα, CCAAT/enhancer-binding protein α; SREBP-1c, sterol regulatory element-binding protein-1c.
Discussion
In this study, we investigated the anti-obesity effects of abietic acid in mice fed HFD. A prolonged obese state, which is normally caused by a chronic imbalance between energy intake and expenditure, is a major risk factor for various diseases such as diabetes, heart disease, and cancer. Normally, the obese state is characterized by hyperlipidemia, hyperleptinemia, and hyperinsulinemia. 11 Mouse models are optimal for studying general obesity induced by the prolonged consumption of HFDs.
Abietic acid is a major component of the rosin fraction produced by conifer species. 5 It was determined that abietic acid has biological effects that include anticonvulsant 12 and anti-inflammatory 9 properties. However, the anti-obesity effect of abietic acid has not been investigated.
Here, our present study shows that the oral administration of 40 mg of abietic acid/kg of body weight/day for 8 weeks significantly decreased body weight gain in mice fed HFD. It is interesting that daily food intake was not significantly different among the HFD-fed groups. Weight reduction in epididymal and perirenal adipose tissues in the HA group was consistent with the significant decreases in body weight in the HA group compared with those in the HFD group. Taken together, our data clearly demonstrate that abietic acid has anti-obesity effects in diet-induced obese mice.
We found that the abietic acid-administered groups had significantly lower serum TG levels than the HFD group. During the development of obesity, high levels of TGs in the bloodstream lead to enlarged adipose tissues due to intracellular lipid accumulation. Enlarged adipose tissues secrete regulatory factors, termed adipocytokines, which are important in many pathological processes. 13 Leptin is one of the most abundant adipocytokines produced by adipocytes. Leptin plays important roles in regulating energy intake and expenditure. However, excess levels of leptin cause insulin resistance, asthma, and cancers. It has been reported in a previous study that abietic acid has anti-inflammatory effects in RAW 264.7 and 3T3-L1 cells. 10 In this study, we found that animals in the HA groups exhibited significantly decreased levels of serum leptin compared with those of the HFD groups. Furthermore, circulating leptin levels were proportional to the weight of adipose tissue. Insulin is also an important hormone that regulates energy in the body. Hyperinsulinemia increases leptin levels in the bloodstream of rodents and humans. 14 Our data demonstrated that serum insulin levels were significantly decreased by 40 mg of abietic acid/kg of body weight in mice fed HFD.
It is known that insulin stimulates the expression of SREBP-1c in adipose tissues. SREBP-1c is a highly important transcriptional factor in adipocytes that regulates lipogenesis and adipogenesis. 15 In adipocytes, lipogenesis is stimulated by C/EBPα and SREBP-1c, which coordinately induce the expression of fatty acid synthase. Adipocyte differentiation involves an interaction between C/EBPα and PPARr. 4 SREBP-1 enhances the transcriptional activity of PPARγ. Therefore, we evaluated the effect of abietic acid on the protein expression of SREBP-1c and C/EBPα, which are reliable markers for adipogenesis. As shown in Figure 3, abietic acid apparently inhibited the protein expression of SREBP-1c and C/EBPα in epididymal adipose tissues relative to the expression in the HFD-fed groups. These data suggest that abietic acid has anti-obesity effects partly through down-regulating adipogenic transcription factors. CD36 mediates the transfer of fatty acid across the membranes of adipocytes, playing a major role in lipid accumulation. 16 Relative to that of the HFD groups, abietic acid clearly inhibited the protein expression of CD36 in adipocytes.
In conclusion, abietic acid inhibited body weight gain and decreased adipose tissue weight in HFD-fed mice. These effects of abietic acid may be mediated by regulating adipogenic transcriptional factors such as SREBP-1c and C/EBPα. This finding suggests that abietic acid, a naturally occurring compound in dietary plants, may be a useful bioactive compound for preventing diet-induced obesity.
Footnotes
Acknowledgment
This study was supported by the Korea Food Research Institute and Platform Technology Project from the Korea Industrial Technology Foundation.
Author Disclosure Statement
No competing financial interests exist.