
Abstract
Soybean is a useful component of traditional Korean medicine with well-documented health-promoting effects. We investigated the effects of alcohol-fermented soybean (AFS) on immune function. When AFS treatment was used in combination with recombinant interferon-γ (rIFN-γ), there was a marked cooperative induction of nitric oxide (NO) and tumor necrosis factor (TNF)-α production in mouse peritoneal macrophages. AFS increased the expression of inducible NO synthase mRNA and protein in rIFN-γ-primed macrophages. Treating macrophages with pyrrolidine dithiocarbamate, an inhibitor of nuclear factor-κB (NF-κB), decreased the synergistic effects of AFS. In addition, AFS in combination with rIFN-γ increased the phosphorylation of p38 and c-Jun N-terminal kinase (JNK) but not extracellular signal-regulated kinase. However, AFS had no effect on phosphorylation of mitogen-activated protein kinases by itself. The p38 inhibitor SB203580 or the JNK inhibitor SP600125 inhibited the AFS-induced NO and TNF-α production. When AFS was used in combination with rIFN-γ, there was a co-operative activation of NF-κB and receptor-interacting protein 2 (Rip2)/IκB kinase (IKK)-β. Our results indicate that AFS increases the production of NO and TNF-α through the activation of Rip2/IKK-β in rIFN-γ-primed macrophages.
Introduction
S
Nitric oxide (NO) is a highly reactive molecule produced from the guanidino nitrogen of arginine by NO synthase (NOS).
8
Over the past decade, NO has become recognized as a potent macrophage-derived effector molecule against various bacteria, parasites, and tumors.
9
The original evidence of tumor cell cytostasis and cytotoxicity was found in macrophage–tumor cell co-cultures in which cytokine- and/or lipopolysaccharide (LPS)-stimulated macrophages inhibited metabolic functioning of co-cultured tumor cells.
10
The activity of NOS can be inhibited by N
G-monomethyl-
The pro-inflammatory cytokine tumor necrosis factor (TNF)-α regulates systemic responses to microbial infection or tissue injury. 13 Production of TNF-α protein was enhanced by the presence of interferon-γ (IFN-γ). TNF-α then acted as an autocrine signal to amplify IFN-γ-induced production of NO in macrophages. 13
Pathways involving mitogen-activated protein kinases (MAPKs) are central components of the intracellular signaling networks that control many aspects of cellular physiology, including inflammation, cell proliferation, differentiation, and apoptosis. 14 Extracellular signal-regulated kinase (ERK), p38 MAPK, and the c-Jun N-terminal kinase (JNK) belong to the MAPK family. In response to appropriate stimuli, MAPK is activated by phosphorylation on both adjacent threonine and tyrosine residues. Whereas ERK has been classically associated with growth- and differentiation-inducing signals, p38 and JNK are involved in inflammatory cytokines and environmental stress inducers. 15
NF-κB plays a critical role in the expression of many genes involved in immune and inflammatory responses. 16,17 In cells that are not stimulated, Rel protein dimers, mainly p50 and p65 subunits, are sequestered in the cytoplasm complexed with one of the several inhibitors of NF-κB. The activation of NF-κB is the consequence of phosphorylation of two specific serines near the N terminus of IκB-α and its degradation. The phosphorylation of IκB-α leads to ubiquitination, which targets the protein for degradation by the 26S proteasome and the translocation of NF-κB to the nucleus. 18
The IκB kinase (IKK)-β enzyme complex is part of the upstream NF-κB signal transduction cascade, 19 and receptor-interacting protein 2 (Rip2), a serine/threonine kinase, is also essential for NF-κB activation through the TNF receptor and Toll-like receptors. 20,21
In the present study, we show that AFS synergistically induces NO and TNF-α production by peritoneal macrophages when the cells are treated with recombinant IFN-γ (rIFN-γ). To investigate the mechanism of AFS-induced NO and TNF-α production, we examined iNOS and COX-2 expression and activations of MAPKs, NF-κB, Rip2, and IKK-β.
Materials and Methods
Reagents
Soybeans were purchased from Omniherb (Youngchun, Korea). Murine rIFN-γ, recombinant TNF-α, biotinylated TNF-α, and anti-murine TNF-α were purchased from R&D Systems (Minneapolis, MN, USA). N-(1-Naphthyl)-ethylenediamine dihydrochloride, LPS, sodium nitrite, and pyrrolidine dithiocarbamate (PDTC) were purchased from Sigma (St. Louis, MO, USA). COX-2, iNOS, p38, phosphorylated p38 (pp38), ERK, phosphorylated ERK (pERK), JNK, phosphorylated JNK (pJNK), NF-κB (p65), Rip2, and IKK-β antibodies were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). NGMMA was purchased from Calbiochem (San Diego, CA, USA). Thioglycollate was purchased from Difco Laboratories (Detroit, MI, USA). Dulbecco's modified Eagle's medium containing
Animals
The original stock of male C57BL/6J mice was purchased from Daehan Biolink Co. (Daejeon, Korea). Animals were housed at a temperature of 23±1°C with a 12/12-hour light–dark cycle. Food and water were available ad libitum. All protocols were approved by the Institutional Animal Care and Use Committee of Kyung Hee University (Seoul, Korea).
Preparation of AFS
Soybeans (500 g) were boiled with 5 L of water and concentrated to about 3 L. We prepared 2 kg of steamed rice, 1 kg of malt, and 50 g of yeast. These were mixed with soybean water extract. The mixture was kept at 20–25°C for 7 days. This fermented material was filtered and heated to 55–60°C for 5 minutes, and voucher specimens (number KH090302) were deposited at the Herbarium at the College of Oriental Medicine, Kyung Hee University.
Assay for endotoxin determination
The AFS was found to be free of endotoxin as determined within the limits of the E-TOXATE assay kit (Sigma), performed according to the manufacturer's protocol. In this assay, saturation occurred at 40 EU/mL, and the resolution limit was >0.1 EU/mL.
Peritoneal macrophage cultures
Thioglycollate-elicited macrophages were harvested from mice by peritoneal lavage 3–4 days after intraperitoneal injection of 2.5 mL of thioglycollate as reported previously. 22 Then, the cells were distributed into Dulbecco's modified Eagle's medium, which was supplemented with 10% heat-inactivated fetal bovine serum, in culture plates, incubated for 3 hours at 37°C in an atmosphere of 5% CO2, washed three times with Hanks' balanced salt solution to remove nonadherent cells, and equilibrated with Dulbecco's modified Eagle's medium that contained 10% fetal bovine serum before treatment.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
Peritoneal macrophages (2.5×105 cells per well) were cultured with rIFN-γ (10 U/mL) for 6 hours. The cells were then stimulated with various concentrations of AFS (0.01–1 mg/mL). Cell aliquots were incubated with 20 μL of a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide solution (5 mg/mL) for 4 hours at 37°C under 5% CO2 and 95% air. Then, 250 μL of dimethyl sulfoxide was added to extract the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide–formazan, and the absorbance of each well read at 540 nm with an automatic microplate reader (Molecular Devices Corp., Sunnyvale, CA, USA).
Measurement of nitrite concentration
Peritoneal macrophages (2.5×105 cells per well) were cultured with rIFN-γ (10 U/mL) for 6 hours. The cells were then stimulated with various concentrations of AFS. After 48 hours of culture, NO synthesis in cell cultures was measured by a microplate assay method, as previously described, 22 using sodium nitrite as the standard. The cell-free medium alone contained 5–8 μM NO2 −. This value was determined in each experiment and subtracted from the value obtained from the medium with peritoneal macrophages.
Assay of TNF-α release
Peritoneal macrophages (2.5×105 cells per well) were incubated with rIFN-γ (10 U/mL), AFS, rIFN-γ plus LPS (10 μg/mL), and rIFN-γ plus various concentrations of AFS (0.01–1 mg/mL) for 24 hours. Then the amount of TNF-α secreted by the cells was measured by a modified enzyme-linked immunosorbent assay, as described previously. 23
Nuclear protein extraction
Nuclear extracts were prepared as described previously. 24 In brief, after cell activation for the times indicated cells were washed with ice-cold phosphate-buffered saline and resuspended in 60 μL of buffer A (10 mM HEPES/KOH, 2 mM MgCl2, 0.1 mM EDTA, 10 mM KCl, 1 mM dithiothreitol, and 0.5 mM phenylmethylsulfonyl fluoride, pH 7.9). The cells were allowed to swell on ice for 15 minutes, lysed gently with 2.5 μL of 10% Nonidet P-40, and centrifuged at 2,000 g for 10 minutes at 4°C. The nuclei pellet was resuspended in 40 μL of buffer B (50 mM HEPES/KOH, 50 mM KCl, 300 mM NaCl, 0.1 mM EDTA, 10% glycerol, 1 mM dithiothreitol, and 0.5 mM phenylmethylsulfonyl fluoride, pH 7.9), left on ice for 20 minutes, and inverted. The nuclear debris was then centrifuged at 15,000 g for 15 minutes. The supernatant (nuclear extract) was collected, frozen in liquid nitrogen, and stored at −70°C until the analysis was conducted.
Western blot analysis
Peritoneal macrophages (5×106 cells per well) were incubated for 6 hours with rIFN-γ (R&D Systems) (10 U/mL). The cells were then stimulated with AFS (1 mg/mL) or LPS (10 μg/mL). Whole-cell lysates were made by boiling peritoneal macrophages in sample buffer (62.5 mM Tris-HCl [pH 6.8], 2% sodium dodecyl sulfate, 20% glycerol, and 10% 2-mercaptoethanol). Proteins in the cell lysates were then separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to nitrocellulose paper. The membrane was then blocked with 5% skim milk in phosphate-buffered saline containing Tween-20 and then incubated with antibodies (Santa Cruz Biotechnology). After washing in phosphate-buffered saline containing Tween-20 three times, the blot was incubated with 1:3,000 diluted anti-rabbit or anti-mouse immunoglobulin G, horseradish peroxidase–linked whole antibody (Amersham Corp., Newark, NJ, USA), and the antibody-specific proteins were visualized using the enhanced chemiluminescence detection system according to the recommended procedure (Amersham Corp.). The relative protein amounts were estimated by an FC-26WL image analyzer (Vilber Lourmat, Marne La Vallée, France).
Reverse transcription–polymerase chain reaction analysis
Total RNA was isolated from macrophages according to the manufacturer's specification using an easy-BLUE RNA extraction kit (iNtRON Biotech, Seoul). Total RNA (2.0 μg) was heated at 75°C for 5 minutes and then chilled on ice. Each sample was reverse-transcribed to cDNA for 60 minutes at 42°C using a cDNA synthesis kit (iNtRON Biotech). Polymerase chain reaction was performed with the following primers: for TNF-α, 5′-ATG AGA ACA GAA AGCA TGA TC-3′ and 5′-TAC AGG CTT GTC ACT CGA ATT-3′; for iNOS, 5′-TCA CTG GGA CAG CAC AGA AT-3′ and 5′-TGT GTC TGC AGA TGT GCT GA-3′; for COX-2, 5′-GGA GAG ACT ATC AAG ATA GTG ATC-3′ and 5′-ATG GTC AGT AGA CTT TTA CAG CTA-3′; and for glyceraldehyde 3-phosphate dehydrogenase (GAPDH), 5′-GGC ATG GAC TGT GGT CAT GA-3′; 5′-TTC ACC ACC ATG GAG AAG GC-3′. GAPDH was used to verify if equal amounts of RNA were used for reverse transcription and polymerase chain reaction amplification from different experimental conditions. The annealing temperature was 58°C for iNOS, 60°C for TNF-α, and 62°C for GAPDH, respectively. Products were electrophoresed on a 1.5% agarose gel and visualized by staining with ethidium bromide. GAPDH mRNA was used as a control of mRNA loading. The relative mRNA amounts were estimated by an FC-26WL image analyzer.
Statistical analysis
Results were expressed as mean±SEM values of independent experiments, and statistical analyses were performed by one-way analysis of variance with Tukey's and Duncan's post hoc tests to separate the differences between groups. All statistical analyses were performed using SPSS version 11.0 statistical analysis software (SPSS Inc., Chicago, IL, USA). A value of P<.05 was considered significant.
Results
Effects of AFS on NO production in peritoneal macrophages
As shown in Figure 1, AFS had no effect on NO production in resting mouse peritoneal macrophages. IFN-γ (10 U/mL) alone does not cause the induction of NO production, in accordance with previous reports. 25,26 However, when IFN-γ was used in conjunction with LPS, there was a marked augmentation of the production of NO. When mouse peritoneal macrophages were primed for 6 hours with murine rIFN-γ and then treated with AFS, NO production was increased compared with nonprimed conditions. AFS did not affect the viability of cells at those concentrations (data not shown).

Effects of alcohol-fermented soybean (AFS) on nitric oxide synthesis in peritoneal macrophages. Cells incubated with recombinant interferon-γ (IFN-γ) were then stimulated with various concentrations of AFS for 6 hours after incubation. After 48 hours of culture, nitric oxide release was measured. Levels of nitric oxide (nitrite) released into the medium are mean±SEM values of three independent experiments performed in duplicate. *P<.05 compared with recombinant IFN-γ alone. LPS, lipopolysaccharide.
Effects of AFS on rIFN-γ-primed iNOS expression
Data in Figure 2 show the effects of rIFN-γ plus AFS treatments on the expression of iNOS mRNA and protein in mouse peritoneal macrophages. AFS (1 mg/mL) synergistically increased the expression of iNOS mRNA and protein in rIFN-γ-primed mouse peritoneal macrophages.
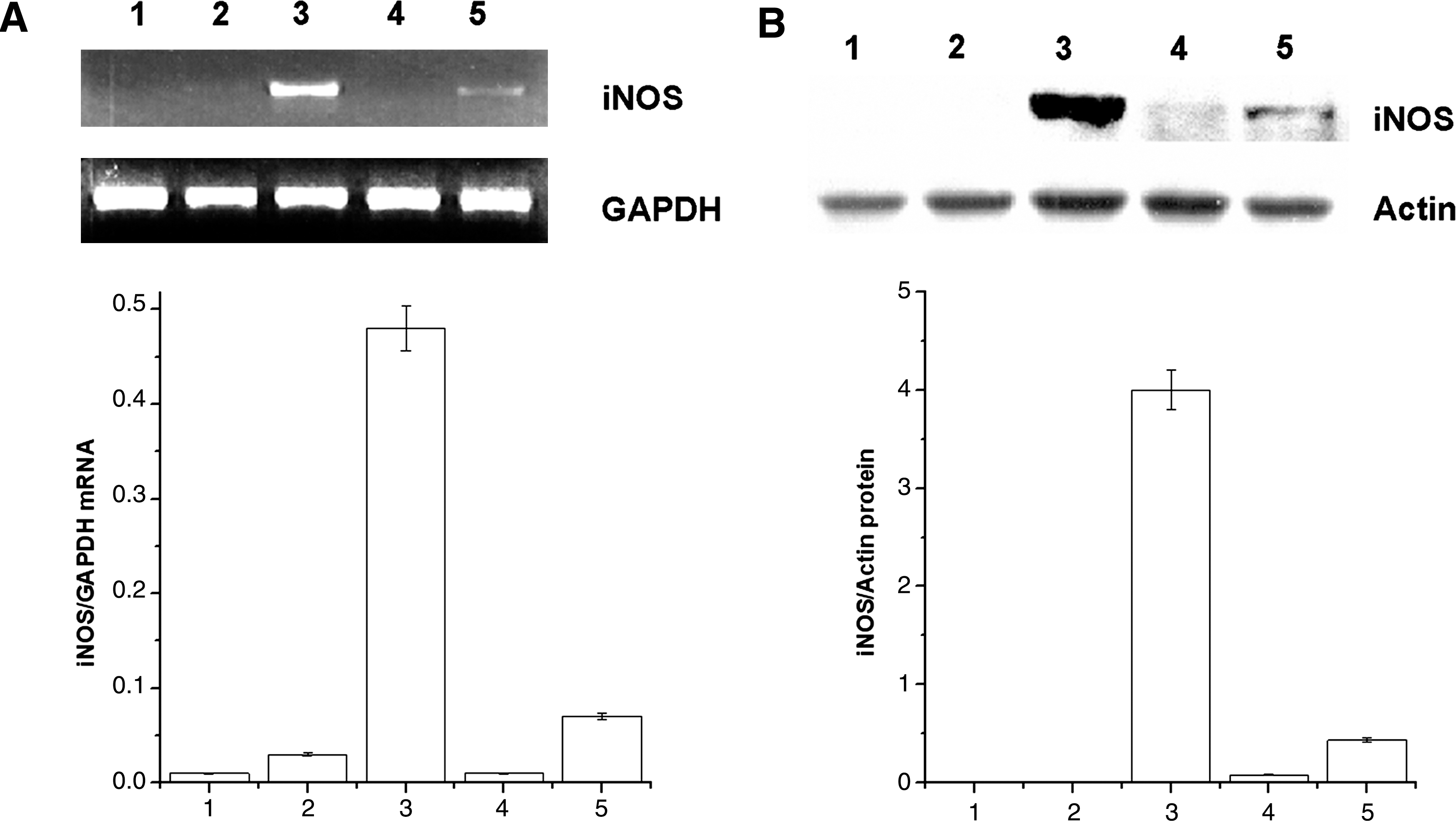
Effects of AFS on the expressions of inducible nitric oxide synthase (iNOS) mRNA and protein in peritoneal macrophages: lane 1, blank; lane 2, recombinant IFN-γ; lane 3, recombinant IFN-γ+LPS; lane 4, AFS (1 mg/mL); and lane 5, recombinant IFN-γ+AFS (1 mg/mL). (
Inhibition of AFS-induced NO production by NGMMA
To determine if the signaling mechanism in AFS-induced NO production participates in the
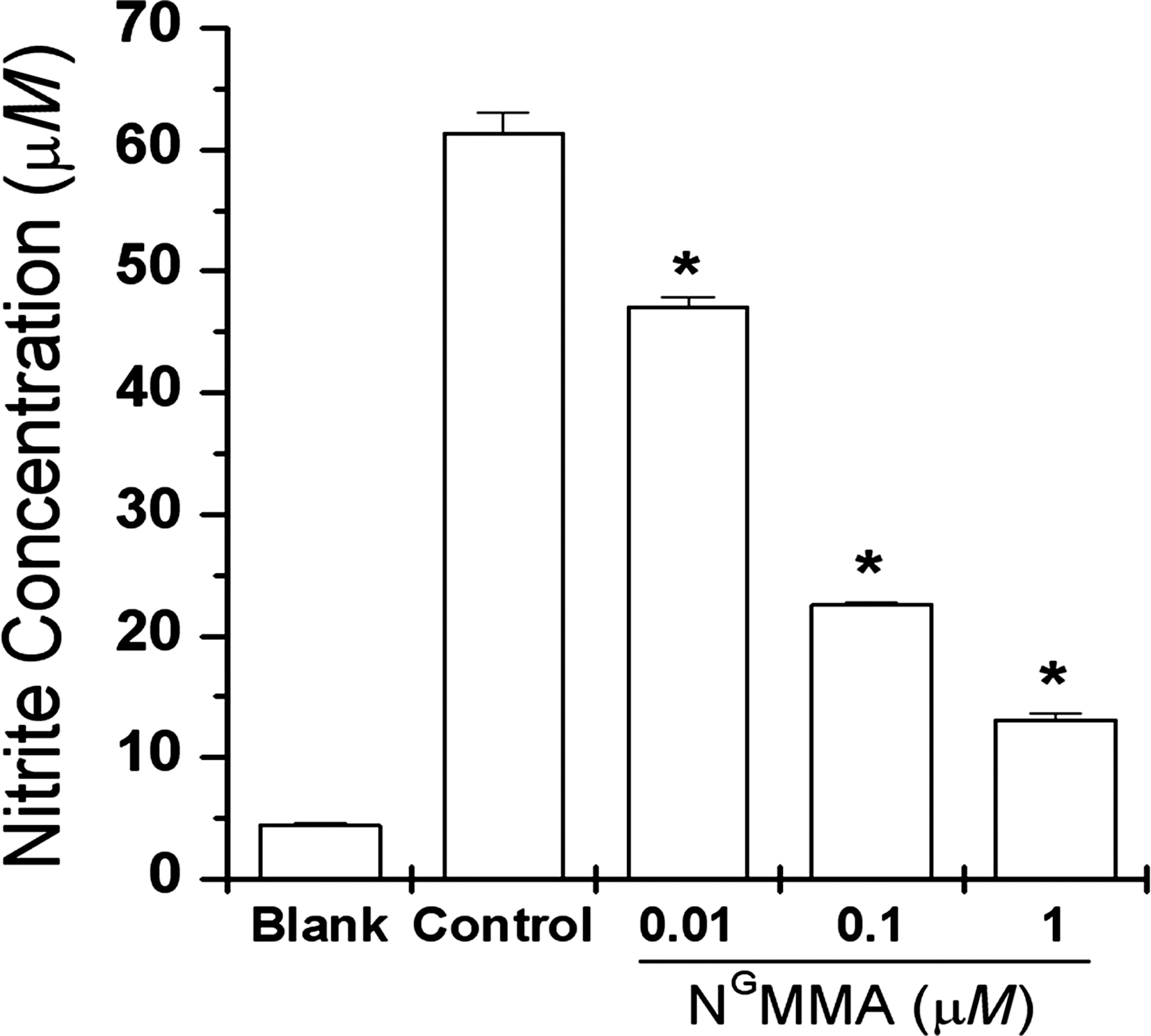
Effects of N
G-monomethyl-
Effects of AFS on rIFN-γ-primed COX-2 expression
Reverse transcription–polymerase chain reaction and western blot analysis for COX-2 revealed that COX-2 mRNA and protein expressions were not changed by treatment with AFS (1 mg/mL) and rIFN-γ (Fig. 4).
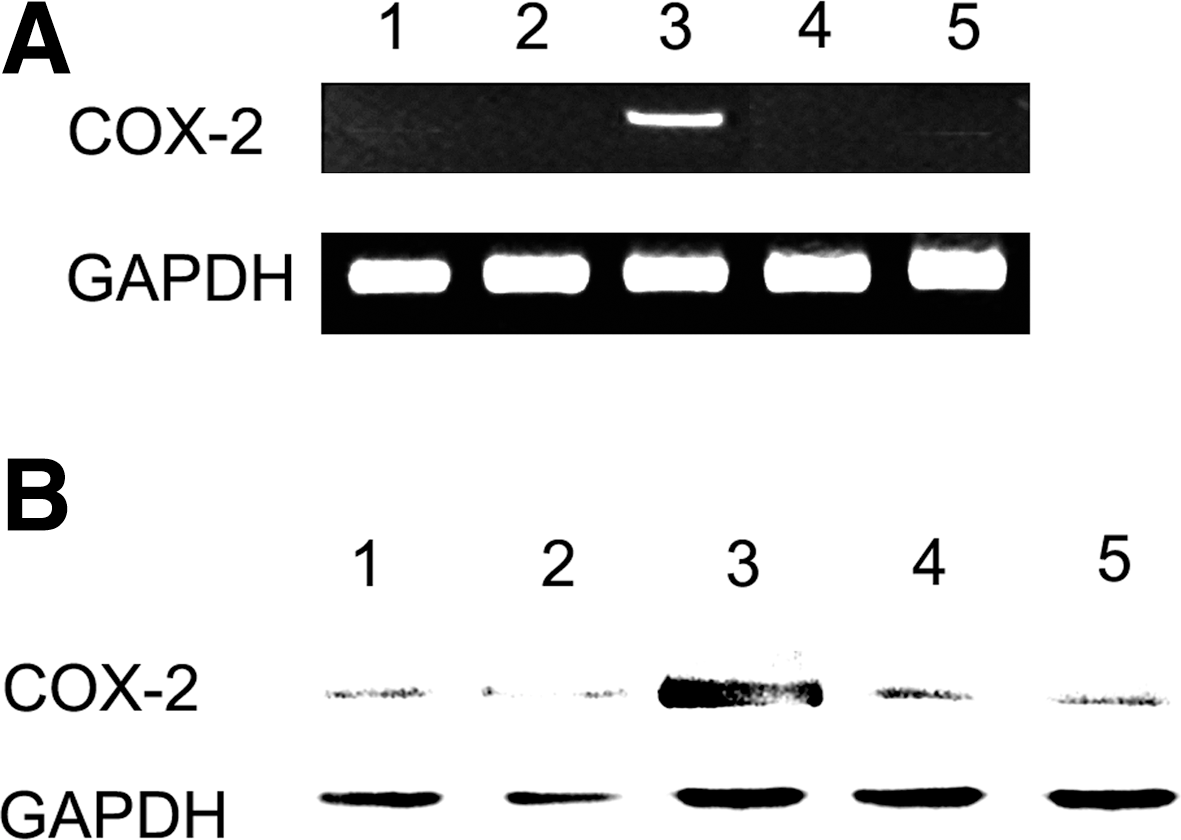
Effects of AFS on expression of cyclooxygenase-2 (COX-2) mRNA and protein in peritoneal macrophages: lane 1, blank; lane 2, recombinant IFN-γ; lane 3, recombinant IFN-γ+LPS; lane 4, AFS (1 mg/mL); and lane 5, recombinant IFN-γ+AFS (1 mg/mL). (
Effects of AFS on rIFN-γ-primed TNF-α expression
We next examined the synergistic effects of AFS on rIFN-γ-induced TNF-α production and expression. Mouse peritoneal macrophages secreted very low levels of TNF-α after a 24-hour incubation with medium alone or rIFN-γ alone. However, AFS in combination with rIFN-γ markedly increased TNF-α production and expression (Fig. 5). It is known that PDTC, an antioxidant compound, inhibits NF-κB activation. 27 The signaling mechanism of AFS on TNF-α was evaluated by examining the influence of PDTC in rIFN-γ plus AFS-treated mouse peritoneal macrophages. PDTC (100 μM) treatment of the rIFN-γ plus AFS-treated mouse peritoneal macrophages decreased the synergistic effects of AFS on TNF-α mRNA expression (Fig. 5B).
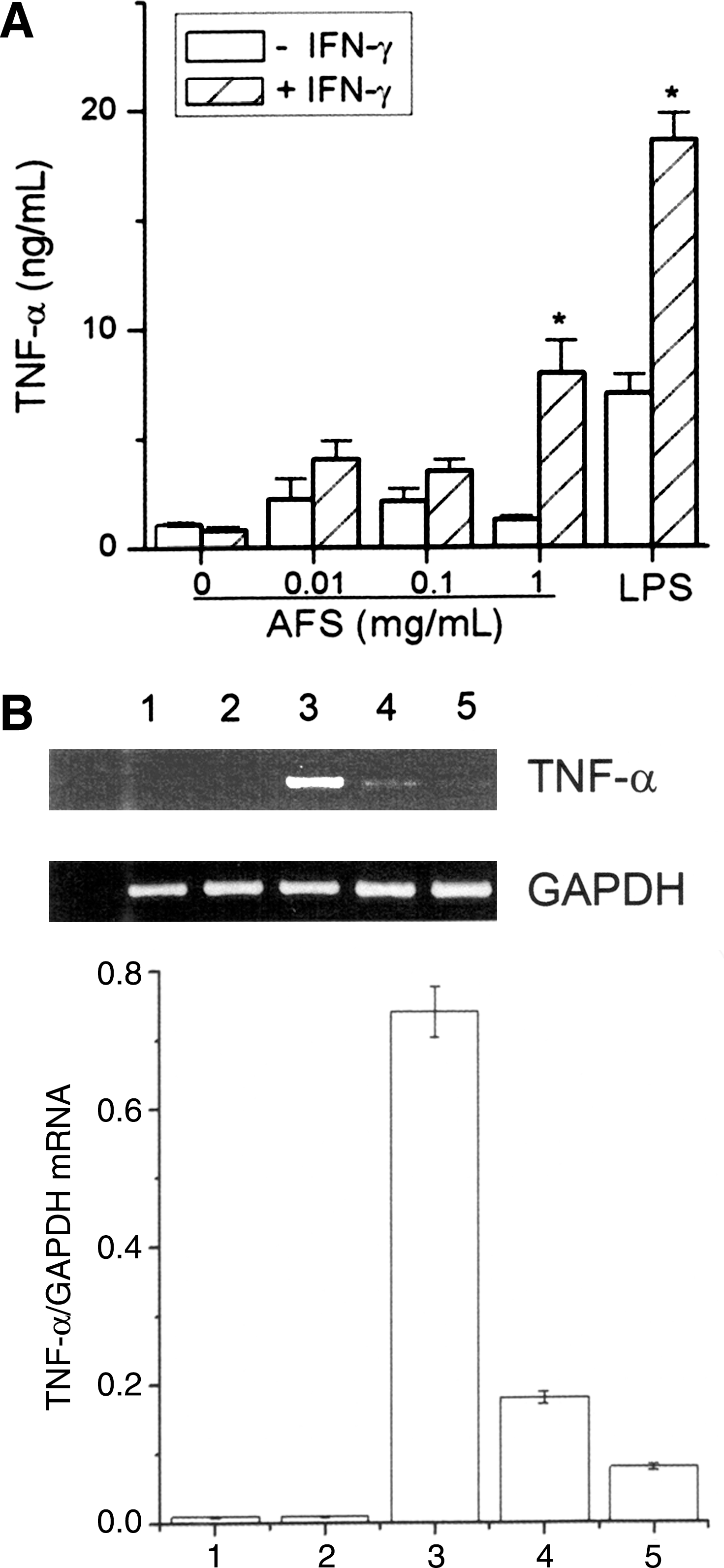
Effects of AFS on expression of tumor necrosis factor-α (TNF-α) in peritoneal macrophages. The cells (2.5×105 cells per well) were stimulated with various concentrations of AFS or recombinant IFN-γ (10 U/mL) plus AFS. (
Effects of AFS on MAPK activation
To demonstrate that AFS activates the MAPK pathway, we studied the effect of AFS on the phosphorylation of p38, ERK, and JNK in rIFN-γ-primed peritoneal macrophages. AFS in combination with rIFN-γ increased the phosphorylation of p38 and JNK but not ERK (Fig. 6A). However, AFS had no effect on phosphorylation of MAPKs by itself (Fig. 6A). We also investigated the influence of MAPK inhibitors (the p38 inhibitor SB203580 or the JNK inhibitor SP600125) on NO and TNF-α production. As shown in Figure 6B and C, SB203580 or SP600125 inhibited the AFS-induced NO and TNF-α production.
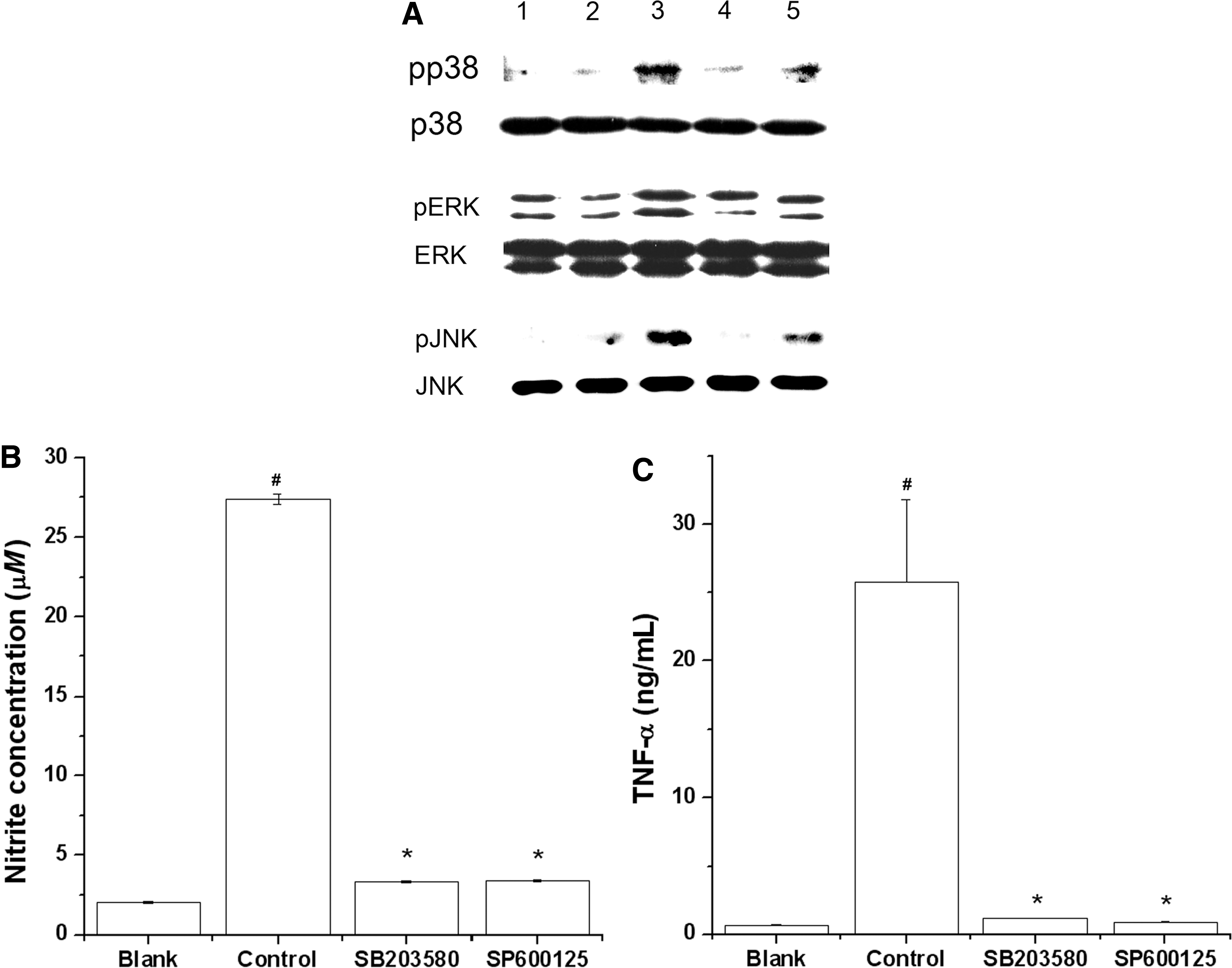
Effects of AFS on mitogen-activated protein kinase activation in peritoneal macrophages. (
Effect of AFS on NO production through NF-κB, Rip2, and IKK-β activation
We also investigated the effects of AFS on the signal transduction pathway of NO production. NF-κB, Rip2, and IKK-β activations were assessed by western blot analysis. As shown in Figure 7, stimulation of peritoneal macrophages with LPS after the treatment of rIFN-γ resulted in increased activation of NF-κB, Rip2, and IKK-β. When AFS was used in combination with rIFN-γ, there was a marked co-operative activation of NF-κB, Rip2, and IKK-β. However, AFS had no effect on activation of NF-κB, Rip2, and IKK-β by itself (data not shown).
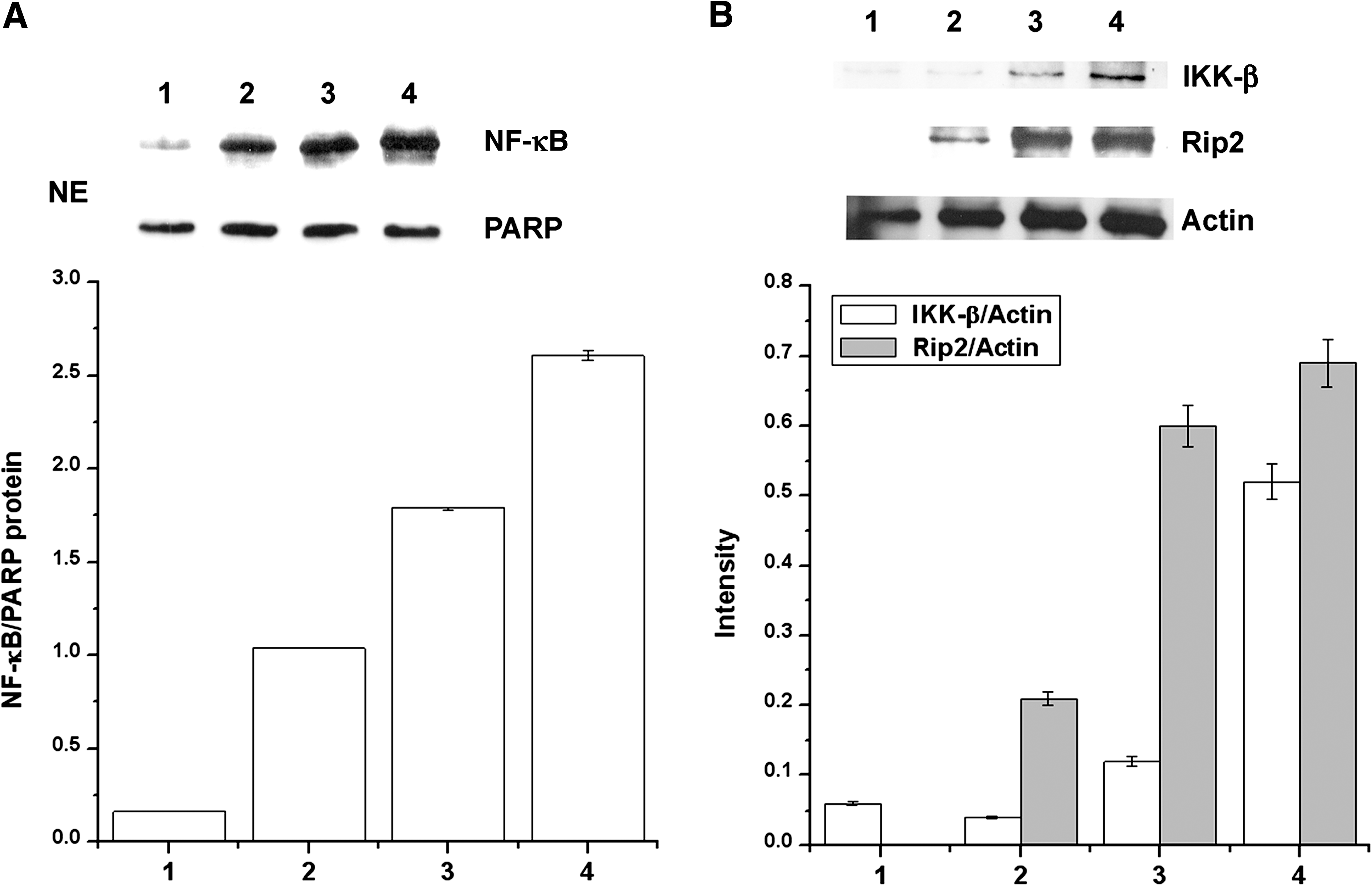
Effect of AFS on activation of nuclear factor-κB (NF-κB), receptor-interacting protein 2 (Rip2), and IκB kinase (IKK-β) in peritoneal macrophages: lane 1, blank; lane 2, recombinant IFN-γ; lane 3, recombinant IFN-γ+LPS; and lane 4, recombinant IFN-γ+AFS (1 mg/mL). (
Discussion
In this study, we demonstrated that AFS induced NO production in mouse peritoneal macrophages could be highly stimulated when combined with rIFN-γ. AFS had a maximal effect on NO production at a concentration of 1 mg/mL in rIFN-γ-treated mouse peritoneal macrophages. The results of this study suggest that AFS may provide a second signal for synergistic induction of NO production in mouse peritoneal macrophages. NGMMA, an analog of
At present, the precise physiological significance of AFS-induced NO production is unknown. However, over the past decade, NO as a potent macrophage-derived effector molecule against various bacteria, parasites, and tumors has received increasing attention. 28 NO has been reported to play an important role in the mitogenic effect of angiogenic factors (e.g., in the angiogenic process). Eroğlu et al. 29 showed that a large tumor burden was associated with significantly increased levels of vascular endothelial growth factor and NO. In addition, Klotz et al. 30 reported that bladder carcinoma tissue had a high iNOS content, whereas benign tissue did not. NO generation by iNOS also influences the cytotoxicity of macrophages and tumor-induced immunosuppression. Increased NO production by AFS indicates that it may exert antimicrobial, antitumor, and antiviral activities under specific conditions in vivo. NO has emerged as an important molecule having functions such as diverse as vasodilation, neural communication, and host defense. 31 It is tempting to hypothesize that this molecule is involved in the local control of various fundamental processes.
It has been previously reported that LPS stimulation of rIFN-γ-primed macrophages induces NF-κB activation. 32 Expression of iNOS and TNF-α genes is dependent on the activation of NF-κB and the phosphorylation of p38, ERK, and JNK. 33,34 We found that addition of the NF-κB modulator PDTC inhibits the synergistic effect of AFS with rIFN-γ on TNF-α expression. Moreover, AFS in combination with rIFN-γ increased the phosphorylation of p38 and JNK. Treatment with SB203580 or SP600125 decreased the synergistic effects of AFS on NO and TNF-α production. Therefore, we suggest that AFS increases NO and TNF-α production via the activation of p38, JNK, and NF-κB in rIFN-γ-primed macrophages.
Chen et al. 12 reported that COX-2 induced NO production in macrophages. However, AFS did not affect COX-2 expression. From this, we can deduce that AFS induces NO production via a COX-2-independent pathway.
The NF-κB system may provide a future target for cancer therapy. Rip2 and IKK complexes may play an important role for NF-κB activation. 35 In this study, we also confirmed that AFS increased the Rip2 and IKK activation. The results of this study suggest that AFS activates NF-κB via interaction with Rip2 and IKK-β.
Soybeans are particularly rich in isoflavones, such as genistein and daidzein. In a previous study, isoflavones up-regulated LPS-induced NO production via the activation of iNOS; moreover, isoflavones enhanced LPS-induced TNF-α production. 36 This study showed that AFS enhanced the immunomodulatory response induced by LPS plus rIFN-γ in mouse peritoneal macrophages. However, the effects of the individual ingredients in AFS on the present experimental model were not investigated. Thus, further investigation is necessary to explain the individual effects of active compounds, such as isoflavones in AFS, on LPS plus rIFN-γ-induced peritoneal macrophages.
Macrophages may have a receptor for yeast. In this study, AFS was fermented at 20–25°C for 7 days. Then, the fermented materials were filtered. LPS contamination was not detected by the endotoxin test. Moreover, we also confirmed that AFS alone did not affect the NO production and TNF-α secretion. From this, we can predict that AFS does not contain any products of heat-killed yeast.
In conclusion, our results suggest that AFS acts as an accelerator of peritoneal macrophage activation by rIFN-γ, via a process involving
Footnotes
Author Disclosure Statement
No competing financial interests exist.