
Abstract
The present study evaluated the hepatoprotective effect of aqueous ethanolic Moringa oleifera leaf extract (MoLE) against radiation-induced oxidative stress, which is assessed in terms of inflammation and lipid peroxidation. Swiss albino mice were administered MoLE (300 mg/kg of body weight) for 15 consecutive days before exposing them to a single dose of 5 Gy of 60Co γ-irradiation. Mice were sacrificed at 4 hours after irradiation. Liver was collected for immunoblotting and biochemical tests for the detection of markers of hepatic oxidative stress. Nuclear translocation of nuclear factor kappa B (NF-κB) and lipid peroxidation were augmented, whereas the superoxide dismutase (SOD), catalase (CAT), reduced glutathione (GSH), and ferric reducing antioxidant power (FRAP) values were decreased by radiation exposure. Translocation of NF-κB from cytoplasm to nucleus and lipid peroxidation were found to be inhibited, whereas increases in SOD, CAT, GSH, and FRAP were observed in the mice treated with MoLE prior to irradiation. Therefore pretreatment with MoLE protected against γ-radiation-induced liver damage. The protection may be attributed to the free radical scavenging activity of MoLE, through which it can ameliorate radiation-induced oxidative stress.
Introduction
H
Recently, interest in the radioprotective properties of plants and plant compounds has escalated owing to their natural origin, cost-effectiveness, and fewer side effects. One such plant, Moringa oleifera Lam. (synonym Moringa pterygosperma Gaertn., Family Moringaceae) is considered one of the world's most useful trees among all the medicinal plants, as almost every part of the tree can be used for food or other beneficial purposes.
M. oleifera leaves have attracted worldwide research interest because they are good source of natural antioxidants such as ascorbic acid, flavonoids, phenolics, and carotenoids. The high concentrations of ascorbic acid, β-sitosterol, iron, calcium, phosphorus, copper, vitamins A, B, and C, α-tocopherol, riboflavin, nicotinic acid, folic acid, pyridoxine, β-carotene, protein, and essential amino acids (methionine, cysteine, tryptophan, and lysine) present in Moringa leaves make a valuable dietary supplement. 1 It also has hepatoprotective activity. 2,3 Moringa shows radioprotective potential as the leaf extract protects bone marrow chromosomes from radiation-induced damage. 4
The liver is a highly metabolically active organ; thus it should reflect any systemic derangement on ionizing radiations. After radiation exposure the possibility of liver cell regeneration is very low, and the detoxification function of the liver is affected significantly. Therefore, we searched for a protective agent that can provide protection against radiation-induced hepatic alterations from a mechanistic viewpoint. The current article also validates the radioprotective properties of a commonly used vegetable at the cellular and nuclear signaling levels. The present study is the first of its kind to investigate the radioprotective efficacy of Moringa leaf against γ-radiation-induced inflammation and lipid peroxidation (LPO) in the mouse liver.
Materials and Methods
Moringa extract
The leaves of M. oleifera were collected from a specific tree in the month of September of 2008 and certified by the Botanical Survey of India (voucher number CNH/I-I/(310)/2009/Tech.II/352). The leaves were air-dried, powdered, and extracted with 50% ethanol. The extract obtained was vacuum-evaporated to produce a powdered form and was dissolved in double-distilled water with the help of a cyclomixer just before oral administration. This M. oleifera leaf extract has been termed MoLE.
Free radical scavenging activity of MoLE
Free radical scavenging activity of MoLE was determined using the 2,2-di-(4-tert-octylphenyl)-1-picrylhydrazyl (DPPH) and 2,2′-azinobis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) assays.
The ability to scavenge stable free radical DPPH• was measured by the method of Oktay et al.
5
Three milliliters of MoLE (10 mg/mL) was added to 1 mL of 0.2 mM ethanolic DPPH solution. This set-up was kept for 30 minutes, and optical density (OD) was recorded at 517 nm. The control consisted of an equal amount of ethanolic DPPH but without leaf extract. Radical scavenging activity was determined from the following equation:
The ABTS radical cation scavenging activity of the extract was determined according to the method of Re et al. 6 In brief, 7 mM ABTS was reacted with 140 mM potassium persulfate overnight in the dark to yield the ABTS• +. Prior to use in assay, ABTS• + was diluted with 50% ethanol for an initial absorbance at 734 nm at 30°C. Free radical scavenging activity was assayed by mixing 1 mL of diluted ABTS• + with 20 μL of leaf extract and monitoring the change in absorbance (734 nm) at zero-time and 1-, 2-, 3-, 4-, and 5-minute intervals along with immediate reading until a steady state was achieved. Ascorbic acid was used as the positive control.
Mice
Swiss albino male mice (Mus musculus), 6–8 weeks old with body weights of 24±2 g, were obtained from Bengal Chemical and Pharmaceuticals Ltd. (Kolkata, India). Mice were maintained according to the guidelines set by the Institutional Animal Ethical Committee of India, maintained under controlled temperature (23±2°C) and humidity (50±5%) conditions and a 12-hour light–dark cycle. Animals were given standard mouse feed (obtained from Hindustan Lever Ltd., Mumbai, India) and water ad libitum.
Irradiation
Mice were irradiated with a 60Co source at the Saha Institute of Nuclear Physics, Kolkata. Unanesthetized animals were restrained in well-ventilated acrylic boxes and exposed to whole-body γ-radiation (5 Gy), at a dose rate of 1 Gy/minute and a source-to-surface distance of 77.5 cm.
Determination of optimum dose of MoLE
Mice were divided into five groups of six animals each and were administered MoLE orally (75, 100, 150, 300, or 450 mg/kg of body weight) for 15 days. One hour after the last administration, animals were exposed to whole-body 5-Gy γ-radiation. Animals were observed for 30 days for signs of radiation sickness. All MoLE-treated mice showed an increase in body weight and reduced levels of radiation sickness such as vomiting and ruffling of hair compared with the irradiated animals. Among all the doses, 300 mg/kg of body weight was determined as the optimum dose from the perspective of amelioration of radiation sickness. This optimum dose (300 mg/kg of body weight) was used for the experiment.
Experimental design
Mice were selected from an inbred colony and divided into four groups of eight animals each: control group, mice were given distilled water through oral gavages once a day for 15 consecutive days; IR, mice were given distilled water for 15 days and then exposed to 5 Gy of γ-radiation; LE, mice were treated with MoLE at 300 mg/kg of body weight through oral gavages for 15 consecutive days; and LE+IR, MoLE was given at 300 mg/kg of body weight for 15 days, and 1 hour after the last dose they were exposed to a 5-Gy dose of γ-radiation.
All the animals were sacrificed by cervical dislocation at 4 hours post-irradiation. Liver was removed for the estimation of nuclear translocation of nuclear factor κB (NF-κB), LPO, superoxide dismutase (SOD) activity, catalase (CAT) activity, reduced glutathione (GSH) level, and ferric reducing antioxidant power (FRAP).
Western blot assay
Liver was sonicated using tissue homogenization buffer (50 mM Tris-HCl [pH 8], 10 mM KCl, 1 mM EDTA disodium salt, 0.2% nonyl phenoxylpolyethoxylethanol, 10% glycerol, and 1 μg/mL each leupeptin and aprotinin in distilled water) and centrifuged. Sixty microliters of cold nuclear extraction buffer (20 mM HEPES, 10 mM KCl, 1 mM EDTA, 20% glycerol, 10 mM NaCl, 1 μg/mL each leupeptin and aprotinin, and 1 mM phenylmethylsulfonyl fluoride in distilled water) was added to the pellet, vortex-mixed vigorously, and centrifuged to collect the supernatant containing nuclear proteins. Protein concentration was determined by the method of Lowry et al. 7 Equal amounts of protein (50 μg) in each lane were subjected to 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane. The membrane was blocked with 3% bovine serum albumin solution and kept overnight at 4°C. Immunoblotting was done as described previously by Ghose et al. 8 using monoclonal antibody to mouse NF-κB (p65) (Imgenex, San Diego, CA, USA). Immunoblots were analyzed using a model GS-700 imaging densitometer and Molecular Analyst version 1.5 software (Bio-Rad Laboratories, Hercules, CA, USA).
Biochemical estimation
The livers were homogenized using Tris buffer (pH 7.4) for biochemical estimation. The thiobarbituric acid–reactive substances (TBARS) level in the homogenate was estimated by using a standard protocol. 9 In brief, the homogenate was incubated with 15% trichloroacetic acid, 0.375% thiobarbituric acid, and 5 N HCl at 95°C for 15 minutes, the mixture was cooled and centrifuged, and the absorbance of the supernatant was measured at 535 nm against the appropriate blank. The amount of LPO was determined by using ɛ=1.56×105 M –1 cm–1 and expressed as amount of TBARS produced in nanomoles per gram of tissue.
SOD activity was determined using the pyrogallol autooxidation method. 10 In brief, liver homogenate was added to 62.5 mM Tris-cacodylic acid buffer, containing 1 mM diethylenetriaminepentaacetic acid, followed by addition of 4 mM pyrogallol. The autooxidation of pyrogallol was monitored at 420 nm.
CAT activity was measured by monitoring the decrease in absorbance resulting from the elimination of H2O2 by the action of CAT. 11 The standard reaction mixture contained 50 mM potassium phosphate buffer (pH 7.0), 30 mM hydrogen peroxide, and 3 μL of liver homogenate for a total volume of 1.0 mL. The reaction was run at 20°C, and only the initial linear rate was used to estimate the CAT activity. The enzyme activity was determined from its ability to decompose 1 μmol of H2O2/minute and taken as 1 U of activity using the ɛ for H2O2 at 240 nm (i.e., 43.6 M –1cm–1).
GSH was determined according to the method described by Moron et al. 12 Liver homogenate was treated with 0.1 mL of 25% trichloroacetic acid, and the resulting precipitate was pelleted by centrifugation at 3,900 g for 10 minutes. Free endogenous sulfhydryl was assayed in a total volume of 3 mL by adding 2 mL of 0.5 mM 5,5′-dithio-bis(2-nitrobenzoic acid) prepared in 0.2 M phosphate buffer (pH 8) to 1 mL of the supernatant. The GSH reacts with 5,5′-dithio-bis(2-nitrobenzoic acid) to form a yellow complex. The absorbance was read at 412 nm.
The FRAP level of liver homogenate was determined using a standard method. 13 In brief, 1 mL of FRAP reagent (300 mM acetate buffer, 10 mM 2,4,6-tripyridyl-s-triazine solution, and 20 mM FeCl3·6H2O solution in a volume ratio of 10:1:1) and 10 μL of liver homogenate were mixed and kept in a water bath at 37°C for 4 minutes. The optical density was measured at 593 nm. Concentration was calculated against a FeSO4 standard curve. One FRAP unit is equal to 100 μmol/dm3 Fe2+. 14
Statistical analysis
Data are expressed as mean±SEM values. One-way analysis of variance with Tukey's post hoc test was used for statistical evaluation of the data and for the determination of level of significance in various groups of animals. In all cases, a value of P<.05 was considered significant.
Results
Free radical scavenging activity of MoLE
DPPH and ABTS+ radical scavenging activities of MoLE were compared with those of the positive control, ascorbic acid. The concentration of MoLE required to scavenge 50% of the DPPH• was 40 mg/mL, and that for ascorbic acid was 4.17 mg/mL.
The concentration of MoLE required to scavenge 50% of the ABTS• + was 0.258 μg/mL, and that for ascorbic acid was 0.188 μg/mL.
Inhibition of radiation-induced NF-κB (p65) activation by MoLE
Maximum nuclear transport of NF-κB (p65) as observed in radiation-exposed mice was lessened in LE+IR animals. The NF-κB levels in nuclear fraction increased 36% in the IR group compared with control mice. In the LE+IR group nuclear translocation of p65 was down-regulated by 13% compared with the IR group (Fig. 1).
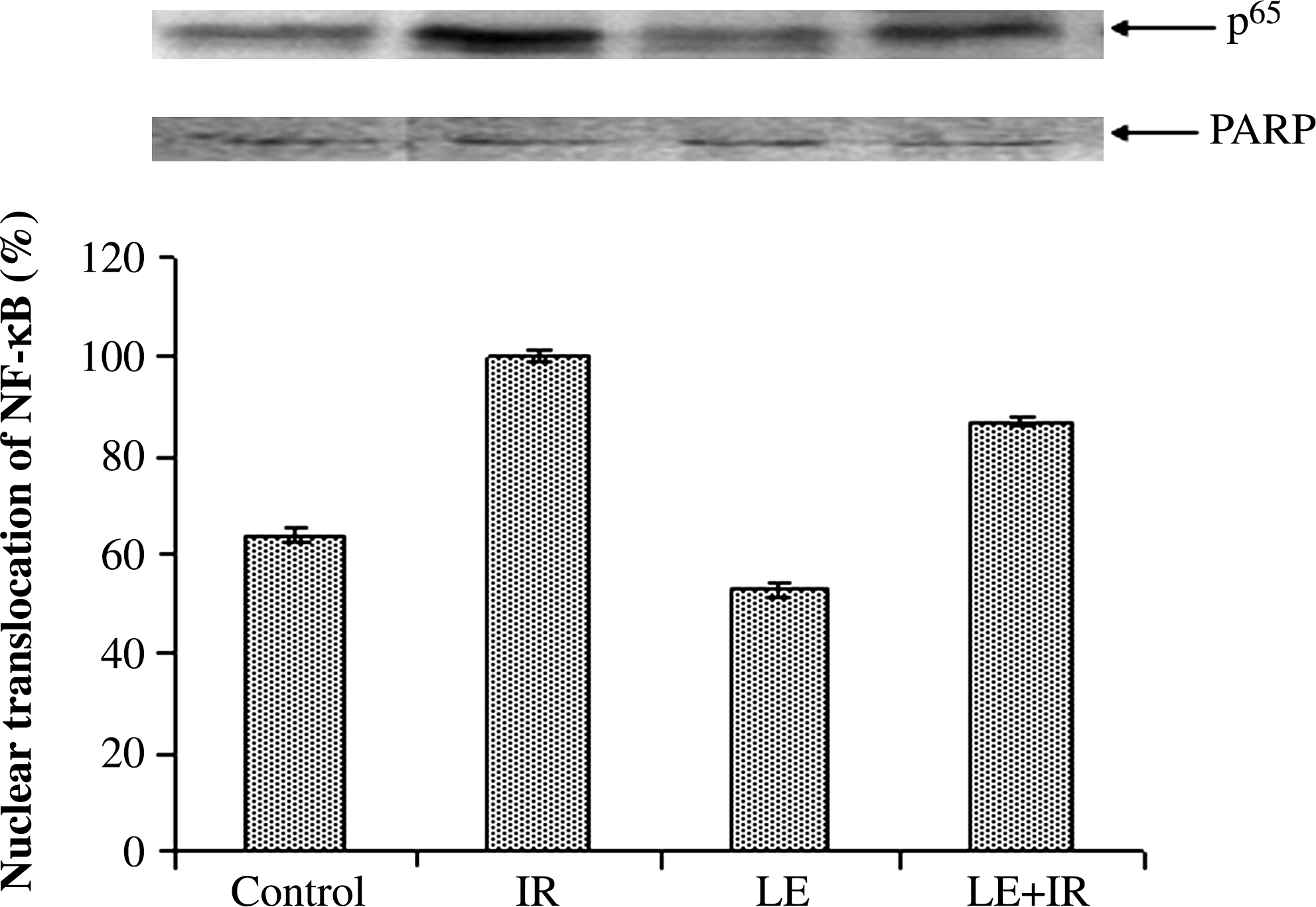
Nuclear factor κB (NF-κB) (p65) activation was determined by western blot analysis: lane 1, control group of mice; lane 2, mice irradiated with 5 Gy (IR); lane 3, mice treated with M. oleifera leaf extract (300 mg/kg of body weight) (LE); and lane 4, mice treated with M. oleifera leaf extract plus irradiated (5 Gy) (LE+IR). The p65 level in the irradiated group was set at 100%, and the other values are relative to that. Error bars are SEM of the data from three independent experiments. PARP, poly(ADP-ribose) polymerase.
MoLE administration ameliorated radiation-induced LPO
γ-Irradiation (5 Gy) induced a significant (P<.05) increase in the level of TBARS (5.68±0.43 nmol/g of tissue) compared with the control group (3.28±0.19 nmol/g of tissue). However, administration of MoLE for 15 consecutive days resulted in significant decreases (2.56±0.21 nmol/g of tissue) in TBARS compared with the control group. In contrast, pretreatment with MoLE ameliorated the effect of radiation exposure as TBARS levels were significantly (P<.05) decreased (2.88±0.21 nmol/g of tissue) compared with the irradiated group (Fig. 2).
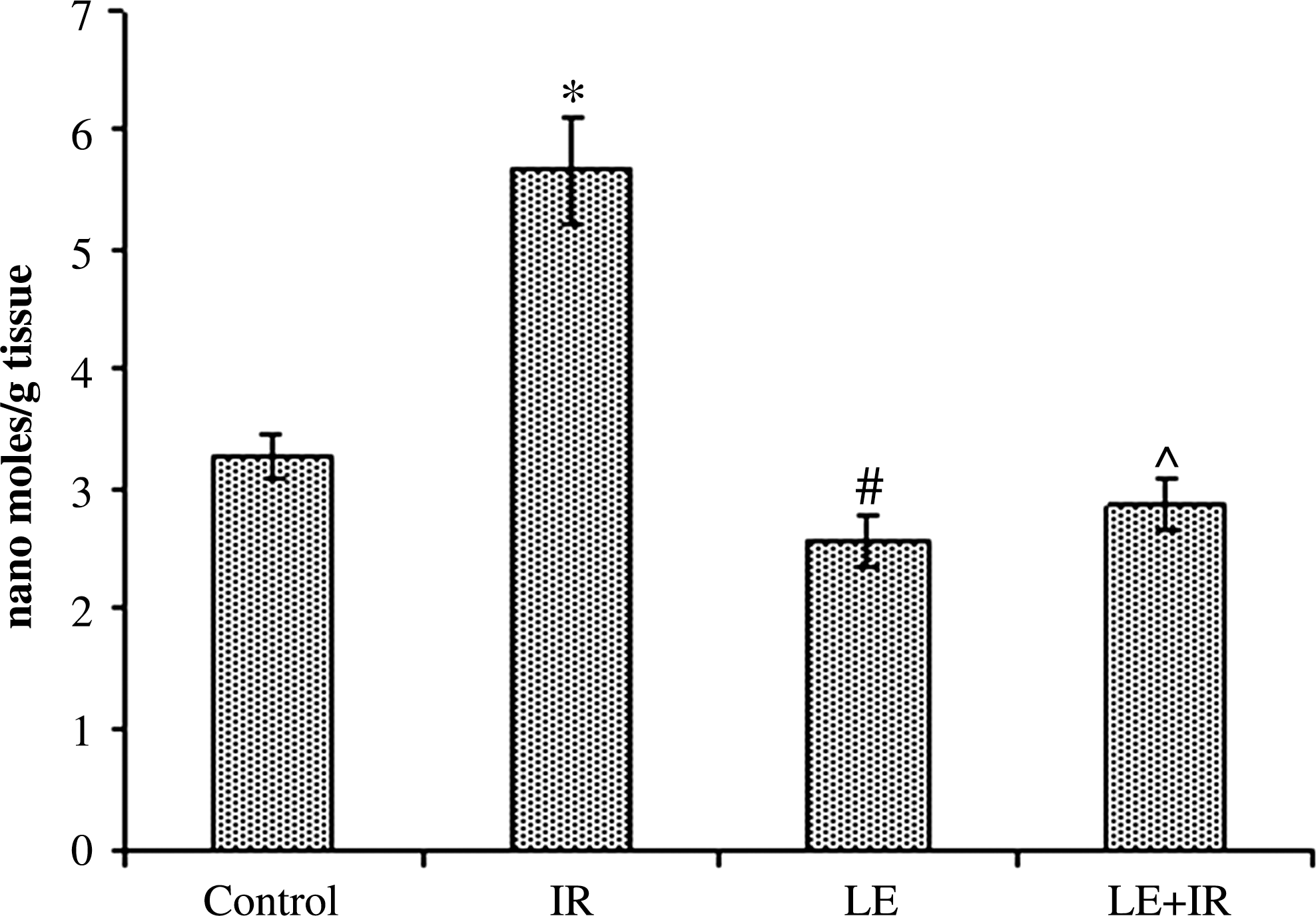
Effect of MoLE on radiation (5 Gy)-induced lipid peroxidation in mouse liver homogenates. Error bars are SEM for n=8. P<.05 was considered significant. Statistical comparison: *IR versus control; #LE versus control; ^LE+IR versus IR.
Determination of SOD activity
Control group SOD activity was 2.52±0.13 U/mg of protein. Mice exposed to 5 Gy of radiation (2.34±0.14 U/mg of protein) exhibited lower levels of SOD activity than controls. MoLE treatment augmented the SOD activity (2.87±0.15 U/mg of protein) compared with control animals. In the LE+IR group, the SOD activity (2.61±0.14 U/mg of protein) increased compared with the radiation control. However, the differences were not significant (Fig. 3).
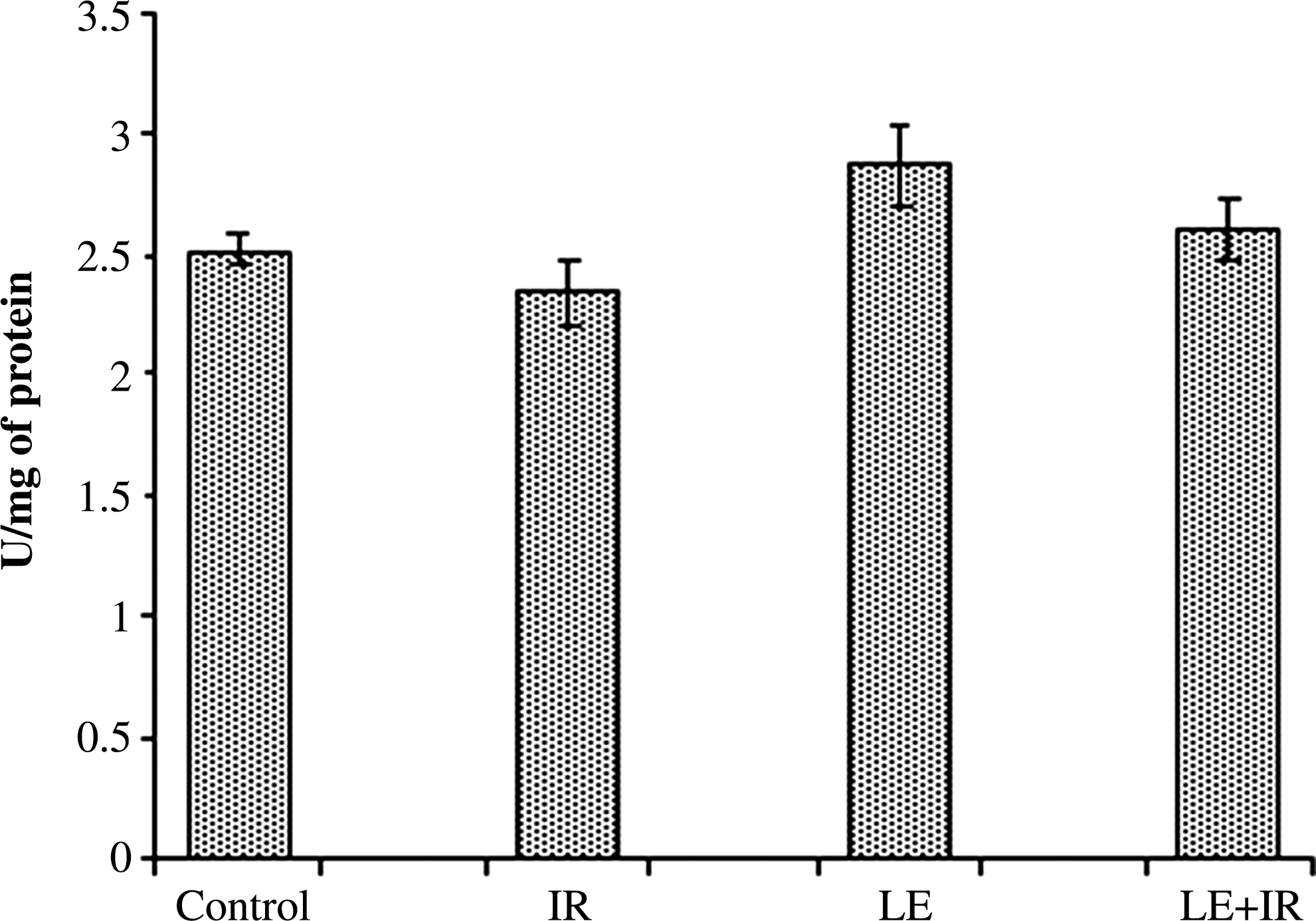
Effect of MoLE on radiation (5 Gy)-induced alteration of superoxide dismutase activity in mouse liver homogenates. Error bars are SEM for n=8.
Estimation of CAT activity
CAT activity of the control group was 30.35±2.04 μmol of H2O2 reduced/mg of protein. Mice exposed to 5 Gy of irradiation had lower levels of CAT activity (12.26±0.97 μmol of H2O2 reduced/mg of protein) in comparison with the controls. In the MoLE-treated group CAT activity was 32.82±1.78 μmol of H2O2 reduced/mg of protein. In the LE+IR group (29.01±1.38 μmol of H2O2 reduced/mg of protein), CAT activity was significantly higher than in irradiated animals (Fig. 4).
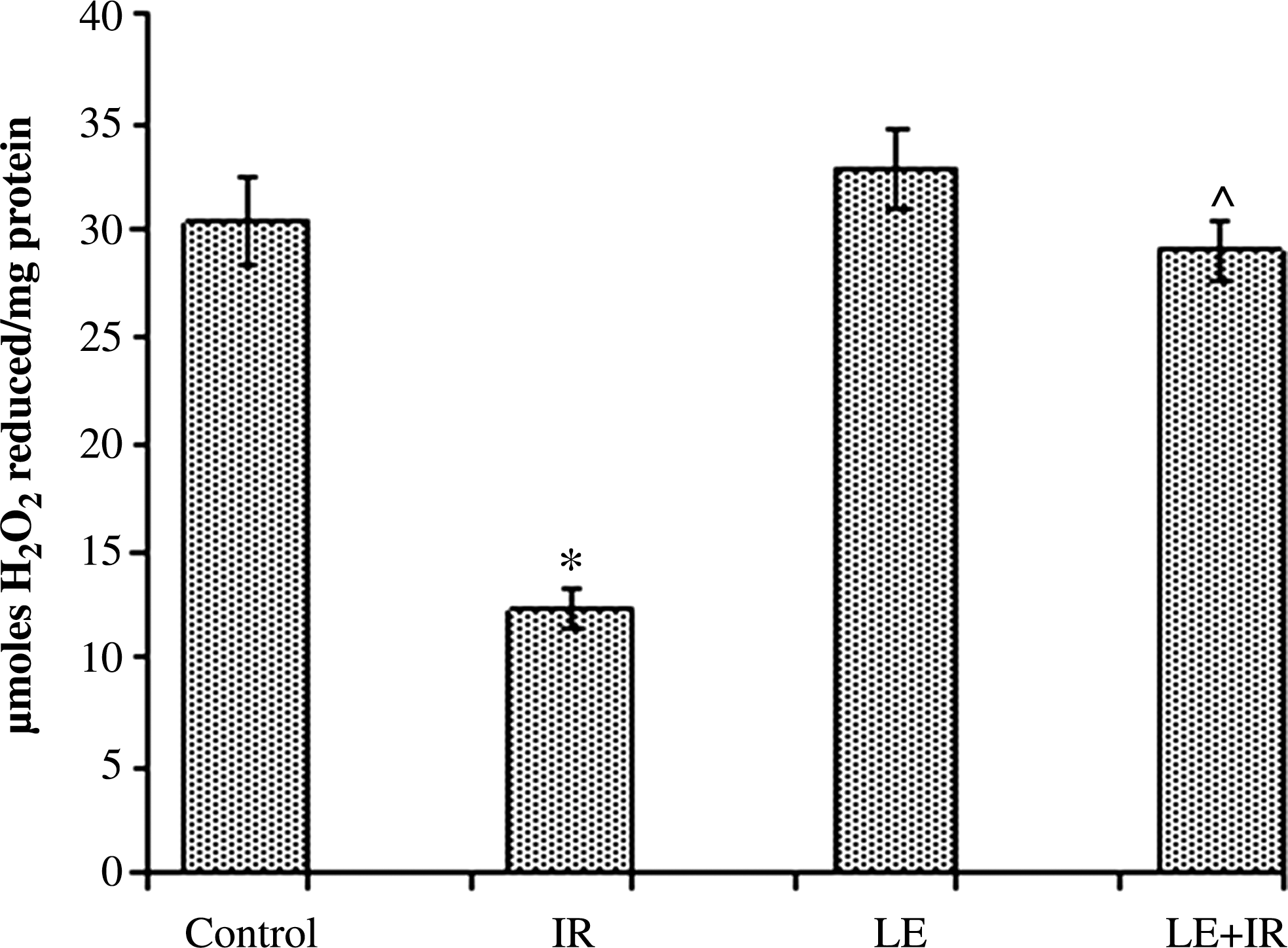
Effect of MoLE on radiation (5 Gy)-induced alteration of catalase activity in mouse liver homogenates. Error bars are SEM for n=8. P<.05 was considered significant. Statistical comparison: *IR versus control; #LE versus control; ^LE+IR versus IR.
Effect of MoLE on radiation-induced alteration in GSH level
γ-Irradiation induced a significant (P<.05) decrease (0.29±0.007 μmol/mg of protein) in GSH levels compared with the control group (0.43±0.006 μmol/mg of protein). Treatment with MoLE resulted in a nonsignificant increase (0.52±0.008 μmol/mg of protein) in GSH level compared with the control group. Pre-administration of MoLE to radiation-exposed mice significantly increased (0.41±0.01 μmol/mg of protein) GSH content compared with the irradiated group (P<.05) (Fig. 5).
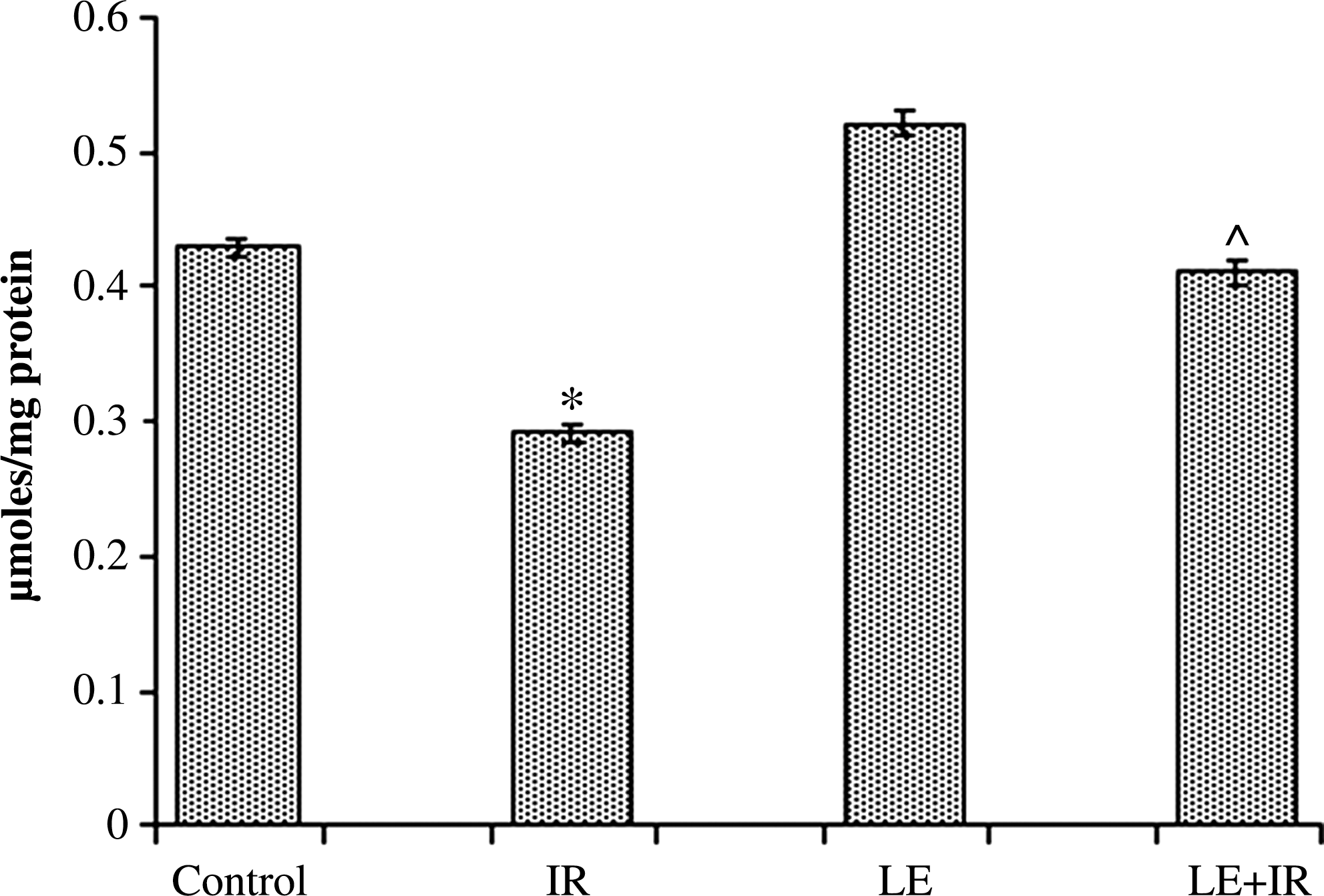
Effect of MoLE on radiation (5 Gy)-induced alteration of reduced glutathione in mouse liver homogenates. Error bars are SEM for n=8. P <.05 was considered significant. Statistical comparison: *IR versus control; #LE versus control; ^LE+IR versus IR.
Effect of MoLE on radiation-induced alteration in FRAP of liver homogenate
FRAP of liver homogenate denotes total antioxidant pool. We observed (Fig. 6) that γ-irradiation (5 Gy) induced a significant (P<.05) decrease (1.07±0.04 FRAP units) in FRAP level compared with the control group (2.04±0.06 FRAP units). Administration of MoLE significantly increased the FRAP level (2.62±0.1 FRAP units) compared with the control group. It is interesting that pre-administration of MoLE prior to radiation resulted in a significant (P<.05) increase in FRAP levels compared with the irradiated group (Fig. 6).
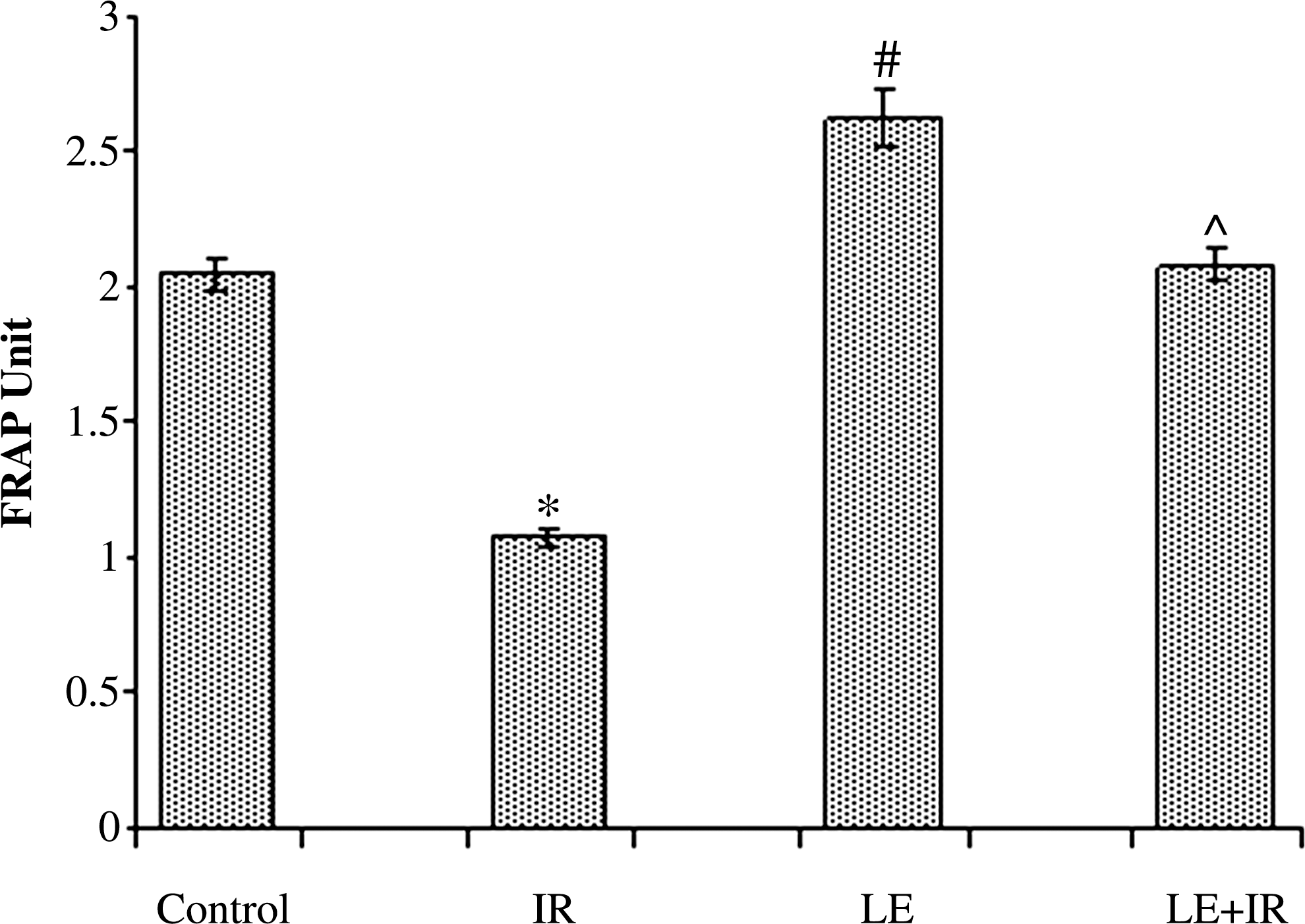
Effect of MoLE on radiation (5 Gy)-induced alteration of ferric reducing antioxidant power activity in mouse liver homogenates. Error bars are SEM for n=8. P<.05 was considered significant. Statistical comparison: *IR versus control; #LE versus control; ^LE+IR versus IR.
Discussion
In the present study, the nuclear translocation of NF-κB (p65) was augmented by γ-irradiation in the mouse model. γ-Radiation is associated with the production of reactive oxygen species, thereby activating the redox-sensitive transcription factor NF-κB. 15 NF-κB is a major mediator of all inflammatory responses. 16 It mediates inflammatory responses through the regulation of inflammatory cytokines (e.g., tumor necrosis factor-α, interleukin-1, and interleukin-6), adhesion molecules, inflammatory enzymes (e.g., cyclooxygenase-2, inducible nitric oxide synthase 2), etc. Our results also showed that 5 Gy of γ-radiation caused an increase in LPO level, whereas the activities of antioxidant enzymes (SOD and CAT), GSH levels, and FRAP values were decreased. Radiation-induced reactive oxygen species react with the molecules of cell membranes and cause LPO. LPO has been reported to be directly proportional to oxidative stress, in which the efficacy of various defense mechanisms is weakened. 17 Defense mechanisms may be strengthened by the addition of exogenous substances. It has been observed that LPO causes degradation of fine structures and alteration of integrity, fluidity, and permeability and functional loss of biomembranes, modifies low-density lipoprotein to pro-atherogenic and pro-inflammatory forms, and generates potential toxic products. 18 LPO products have been shown to be mutagenic and carcinogenic. 19 The reactive carbonyl compounds, the secondary products of LPO, modify biologically essential molecules such as proteins and DNA bases. 20 –22 Reactive oxygen species also negatively impact the antioxidant defense mechanism and decrease the intracellular concentration of GSH as well as the activities of enzymatic antioxidants. This could be due to an enhanced utilization of the antioxidant system during detoxification of the free radicals generated by irradiation.
In our study MoLE treatment prior to irradiation was found to protect the liver from radiation-induced inflammation as it decreased nuclear translocation of NF-κB and also reduced hepatic LPO. MoLE protected the liver from radiation-induced insults through scavenging the reactive oxygen species at the cost of its antioxidative principle. MoLE exhibited antioxidant activity directly through a free radical scavenging mechanism, indirectly through up-regulation of inherent antioxidant systems in the animal, or by both mechanisms.
Our results showed that MoLE neutralized the stable free radicals DPPH• and ABTS• + in an in vitro system. This radical scavenging activity was attributed to the presence of phytoantioxidants in the extract. Previously, it was reported that Moringa leaves act as a good source of natural antioxidants due to the presence of various types of antioxidant compounds such as ascorbic acid, flavonoids, phenolics, and carotenoids. 1 Moreover, we have identified ascorbic acid and various types of polyphenols such as catechin, epicatechin, ferulic acid, ellagic acid, and myricetin in MoLE (data not shown), which might neutralize the effects of free radicals generated by irradiation in mice.
In this study, it was also observed that SOD, CAT, and GSH levels were maintained after irradiation when the mice were pretreated with the leaf extract. This may be due to the leaf extract–fed group consuming high amounts of phytoantioxidants, which combat the radiation-induced free radicals; therefore SOD, CAT, and GSH were less utilized after irradiation. These dietary antioxidants restore SOD, CAT, and GSH and additionally play an essential role in combating against these free radicals. Furthermore, MoLE increased the total antioxidant pool in mice liver as the FRAP level elevated after the supplementation of leaf extract, which helps to ameliorate oxidative stress (Fig. 7).
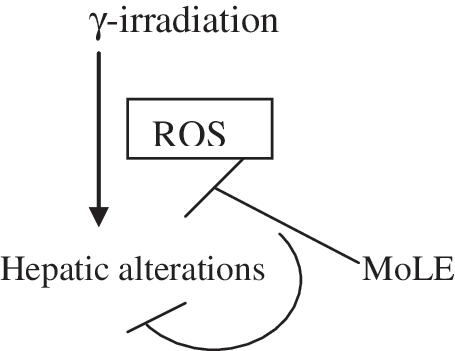
Proposed mechanism of effect of M. oleifera leaf extract on γ-radiation-mediated hepatic alterations.
We report here for the first time that M. oleifera leaf protects the liver from radiation hazards through its free radical scavenging principle. We also substantiate here that the radical scavenging principle of MoLE is responsible for preventing NF-κB entry into the nucleus and LPO. Characterization of this signaling pathway needs further study; however, our study is undoubtedly one of the first detailed studies of its kind linking radiation-induced cellular damage and its prevention.
Our present findings therefore also suggest the usefulness of M. oleifera leaf as a dietary component for providing general health benefits for protecting against accidental or deliberate radiation exposure. Therefore, we conclude that M. oleifera leaf, a popular vegetable, may prove to be a promising radioprotector for people of all socioeconomic backgrounds, especially for people in adverse and hazardous circumstances who are living in poverty as it is cost-effective as well as widely available. This common food-based medicinal plant can also be used as the supplement for patients who undergo radiotherapy; however, this needs further investigation before clinical trials.
Footnotes
Acknowledgments
A Junior Research Fellowship to M.S. from UGC-DAE, University of Calcutta, Kolkata, India, is gratefully acknowledged. The authors are also thankful to Dr. M. Nandi of SINP, Kolkata for providing necessary irradiation facilities. S.D. acknowledges the financial assistance of the UGC-DAE collaborative research scheme of Kolkata Centre.
Author Disclosure Statement
No competing financial interests exist.