
Abstract
Yukmijihwang-tang (YMT) is a traditional herbal medicine known to enhance memory in brain injury models. The aims of this study were to evaluate the inhibitory effect of YMT on osteoclast differentiation and to determine its molecular mechanism of action. YMT dose-dependently inhibited receptor activator for nuclear factor-κB (NF-κB) ligand (RANKL)-induced tartrate-resistant acid phosphatase (TRAP) activity and the formation of multinucleated osteoclasts in RAW264.7 cells. In addition, quantitative reverse transcription–polymerase chain reaction showed that YMT significantly decreased RANKL-induced expression of osteoclast differentiation-specific genes (TRAP, matrix metalloproteinase-9, cathepsin K, and the d2 isoform of vacuolar ATPase V0 domain). Furthermore, YMT inhibited RANKL-induced phosphorylation of mitogen-activated protein kinases (extracellular signal-regulated kinase, c-Jun N-terminal kinase, and p38), phosphorylation of I-κBα, phosphorylation of NF-κB p65, and the expression of transcription factors Fra-2 and nuclear factor of activated T-cells, cytoplasmic 1. Furthermore, YMT inhibited the bone-resorptive activity of differentiated osteoclasts, suggesting that YMT inhibits osteoclast differentiation by suppressing RANKL-induced signaling molecules and transcription factors that affect the regulation of genes for osteoclast differentiation. As such, YMT may have therapeutic potential in bone diseases.
Introduction
B
Traditional herbal medicines exert inhibitory or preventive effects on bone disease models by regulating osteoclast differentiation, which suggests pharmacological potential in bone diseases. 5 Yukmijihwang-tang (YMT), or Liuwei Dihuang, is a traditional herbal medicine consisting of six different herbs: Rehmanniae Radix Preparata, Dioscoreae Rhizoma, Corni Fructus, Alismatis Rhizoma, Hoelen, and Moutan Cortex Radicis. YMT induces gene expression that protects from neuronal degeneration and induces neurite growth, enhancing memory. 6 YMT also ameliorates learning and memory impairment in ibotenic acid–induced amnesia, partly because of its neuroprotective effect in hippocampal cholinergic neuron degeneration. 7 In addition, protective effects of YMT in different injury models, such as cyclophosphamide-induced reproductive toxicity, ischemia/reperfusion-induced renal injury, and N,N′-dimethyl-4,4′-bipyridinium dichloride-induced liver damage, have been reported. 8 –10 Recently, an inhibitory effect of YMT on parathyroid hormone–induced bone resorption and bone loss in ovariectomized rats was suggested. 11 However, no studies have evaluated the YMT inhibition of RANKL-induced osteoclast differentiation and the molecular mechanism affected by YMT.
RAW264.7 cells have been widely used as an in vitro model of osteoclast differentiation and bone resorption. Upon RANKL treatment, RAW264.7 cells differentiate into osteoclast-like cells expressing osteoclast differentiation-specific genes, such as tartrate-resistant acid phosphatase (TRAP) and cathepsin K. 12,13 In addition, MAP kinases and several transcription factors are activated during RANKL-induced osteoclastogenesis in RAW264.7 cells. 14,15 In this study, we investigated the inhibitory effects of YMT on RANKL-induced osteoclast differentiation and the molecular mechanism regulated by YMT in RAW264.7 cells.
Materials and Methods
Preparation of YMT
All six constituents of YMT—Rehmanniae Radix Preparata (800 g), Dioscoreae Rhizoma (400 g), Corni Fructus (400 g), Alismatis Rhizoma (300 g), Hoelen (300 g), and Moutan Cortex Radicis (300 g)—were used in this study. The medicinal herbs were purchased from the Korea Medicine Herbs Association (Yeongcheon, Korea). All voucher specimens were deposited in the herbal bank of the Center for Herbal Medicine Improvement Research, Korea Institute of Oriental Medicine (Daejeon, Korea). YMT was prepared by using a water extraction method (Cosmos-600 extractor, Gyeongseo, Inchon, Korea). The total quantity of medicinal herbs was placed in 25 L of distilled water and then extracted by heating for 3 hours. After extraction, YMT was filtered out using standard testing sieves (pore size, 106 μm) (Retsch, Haan, Germany), lyophilized overnight, and stored at 4°C before use.
High-performance liquid chromatography analysis
All high-performance liquid chromatography (HPLC) reagents were purchased from J.T. Baker (Phillipsburg, NJ, USA). The HPLC system (Dionex, Idstein, Germany) consisted of a pump (LPG 3X00), autosampler (ACC-3000), column oven (TCC-3000SD), and diode array ultraviolet/visible detector [DAD-3000(RS)]. Detector output signal was recorded using a Dionex Chromelon™ chromatography data system. Chromatographic separation was achieved on a Dionex C18 column (particle size, 5 μm; 120 Å; 4.6 mm×150 mm). The ultraviolet wavelength was 230 nm (paoniflorin), 240 nm (loganin), and 280 nm (5-hydroxymethyl-2-furaldehyde and paeonol). The authentic standards (NPC Biotechnology, Seoul, Korea) for HPLC analysis were prepared at concentrations of 30, 100, 200, and 240 μg/mL using 60% methanol, respectively. The mobile phase was consisted of water and methanol with gradient elution at a flow rate of 1.0 mL/minute (Table 1), and column temperature was maintained at 25°C. YMT (10 mg/mL) was syringe-filtered (pore size, 0.45 μm), and 20 μL of YMT was injected for HPLC analysis.
Cell culture and induction of multinucleated osteoclasts
All materials for cell culture were purchased from Gibco (Invitrogen Inc., Grand Island, NY, USA). RAW264.7 cells (TIB-71) (ATCC, Rockville, MD, USA) were maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 mg/mL streptomycin, with a change of medium every 3 days in 5% CO2 at 37°C. For osteoclast differentiation, RAW264.7 cells (1×103 cells per well) were cultured in α-minimal essential medium supplemented with 10% fetal bovine serum and 100 ng/mL RANKL (R&D Systems Inc., Minneapolis, MN, USA). After 4 days, multinucleated osteoclasts were observed.
Cell viability assay
RAW264.7 cells (1×103 cells per well) were plated in a 96-well plate in α-minimal essential medium containing 10% fetal bovine serum. After 24 hours, serially diluted YMT was treated and incubated with the cells for 3 days. Cell viability was then measured with a Cell Counting Kit-8 (Dojindo Molecular Technologies, Rockville, MD, USA) according to the manufacturer's protocol. Three independent experiments were performed in triplicate.
TRAP staining and activity assay
TRAP staining and activity were studied as previously described. 16 In brief, RAW264.7 cells (1×103 cells per well in a 96-well plate) were incubated with RANKL (100 ng/mL) and the indicated concentrations of YMT for 4 days. Multinucleated osteoclasts were fixed in 10% formalin for 10 minutes and ethanol/acetone (1:1 vol/vol) for 1 minute and then stained using the leukocyte acid phosphatase kit (catalog number 387-A, Sigma, St. Louis, MO, USA). The images of TRAP-positive multinucleated cells were captured under a microscope at×100 magnification with a DIXI eXcope 5.0 (DIXI Optics Co. Ltd., Daejeon, Korea). To measure TRAP activity, multinucleated osteoclasts were fixed with 10% formalin for 10 minutes and 95% ethanol for 1 minute and then incubated with 100 μL of citrate buffer (50 mM, pH 4.6) containing 10 mM sodium tartrate and 5 mM p-nitrophenylphosphate (Sigma). After 1 hour of incubation, the enzyme reaction mixtures were transferred into new plates containing an equal volume of 0.1 N NaOH. Absorbance was measured at 410 nm, and TRAP activity was presented as a percentage of control. Three independent experiments were performed in triplicate. Significance was determined with Student's t test. A P value of <.05 was considered statistically significant.
Primer design and real-time quantitative polymerase chain reaction
Primers were designed using an online primer design program. 17 Primer nucleotide sequences were presented in Table 2. For osteoclast differentiation-related gene expression, RAW264.7 cells (2×105 cells per well in a six-well plate) were treated with RANKL for 3 days, YMT was added 1 day after beginning RANKL treatment, and mRNA expression of osteoclast differentiation-related genes was evaluated after the 3 days of treatment. For transcription factor gene expression, cells (2×105 cells per well in a six-well plate) were pretreated with or without YMT for 2 hours and then stimulated with RANKL (100 ng/mL) for 1 day. Total RNA was isolated with the RNeasy mini kit (Qiagen, Valencia, CA, USA) according to the manufacturer's protocol. First-strand cDNA was synthesized with 2 μg of total RNA, 1 μM oligo(dT)18 primer, 10 units of the RNase inhibitor RNasin (Promega, Fitchburg, WI, USA), and Omniscript reverse transcriptase (Qiagen), according to the manufacturer's protocol. Subsequently, SYBR® Green–based quantitative polymerase chain reaction amplification was performed using SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) and the Applied Biosystems 7500 Real-Time PCR System with first-strand cDNA diluted to 1:50 and 20 pmol of primers according to the manufacturer's protocol. The polymerase chain reaction procedure consisted of three segments. The first segment was conducted at 95°C for 10 minutes for the polymerase activation. The second segment consisted of three-step cycling (40 cycles) at 94°C for 40 seconds (denaturation), 55°C for 40 seconds (annealing), and 72°C for 1 minute (extension). The third segment was conducted to generate polymerase chain reaction product temperature dissociation curves (melting curves) at 95°C for 1 minute, 55°C for 30 seconds, and 95°C for 30 seconds. All reactions were performed in triplicate, and data were analyzed using the 2-ΔΔCT method described previously. 18 Glyceraldehyde 3-phosphate dehydrogenase was used as the internal standard for quantitative polymerase chain reaction. Significance was determined with Student's t test using glyceraldehyde 3-phosphate dehydrogenase–normalized 2-ΔΔCT values. A P value of <.05 was considered statistically significant.
ATP6v0d2, d2 isoform of vacuolar ATPase V0 domain; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; MMP-9, matrix metalloproteinase 9; NFATc1, nuclear factor of activated T-cells, cytoplasmic 1; TRAP, tartrate-resistant acid phosphatase.
Western blot analysis
RAW264.7 cells (2×105 cells per well) were plated in a six-well plate, pretreated with or without YMT for 2 hours, and then stimulated with RANKL (100 ng/mL) for 0, 5, 15, and 30 minutes. Cells were homogenized in an ice-cold protein extraction buffer consisting of 50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1% Nonidet P-40, 0.1% sodium dodecyl sulfate, 1 mM phenylmethylsulfonyl fluoride, one protease inhibitor cocktail tablet, and one phosphatase inhibitor cocktail tablet (Roche, Mannheim, Germany). Whole-cell lysates were centrifuged at 10,000 g for 15 minutes at 4°C. Protein concentration was determined with the BCA protein assay kit (Pierce, Rockford, IL, USA). Protein samples (10 μg) from whole-cell lysates were mixed with sample buffer (100 mM Tris-HCl, 2% sodium dodecyl sulfate, 1% 2-mercaptoethanol, 2% glycerol, and 0.01% bromophenol blue, pH 7.6), incubated at 95°C for 5 minutes, and loaded onto 10% polyacrylamide gels. Electrophoresis was performed using a Mini Protean 3 cell (Bio-Rad, Hercules, CA, USA). Proteins separated on the gels were transferred onto a nitrocellulose membrane (Whatman, Dassel, Germany) and stained with Ponceau S staining solution to ascertain protein loading quantity and transfer efficiency. The stained membrane was washed and incubated in blocking buffer (10 mM Tris-HCl [pH 7.5], 150 mM NaCl, 0.1% Tween-20, and 3% nonfat dry milk). The membrane was then incubated for 2 hours at room temperature with 1:1,000 diluted primary antibodies. Antibodies specific for phosphorylated ERK1/2 (Thr202/Tyr204), ERK, phosphorylated JNK1/2 (Thr183/Tyr185), JNK, phosphorylated p38 (Thr180/Tyr182), p38, phosphorylated I-κBα (Ser32/36), I-κBα, phosphorylated NF-κB p65 (Ser536), and NF-κB p65 were from Cell Signaling Technology (Danvers, MA, USA). Antibody for β-actin was from Abcam (Cambridge, MA, USA). After three washes with blocking buffer for 10 minutes each, the membrane was probed with 1:5,000 diluted secondary antibodies (Cell Signaling Technology) for 1 hour. The membrane was then washed three times for 10 minutes each and developed with SuperSignal® West Femto Maximum Sensitivity Substrate (Pierce). Chemiluminescent signal was detected with an LAS-3000 luminescent image analyzer (Fuji Photo Film Co., Tokyo, Japan). Band densities were measured with Multi Gauge software version 3.0 (Fuji Photo Film Co.).
Pit formation assay
RAW264.7 cells (1×103 cells per well) were seeded onto calcium phosphate apatite–coated plates (BioCoat™ Osteologic™ multitest slides, BD Biosciences, Bedford, MA, USA) with α-minimal essential medium containing 10% fetal bovine serum and 100 ng/mL RANKL. YMT was treated with RANKL every 3 days from differentiation day 6, at which time the mature osteoclasts were almost functionally activated. At 10 days of incubation, the slides were washed with 6% sodium hypochlorite solution to remove cells. The resorbed areas on the slides were observed with a microscope at×100 magnification.
Results
HPLC analysis of YMT constituents
HPLC analysis simultaneously identified four marker components of YMT: paeoniflorin and paeonol of Moutan Cortex Radicis, loganin of Corni Fructus, and 5-hydroxymethyl-2-furaldehyde of Rehmanniae Radix Preparata. The four components were identified in the same retention time window as studied previously (data not shown). 19 The concentrations of paeoniflorin, paeonol, loganin, and 5-hydroxymethyl-2-furaldehyde in YMT were 0.67, 3.86, 0.67, and 3.89 μg/mg, respectively.
YMT inhibited RANKL-induced osteoclastogenesis in RAW264.7 cells without cytotoxicity
We evaluated the effect of YMT on RAW264.7 cell proliferation to exclude potential cytotoxic effects exerted by YMT. At the concentrations used (50–400 μg/mL), YMT did not affect cell proliferation (Fig. 1A). To investigate the effect of YMT on osteoclast differentiation in RAW264.7 cells, TRAP activity and multinucleated osteoclast formation were evaluated after 3 days of YMT treatment. RANKL treatment induced the formation of TRAP-positive multinucleated osteoclasts in RAW264.7 cells. However, YMT significantly inhibited RANKL-induced TRAP activity (Fig. 1B) and the formation of multinucleated osteoclasts in a dose-dependent manner (Fig. 1C).

Effects of Yukmijihwang-tang (YMT) on (
YMT inhibited RANKL-induced expression of osteoclast differentiation-specific genes
TRAP, c-Src, matrix metalloproteinase-9 (MMP-9), cathepsin K, and the d2 isoform of vacuolar ATPase V0 domain (ATP6v0d2) are necessary for osteoclast differentiation and bone-resorptive function. Therefore, we investigated the inhibitory effects of YMT on osteoclast differentiation-specific gene expression in RAW264.7 cells with real-time quantitative polymerase chain reaction. RANKL treatment dramatically up-regulated the mRNA expression of genes encoding TRAP, c-Src, MMP-9, cathepsin K, and ATP6v0d2 in RAW264.7 cells, but YMT (400 μg/mL) significantly down-regulated the RANKL-induced expression of TRAP, MMP-9, cathepsin K, and ATP6v0d2 mRNA (P<.05) (Table 3).
The mRNA expression was analyzed by quantitative polymerase chain reaction. For osteoclast differentiation-related gene expression, cells were treated with RANKL for 3 days, adding YMT 1 day after beginning RANKL treatment; mRNA expression of osteoclast differentiation-related genes was evaluated after the 3 days of treatment. Data are mean±SD values of triplicate experiments.
P<0.05 compared with the group given RANKL.
YMT inhibited RANKL-induced MAP kinase activation
RANKL binding to its receptor, RANK, activates MAP kinase (ERK, JNK, and p38) signaling pathways, transferring RANKL signaling to the transcription factor for osteoclast differentiation. To elucidate YMT's molecular mechanism of action, we evaluated the inhibitory effect of YMT on MAP kinase activation in RANKL-induced osteoclast differentiation. We analyzed these activities by comparing the phosphorylated form to the nonphosphorylated form in western blot analysis of whole-cell extracts from RAW264.7 cells. RANKL treatment evidently induced MAP kinase phosphorylation, but pretreatment with YMT inhibited RANKL-induced MAP kinase phosphorylation in a dose-dependent manner (Fig. 2).

Effects of YMT on RANKL-induced mitogen-activated protein kinase activation: extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38. RAW264.7 cells (2×105 cells per well) were plated in a six-well plate and pretreated with or without YMT (400 μg/mL) for 2 hours and then stimulated with RANKL (100 ng/mL) for 30 minutes. Western blot analysis was performed using whole-cell lysate (10 μg of protein) and an antibody specific for each mitogen-activated protein kinase. The densities of phosphorylated protein (p-) levels (upper panels) were normalized to the density of nonphosphorylated protein levels (lower panels). β-Actin was used as a loading control.
YMT inhibited RANKL-induced phosphorylation of I-κBα and NF-κBp65
RANKL significantly induced I-κBα and NF-κB p65 phosphorylation in RAW264.7 cells after 5 minutes of RANKL treatment. YMT slightly inhibited I-κBα and NF-κB p65 phosphorylation after 5 minutes of RANKL stimulation. However, YMT markedly inhibited NF-κB p65 phosphorylation roughly by half at 15 minutes (Fig. 3).
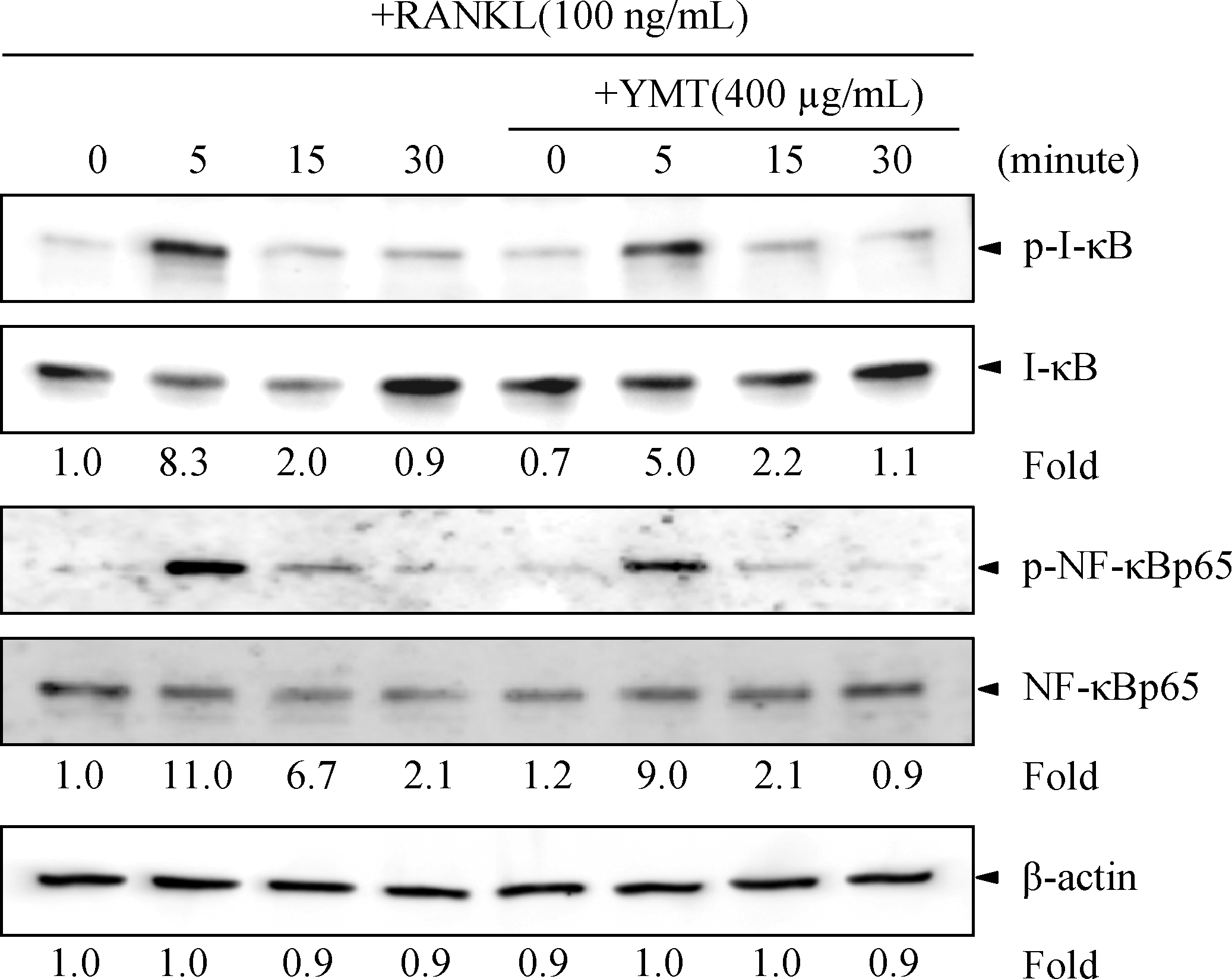
Effects of YMT on RANKL-induced I-κBα and nuclear factor-κB (NF-κB) p65 phosphorylation. RAW264.7 cells (2×105 cells per well) were plated in a six-well plate and pretreated with or without YMT (400 μg/mL) for 2 hours and stimulated with RANKL (100 ng/mL) for the time indicated. Western blot analysis was performed using whole-cell lysates (10 μg of protein) and an antibody specific for I-κBα or NF-κBp65. The densities of phosphorylated protein (p-) levels (upper panels) were normalized to the density of nonphosphorylated protein levels (lower panels). β-Actin was used as a loading control.
YMT inhibited RANKL-induced activator protein-1 and nuclear factor of activated T-cells, cytoplasmic 1 mRNA expression
Activator protein-1 (AP-1) and nuclear factor of activated T-cells (NFAT), cytoplasmic 1 (NFATc1) are key transcription factors that regulate gene expression in RANKL-induced osteoclast differentiation. Therefore, the inhibitory effects of YMT on the expression of mRNA for four transcription factors (c-Fos, Fra-1, Fra-2, and NFATc1) in RAW264.7 cells was further investigated by real-time quantitative polymerase chain reaction. The mRNA expression was significantly up-regulated by RANKL, but pretreatment with YMT (400 μg/mL) significantly down-regulated the expression of Fra-2 and NFATc1 mRNAs (P<.05) (Table 4).
The mRNA expression was analyzed by quantitative polymerase chain reaction. For transcription factor gene expression, cells were treated with YMT 2 hours before RANKL treatment, and mRNA expression was evaluated 1 day after RANKL treatment. Data are mean±SD values of triplicate experiments.
P<.05 compared with the group given RANKL.
NFATc1, nuclear factor of activated T-cells, cytoplasmic 1.
YMT inhibited RANKL-induced pit formation
To determine the inhibitory effect of YMT on the bone-resorptive function of osteoclasts, we performed a pit formation assay. RAW264.7 cells seeded on calcium phosphate apatite–coated plates were incubated with RANKL for 6 days and then treated with YMT for an additional 10 days. YMT dose-dependently decreased RANKL-induced pit formation (Fig. 4).

Effects of YMT on RANKL-induced pit formation. RAW264.7 cells (1×103 cells per well) were seeded onto calcium phosphate apatite–coated plates with α-minimal essential medium containing 10% fetal bovine serum and 100 ng/mL RANKL. YMT was treated with RANKL every 3 days from differentiation day 6, at which time the mature osteoclasts were almost functionally activated. After 10 days of incubation, cells were lysed, and images were obtained with a microscope at×100 magnification.
Discussion
Individual quantitative analysis of major components of each herb is used as a quality control measure for herbal medicine. However, because herbal medicines contain several components of a multitude of herbs, individual analysis of major components is time-consuming, and efficient quality control and standardization are difficult. Therefore, we needed to develop a quality control method with simultaneous determination of different components in herbal medicine, simultaneously determining four components in YMT. HPLC analysis separated four marker components without affecting each component, suggesting that this analysis is accurate and reliable for assessing the quality of YMT.
Interaction between RANKL and RANK initiates recruitment of tumor necrosis factor receptor–associated factor family proteins, which sequentially activate several downstream signaling pathways. 20 MAP kinases are proline-directed serine/threonine kinases that transmit RANKL signals to transcription factors for the induction of osteoclastogenesis-specific genes in osteoclast differentiation. 21 –23 The primary target of MAP kinase signaling is AP-1, formed by a transcriptional complex with Jun (c-Jun, JunB, and JunD) and Fos proteins (c-Fos, FosB, Fra-1, and Fra-2). JNK phosphorylates c-Jun and Jun-D, which are necessary for AP-1 transcriptional activity, whereas p38 induces c-fos expression and transcriptional activity during RANKL-induced osteoclastogenesis. 24,25 RANKL-stimulated AP-1 selectively initiates the transcription of NFATc1, which cooperatively forms a transcription complex with AP-1 to induce osteoclast-specific genes in the early phase of osteoclastogenesis. 26,27 The essential role of NFATc1 and c-Fos in osteoclastogenesis has been shown in NFATc1 and c-fos knockout mice that are severely osteopetrotic. 28 It is interesting that pharmacological inhibition or inactivation of p38 using dominant-negative MAP kinase kinase-6 decreases the induction of NFATc1 mRNA in osteoclasts. 29 In addition, p38 phosphorylates NFATc1, resulting in the nuclear accumulation required for maximum NFATc1 transcriptional activity. 30 Furthermore, JNK and p38 promote NFAT nuclear export by selectively phosphorylating NFAT proteins. 31 Among Fos proteins, Fra-1 compensates for c-Fos deficiency, whereas the ability of Fra-2 to compensate for c-Fos deficiency is relatively weak. 32,33 Therefore, the inhibitory effect of YMT on MAP kinase activation and NFATc1 expression suggests that NFAT might be a target of the YMT inhibitory effect on osteoclast differentiation.
NF-κB signaling is essential for the regulation of RANKL-induced osteoclast differentiation into mature osteoclasts. In the canonical NF-κB pathway, phosphorylation and subsequent degradation of the inhibitory subunit, I-κB, are followed by translocation of the NF-κB subunits (mostly RelA-p50) into the nucleus for DNA binding and gene transcription. 34 NF-κB cooperates with NFAT, cytoplasmic 2 to initially trigger the induction of NFATc1, resulting in auto-amplification of NFATc1 in osteoclastogenesis. 26 An NF-κB inhibitor, (–)-dehydroxymethylepoxyquinomicin, inhibits osteoclastogenesis and bone-resorptive activity by down-regulating RANKL-induced NFATc1 expression. 35 It is interesting that RANKL-induced NF-κB p65 phosphorylation via the p38 pathway increases the transactivation activity of NF-κB involved in NFATc1 induction in osteoclastogenesis. 29 Cells expressing mutant NF-κB p65 that lacks phosphorylation show direct repression of some NF-κB target genes and repression of non–NF-κB-regulated genes through epigenetic mechanisms. 36 We found that YMT inhibits RANKL-induced phosphorylation of I-κBα and NF-κB p65. Therefore, the results suggest that YMT might inhibit several steps of NF-κB signaling, affecting the transcriptional activity of NF-κB in the early phase of osteoclast differentiation.
RANKL-induced activation of NFATc1 and AP-1 directly regulates genes for osteoclast differentiation-specific proteins (TRAP, cathepsin K, MMP-9, and ATP6v0d2) through multiple NFAT and AP-1 binding sites on their promoters. 37,38 During osteoclast differentiation, the expression of these osteoclast-specific genes is increased. Inactivation of NFAT by small interfering RNA or a pharmacological inhibitor significantly attenuates RANKL-induced up-regulation of the expressions of these genes or their activities. 39 –41 TRAP, cathepsin K, MMP-9, and ATP6v0d2 are also known to participate in the bone-resorptive function of osteoclasts. 42 –45 We found that YMT suppressed the expression of these osteoclast differentiation-specific genes (Table 3) and bone resorption (Fig. 4). The findings suggest that the inhibitory effects of YMT on the expression of osteoclast differentiation-specific genes might prevent RANKL-induced bone resorption as well.
Diarylheptanoids or water extract from Dioscoreae Rhizoma, which makes up part of YMT, inhibits the formation of osteoclasts or bone resorption via inhibition of the generation of functional cathepsin K. 46,47 In addition, YMT inhibited parathyroid hormone–induced bone resorption in bone organ culture and partially prevented bone loss in ovariectomized rats as a result of the inhibition of YMT on src tyrosine kinase activity in osteoclasts. 11 Supporting an inhibitory activity of YMT on bone resorption, we found an inhibitory effect of YMT on the activation of MAP kinase signaling and NF-κB signaling, the expression of transcription factors (Fra-2 and NFATc1), and genes for osteoclast differentiation-specific proteins like cathepsin K, which might be part of the molecular mechanisms underlying the inhibitory activity of YMT on osteoclast differentiation and bone resorption. Considering that herbal medicines generally contain complex chemical constituents from different herbs, several components within YMT might affect different molecular mechanisms that work together to reduce osteoclast differentiation and bone loss.
In conclusion, this study demonstrated that YMT suppresses osteoclast differentiation by inhibiting MAP kinases, NF-κB p65 activation, and the expression of NFATc1 and Fra-2, consequently decreasing the expression of osteoclast differentiation-specific genes. This study provides a molecular basis for YMT regulation of osteoclast differentiation, which suggests that YMT has therapeutic potential in the prevention of bone diseases. Further study is needed to identify the inhibitory component in YMT for osteoclast differentiation.
Footnotes
Acknowledgment
This work was supported by a grant (K11050) from the Korea Institute of Oriental Medicine, Ministry of Education, Science and Technology, Korea.
Author Disclosure Statement
No competing financial interests exist.