
Abstract
Population studies have shown a positive correlation between diets rich in whole grains and a reduced risk of developing metabolic diseases, like diabetes, cardiovascular disease, and certain cancers. However, little is known about the mechanisms of action, particularly the impact different fermentable components of whole grains have on the human intestinal microbiota. The modulation of microbial populations by whole grain wheat flakes and the effects of toasting on digestion and subsequent fermentation profile were evaluated. Raw, partially toasted, and toasted wheat flakes were digested using simulated gastric and small intestinal conditions and then fermented using 24-hour, pH-controlled, anaerobic batch cultures inoculated with human feces. Major bacterial groups and production of short-chain fatty acids were compared with those for the prebiotic oligofructose and weakly fermented cellulose. Within treatments, a significant increase (P<.05) in bifidobacteria numbers was observed upon fermentation of all test carbohydrates, with the exception of cellulose. Toasting appeared to have an effect on growth of lactobacilli as only fermentation of raw wheat flakes resulted in a significant increase in levels of this group.
Introduction
D
Wheat can be rolled and flaked and added to breakfast cereals as a raw component, but it is often toasted to improve taste and texture. Despite the fact that most components of a typical Western diet are heat-processed, very little is known about the impact of different food processing procedures on the digestion and fermentation of whole grain wheat flakes. The Maillard reaction is a complex set of reactions that occur between food protein and sugar during thermal processing. These reactions result in fusion of macromolecules like carbohydrates and proteins that are present in complex foods and increase their resistance to digestion in the small intestine, therefore increasing persistence into the colon where they become available for microbiota fermentation. Not all Maillard reaction products are biologically active; nevertheless, the toasting process required for many wheat-based breakfast cereals may affect the composition of molecules that can be fermented and therefore alter the impact of whole grain wheat on bacterial population levels within the colon.
In the present study we used pH-controlled, fecal batch cultures as an in vitro mixed culture system to investigate the microbiota-modulating capabilities of whole grain wheat flakes toward a more beneficial composition, often seen as increased numbers of bifidobacteria and/or lactobacilli. Additionally, we measured the impact toasting has on digestion and the subsequent fermentation of wheat flakes. This is a novel investigation that may have implications for the optimal design of cereal-based foods targeting intestinal health.
Materials and Methods
Substrates
Raw wheat flakes were prepared from flaking fresh pure wheat grain, undamaged and unsprouted, that had been dehulled, steamed, and then dried on a fluid bed dryer. The toasted wheat flakes were prepared by feeding the raw wheat flakes onto an oven belt and baked up to approximately 200°C until fully toasted. The partially toasted wheat flakes were prepared by removing them from the oven before they had become fully toasted. The samples, supplied from Jordans Cereals (Biggleswade, United Kingdom) and prepared from the same crop, contained the following (per 100 g): for raw wheat flakes, 64.4 g of carbohydrates, of which 0.4 g was sugars, 11.8 g was protein, 1.9 g was fat, and 10.4 g of fiber; for partially toasted wheat flakes, 64.4 g of carbohydrates, of which 0.4 g was sugars, 11.8 g was protein, 1.9 g was fat, and 10.4 g was fiber; and for toasted wheat flakes, 82.4 g of carbohydrates, of which 1.7 g was sugars, 10.3 g was protein, 1.9 g was fat, and 12.9 g was fiber. Two control samples were included in the in vitro fermentation experiment: oligofructose (Raftilose® P95, Orafti N.V., Oreye, Belgium) and cellulose (Oxoid, Cambridge, United Kingdom).
Simulated human digestion
Simulated digestion was performed essentially as described by Mills et al. 16 In brief, sterile distilled water (600 mL) was added to 240 g of wheat flake sample, and the mixture was stomached for 5 minutes. The sample solution was transferred to a glass screw-topped bottle, mixed with 25 mL of CaCl2 (1 mM, pH 7.0) containing 78 mg of α-amylase, and incubated at 37°C for 30 minutes. The pH was decreased to 2.0 with 6 M HCl, and pepsin (10.8 g) dissolved in HCl (0.1 M; 100 mL) was added. The sample was incubated at 37°C for 2 hours. The pH was increased to 7.0 with 6 M NaOH, and 500 mL of 0.5 M sodium bicarbonate containing 2.25 g of pancreatin and 14.04 g of bile was added. The sample was incubated at 37°C for 3 hours. The sample solution was transferred to 1-kDa molecular weight cutoff dialysis tubing and dialyzed for 2 hours against NaCl (10 mmol/L) at 4°C to remove low-molecular-mass digestion products. The dialysis fluid was changed, and dialysis was continued overnight and for an additional 2 hours the next day again after the dialysis fluid was changed. Finally, the sample was freeze-dried for 7 days to remove all liquid content.
Fecal batch culture fermentation
The fermentation profile of the freeze-dried digested wheat flake samples, the prebiotic oligofructose, and the weakly fermented control carbohydrate cellulose was determined using anaerobic, pH-controlled fecal batch cultures. Sterile stirred batch culture fermentation vessels (200 mL working volume) were prepared and aseptically filled with 180 mL of sterile basal nutrient medium. Fermentation by human fecal bacteria with the carbohydrate as the substrate was conducted in triplicate using three different healthy fecal donors on each separate assay performance. All donors were healthy men between 23 and 31 years old who had not received antibiotic treatment for at least 3 months, had not consumed pre- or probiotic supplements prior to experimentation, and had no history of bowel disorders. Basal medium contained, per liter, 2 g of peptone, 2 g of yeast extract, 0.1 g of NaCl, 0.04 g of K2HPO4, 0.04 g of KH2PO4, 0.01 g of MgSO4·7H2O, 0.01 g of CaCl2·6H2O, 2 g of NaHCO3, 2 mL of Tween 80, 0.05 g of hemin dissolved in 1 mL of 4 M NaOH, 10 μL of vitamin K, 0.5 g of
Culture-independent enumeration of fecal bacteria using 16S rRNA probes labeled with Cy3 and fluorescence in situ hybridization
Fecal batch culture samples (375 μL) were fixed on removal from the batch culture vessels using ice-cold 4% paraformaldehyde (pH 7.2) at a ratio of 1:3 (vol:vol) in a 1.5-mL Eppendorf tube and were stored at 4°C for 4 hours. The samples were then centrifuged at 13,000 g for 5 minutes and washed twice by resuspending the pellet in 1 mL of filtered (polyvinylidene difluoride filter; pore size, 0.22 μm) phosphate-buffered saline and centrifuging. The remaining pellet was resuspended in a filtered, sterilized phosphate-buffered saline/ethanol mix (1:1, vol:vol) and stored at −20°C for up to 3 months. Microbial populations were enumerated as previously described by Daims et al., 17 using fluorescently labeled 16S rRNA-targeted oligonucleotide probes and fluorescent in situ hybridization. Slides were enumerated using a Nikon (Tokyo, Japan) microscope (×1,000) fitted with an epifluorescence attachment; 15 randomized views were counted for each sample.
The hybridization was carried out as previously described by Rycroft et al.
18
using genus- and group-specific 16S rRNA gene-targeted oligonucleotide probes labeled with Cy3 (Sigma-Aldrich, St. Louis, MO, USA) or the nucleic acid stain 4',6-diamidino-2-phenylindole for total cell counts. A 20-μL volume of each sample was pipetted onto Teflon® (Dupont, Wilmington, DE, USA)- and poly-
Quantification of short-chain fatty acids by gas chromatography
Gas chromatography was performed to determine a change in short-chain fatty acid (SCFA) concentration during the fermentation of the cereal digests; the method was adapted from that of Zhao et al. 24 One milliliter of batch culture was centrifuged at 13,000 g for 10 minutes, and the supernatant was removed to a fresh 1.5-mL plastic tube, which was stored at −20°C for up to 3 months. An internal standard of ethylbutyric acid was prepared with high-performance liquid chromatography gradient water to give a final concentration of 10 mM. For preparation of the external standard containing the SCFAs, acetic, propionic, butyric, isobutyric, valeric, isovaleric, and caproic acids were added to give a final concentration of 25 mM. Dilutions of the external standards were prepared and added to the internal standard (ratio 4:1) to give a final concentration for the internal standard of 2 mM ethylbutyric acid and a final concentration of external standards as 20 mM, 10 mM, 5 mM, 1 mM, and 0.5 mM. Samples were defrosted, and HCl (6 M) was added (ratio 1:3) and left at room temperature for 10 minutes. Samples were centrifuged at 13,000 g for 5 minutes, and the supernatant was passed through a polyvinylidene difluoride filter (pore size, 0.2 μm) (Whatman, Maidstone, United Kingdom). Internal standard was added to the filtered supernatant (ratio 1:4). Samples were run through a 5890 SeriesII gas chromatography system (Hewlett Packard, Palo Alto, CA, USA) fitted with an FFAP column (30 m×0.53 mm i.d.; film thickness, 0.50 μm; J&W Scientific, Folsom, CA, USA) and a flame-ionization detector. Samples were automatically injected into a fused-silica capillary column. Carrier gas (helium) was delivered at a flow rate of 14 mL/minute, and the split ratio was 10:1. Total flow rate was 140 mL/minute. Initial oven temperature was 100°C, maintained for 0.5 minute, raised to 150°C at 8°C/minute, then increased to 250°C at 50°C/minute, and held at 250°C for 2 minutes. Injector and detector temperatures were set at 280°C and at 300°C, respectively. The injected volume of the sample and standards was 1 μL. The SCFA peaks were integrated using Atlas Lab managing software (Thermo Lab Systems, Waltham, MA, USA), and concentrations were calculated by comparing their peak areas with those of the standards and were expressed as micromoles per liter of feces.
Scanning electron microscopy
The wheat flakes were mounted on an aluminum platform with a conductive adhesive and coated with gold, using a sputter coater. Scanning electron microscopy images were obtained at an optimum magnification (×1,000 and ×500), the endosperm portion was scanned, and the selected portions were photographed. All scanning electron microscopy images were obtained at an accelerating voltage of 25 kV using an electron microscope (model Quanta 600 F, FEI™, Hillsboro, OR, USA).
Statistical analysis
Statistical analysis was performed using Minitab for Windows, version 15 (Minitab Inc., State College, PA, USA). Univariate analysis of variance and the post hoc Tukey's test were used to determine the significance of the effect of substrates on bacterial group populations and SCFA production. Significant differences for changes in bacterial populations over a 24-hour period after treatment and SCFA ratio at all time points between treatments were also determined. Differences were deemed significant when P<.05.
Results
Growth of human colonic microbiota on test digested substrates
Changes in bacterial populations after 0, 5, 10, and 24 hours of fermentation with the different test substrates are shown in Table 1. No significant changes in total bacteria or bacterial groups enumerated with the phylogenetic fluorescent in situ hybridization probes were observed in the cellulose fermentations. Numbers of total bacteria significantly increased in all fermenters (P<.05) with the exception of cellulose. Significant increases after 24 hours occurred in the Bifidobacterium genus in the presence of raw wheat flakes (P=.02), with numbers increasing from 7.7 log10 cells/mL to 8.5 log10 cells/mL, for partially toasted wheat flakes from 7.8 log10 cells/mL to 8.1 log10 cells/mL (P=.01), for toasted wheat flakes from 7.8 log10 cells/mL to 8.5 log10 cells/mL (P=.02), and for oligofructose from 7.8 log10 cells/mL to 8.4 log10 cells/mL (P=.04). Oligofructose (P=.03) and toasted wheat flakes (P=.006) fermentations resulted in significant increases in the Atopobium cluster enumerated by the Ato291 probe after 5 hours. Raw wheat flake fermentation caused a significant increase in the Atopobium cluster (P=.01), from 7.4 log10 cells/mL to 7.9 log10 cells/mL, in the initial stages (5 hours); this was followed by a significant decrease between 5 hours and 24 hours (P<.001). The toasted wheat flake substrate was the only one to produce a significant increase (P=.04) in Bacteriodes populations, increasing from 8.1 log10 cells/mL to 8.5 log10 cells/mL, after 10 hours of fermentation. The only substrate to result in a significant increase in Lactobacillus–Enterococcus group growth was raw wheat flakes (P=.004), with an increase from 7.4 log10 cells/mL to 7.8 log10 cells/mL after 24 hours. All wheat flake substrates resulted in a significant decrease in the C. histolyticum subgroup after a 10-hour fermentation; however, this was followed by a significant increase after a 24-hour fermentation.
Data are log10 cells/mL of batch culture fluid, in mean (SE) values.
Univariate analysis of variance and Tukey's multivariate comparison tests were used to determine a significant increase/decrease of bacterial populations compared with 0 hour, within each treatment: * P<.05, significant difference from initial value; † P<.05, significant difference from 5-hour value; ‡ P<.05, significant difference from 10-hour value.
Comparison of treatment effects between bacterial populations over a 24-hour fermentation
No significant differences for changes in Bacteriodes, Eubacteria, and total bacterial populations were observed between treatments. Changes in bifidobacteria and clostridia populations compared with cellulose were significantly higher for all test substrates (P<.001); however, no significant difference among oligofructose, wheat flakes, partially toasted wheat flakes, and toasted wheat flakes was observed. The decrease observed in the atopobia population for wheat flake (P=.01) and partially toasted wheat flake (P<.001) fermentation was significantly lower compared with the increase seen for oligofructose. This was also the case when raw wheat flake (P=.003) and partially toasted wheat flake (P<.001) and cellulose (P<.001) were compared with toasted wheat flake. A significant difference between treatments was observed for wheat flake and toasted wheat flake (P=.01) for lactobacilli population changes, with raw wheat flake fermentation significantly increasing this bacterial group whereas toasted wheat flake resulted in a nonsignificant decrease.
SCFA production
Changes in SCFA concentrations after 0, 5, 10, and 24 hours of fermentation with the different test substrates are shown in Table 2. All substrates resulted in a significant increase in total (acetate, propionate, and butyrate combined) SCFA concentration after 10 hours of fermentation by the fecal microbiota. Acetate was the dominant SCFA produced in all fermenters, and a significant increase was observed for cellulose, raw wheat flakes, partially toasted wheat flakes, and toasted wheat flakes after 10 hours of incubation, whereas for oligofructose the concentration of acetate increased significantly after 24 hours. Propionate increased significantly after 10 hours of fermentation for both raw wheat flakes and partially toasted wheat flakes, whereas toasted wheat flakes, cellulose, and oligofructose resulted in significant production only after 24 hours. A significant increase in butyric acid was recorded after 10 hours of fermentation for raw wheat flakes and toasted wheat flakes, and significant production was detected after 24 hours for all other substrates. As expected, levels of all SCFAs increased over the fermentation period for all substrates, and therefore a better indicator is given by the ratios of the individual SCFA to total SCFAs (Fig. 1). There were no significant differences in the proportion of butyrate between the different flakes or controls and no significant difference among the different flakes for any of the SCFA proportions. A significant difference was observed when the larger proportion of acetate after 24 hours of oligofructose fermentation was compared with lower values after raw wheat (P=.004), partially toasted (P=.003), and toasted (P=.006) wheat flake and cellulose (P=.002) fermentation. The ratio of propionate after 24-hour toasted wheat flake fermentation was significantly greater than that after 24-hour cellulose fermentation (P=.04). Also, the ratio of propionate after 10-hour oligofructose fermentation was significantly greater than that after 10-hour cellulose fermentation (P=.03) (Table 3).
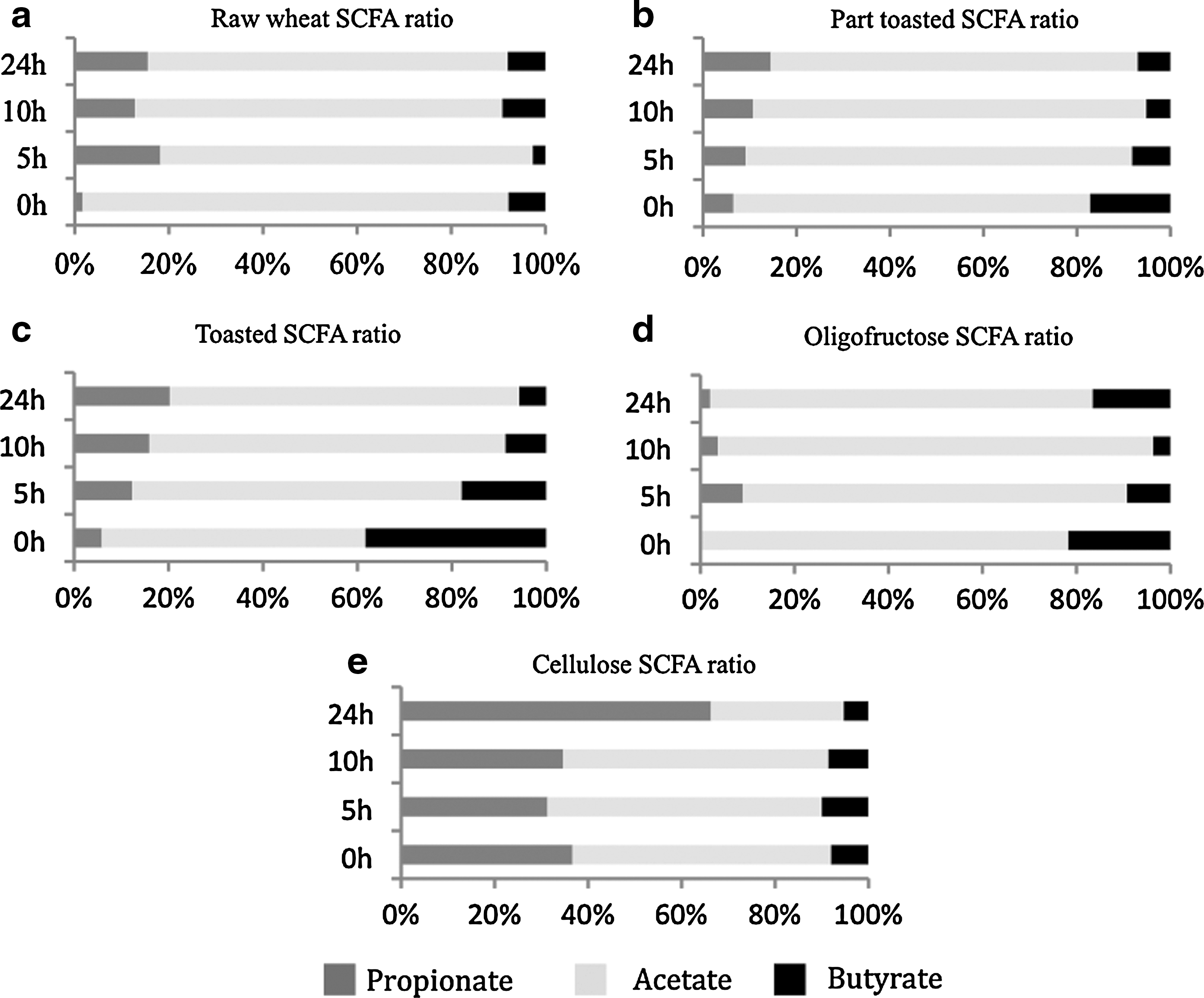
Ratio of SCFA (propionate:acetate:butyrate) production by substrates during a 24-hour fermentation in pH-controlled and stirred batch cultures:
Data are log10 cells/mL of batch culture fluid, in mean (SE) values.
Univariate analysis of variance and Tukey's multivariate comparison tests were used to determine a significant increase in short-chain fatty acid (SCFA) production compared with 0 hour, within each treatment: * P<.05, significant difference from initial value; † P<.05, significant difference from 5-hour value; ‡ P<.05, significant difference from 10-hour value.
Different letters indicate significant differences (P<.05) for each SCFA (i.e., within column) between the different substrates and the time points.
Different letters indicate significant differences (P<.05) for each SCFA (i.e., within column) between the different substrates and the time points.
Scanning electron microscopy
Scanning electron micrographs (Fig. 2) indicate the microstructure of raw, partially toasted, and toasted rolled wheat grain flakes before in vitro digestion. It can be seen that before digestion the raw wheat flakes have a closely packed organized structure, whereas the toasted flakes have a disrupted ridged structure. The partially toasted flakes are seen to have both the initial compact structure in parts and also the disruption of carbohydrate components due to the toasting process. Post-digestion scanning electron micrographs (Fig. 3) again show the raw wheat flakes to have the most compact structure, whereas the digestion of the toasted flakes resulted in small sections of plant cell wall material. The partially toasted flakes are seen to be a mix of the structures.

Scanning electron micrographs of

Scanning electron micrographs of
Discussion
Whole grain cereal consumption has been linked to reduction in risk of the metabolic disorders cardiovascular disease, 25 diabetes, 26 and colon cancer. 27 Population studies have recognized their protective effects, and recently studies have investigated possible mechanisms of action. 6 One possible health-promoting activity of whole wheat grain is its ability to modulate the composition and activity of the human intestinal microbiota. In the present study the effects toasting had on the fermentation profile of wheat flakes in relation to changes of the dominant members of the intestinal microbiota and metabolites produced were determined. The wheat flakes investigated differed in their stage of toasting: raw, partially toasted, and toasted.
The present study has demonstrated the potential for wheat flakes to beneficially modulate the intestinal microbiota, with the observation of significant increases in the Bifidobacterium genus, and the Lactobacillus–Enterococcus group in the case of raw wheat flakes. A significant increase in growth of the Bifidobacterium genus at 24 hours was observed for oligofructose. The significant increase in bifidobacteria and lactobacilli detected corresponds with data reported by Costabile et al., 6 who carried out a human feeding study using a whole wheat grain breakfast cereal. The ability of the wheat flake substrates to promote growth of only the Bifidobacterium genus and Lactobacillus-Enterococcus groups of bacteria was not as great as that of the prebiotic oligofructose, as significant changes were also observed for the C. histolyticum subgroup and the Bacteroides–Prevotella group.
The processing of food adds to the complexity. Heating foods can lead to Maillard reactions between proteins and sugars, which can result in a range of effects, from altering food components' digestibility and possible biological activity and fermentability. 5 Toasting of the flakes appeared to diminish their ability to stimulate the growth of the Lactobacillus–Enterococcus group as only the raw wheat flakes resulted in a significant increase in population. Scanning electron microscopy observations reported by Venkatesh Murthy et al. 28 demonstrated that roasting wheat flakes resulted in changes in the wheat flake structure. The microstructure of the raw wheat flakes had a closed structure with closely packed starch molecules, whereas the toasted wheat flakes had a porous structure. We observed similar results: our scanning electron microscopy analysis of the raw wheat flakes (Fig. 2a) illustrated that initially the starch molecules are compacted together and uniformed. Partially toasting the wheat flakes (Fig. 2b) resulted in the structure breaking up and becoming increasingly disorganized, whereas the fully toasted wheat flakes had no distinguishable starch molecules, and the overall structure was almost completely disrupted and porous (Fig. 2c). The scanning electron microscopy images of the different flakes post-digestion and freeze–drying again demonstrated that the raw flakes (Fig. 3a) contained starch granules and remained a closed structure. The digested partially toasted flakes (Fig. 3b) also showed starch granules present; however, the microstructure was more porous and open compared with the raw flakes. The toasted flakes (Fig. 3c) had no starch granules present, with cell wall structures being the main components present after digestion. In the present study toasting induced changes in flake structure, which may have had consequences on the compounds produced after simulated chewing, pH drop, enzymatic digestions, and bile activity of the simulated digestion process. Indeed, a more porous flake structure may allow easier access of digestive enzymes to the whole grain carbohydrates. This may explain the significant changes in total bacteria populations observed after 5 hours of fermentation for the toasted wheat flakes, which was not observed for the raw or partially toasted wheat flakes (significant changes were observed after 10 hours of fermentation). The toasting may have resulted in increase Maillard reaction products between grain proteins and carbohydrates, which may explain the sustained and significant growth of the Clostridia group and significant increases in Bacteroides, which were only seen for toasted wheat flakes.
Food structure appears to be of general importance in relation to the glucose and insulin response, especially with foods that contain low-molecular-weight carbohydrates as well as starchy foods. It has previously been shown that disruption of the botanical structure of apples, 29 rice, 30 and legumes 31 results in increased availability of the carbohydrate moiety for digestion and absorption. The physical texture of foods has also been reported to play a role in metabolic response. 32 The processing of grains may improve or impair any potential mechanisms associated with improving host health. Processing to increase extractability of β-glucan, which can lead to viscous solution formation in the upper intestine, resulting in a lower metabolic response, is desirable. However, processes that allow increased activity of host digestive enzymes to break down starch (increasing glycemic index), which would otherwise remain resistant, could be considered detrimental. It has been shown in vitro that the composition and concentration of dietary fiber-rich extrudates (e.g., β-glucan and resistant starch) as well as molecular weight of these extrudates (i.e., high-, medium-, and low-molecular-weight β-glucan) can affect binding to bile acid. 33,34 This is of particular interest as bile acid is a major route of cholesterol metabolism, and binding has been proposed to reduce the concentration of plasma cholesterol. 35
Acetate is the most abundant SCFA produced by our colonic microbiota, whereas propionate and butyrate are produced in equal concentrations. 36 Acetate and propionate production by intestinal bacteria has been proposed to have opposing effects in hyperlipidemia (abnormal levels of lipids in the blood), a risk for cardiovascular disease. Studies using animals have suggested that propionate derived from colonic carbohydrate fermentation inhibits cholesterol synthesis in the liver. 37 Wolever et al. 38 investigated this by performing rectal infusion of both acetate and propionate separately (180 mmol) and as a mixed dose (acetate [180 mmol] and propionate [60 mmol]) on healthy human volunteers. Infusion of both acetate and propionate resulted in an attenuated increase in serum cholesterol compared with acetate infusion alone. 39 The studies by Wolever et al. 38,39 support the hypothesis that propionate inhibits cholesterol production by inhibiting acetate utilization. Alvaro et al. 40 demonstrated that propionate down-regulated the expression of key genes involved in intestinal cholesterol biosynthesis after incubation of human Caco-2 enterocytes with propionate. In the present study the acetate, propionate, and butyrate ratio after a 24-hour fermentation for raw wheat flakes was 76:15:9, that for partially toasted wheat flakes was 79:15:6, and that for toasted wheat flakes was 73:20:7, whereas the ratio for oligofructose was 81:2:17. Previous studies have reported variable and inconsistent SCFA profiles after fermentation of whole grain components. Monsma et al. 41 in a previous study used rat cecal inocula to ferment oat bran, which resulted in an SCFA ratio of 69:19:12 for acetate:propionate:butyrate, and concluded this was a high proportion of propionate compared with the SCFA profile generated by wheat bran. This suggests that the production of propionate observed in the present study was through fermentation of components present in the carbohydrate portion of the whole grain, possibly resistant starch. It has been suggested that because of the thick indigestible cell walls of the bran layer, the vitamins and proteins are not easily accessible when ingested by humans. They are therefore an excellent source for microbial fermentation; however, processing the grain will impact on fermentation, which has been shown previously to in turn impact on the SCFA profile. 42 A study carried out by Anson et al. 43 investigated the effect bioprocessing of wheat bran in wholemeal bread had on the colonic SCFA production in vitro. Five different bread samples were included in the study: wheat bread, wholemeal wheat bread, wholemeal wheat bread with native wheat bran, wholemeal wheat bread with fermented wheat bran, and wholemeal wheat bread with fermented and enzyme-treated wheat bran. They reported that processing affected the SCFA profiles after fermentation with a higher proportion of butyrate in exchange for propionates, whereas the total SCFA production remained similar. These results are comparable to the present study, as it can be seen (Fig. 1) that as the concentration of propionate increases or decreases, the butyrate concentration mirrors the change, either increasing or decreasing. Whole wheat bran (large/coarse vs. small/fine) was evaluated to determine the impact on SCFA production using in vitro fermentation. 44 It was reported that the fine bran produced the greatest SCFA concentrations; however, the percentage of butyrate at 24 hours was significantly greater for large-particle bran. This study is again comparable to the findings of the present study in that processing impacts on SCFA profile. In vivo investigations often report no change or nonsignificant changes in SCFA profiles, which is not surprising as they are readily absorbed in the large colon. To date only one study has investigated the impact of feeding a whole grain wheat cereal on microbiota. 6 This group did not detect any changes in SCFA concentrations, most likely for the reason detailed above.
In conclusion, the present study has shown that wheat flake fermentation modulated the intestinal microbiota in a beneficial manner. Significant growth of members of the Bifidobacterium genus and, in the case of raw wheat flakes, the Lactobacillus–Enterococcus group was observed. However, the bacterial selectivity was lower than that observed for the prebiotic oligofructose. A relationship between processing and its subsidiary effects on the colonic microbiota fermentation profile has been identified. The present work confirms that food processing can impact on the fermentation profile of whole grain cereals and on the ability to stimulate beneficial members of the intestinal microbiota, an important consideration to the design of optimal functional foods targeting intestinal health.
Footnotes
Acknowledgments
This study was supported by the Food Microbial Sciences Unit at University of Reading, Reading, United Kingdom, and The Jordans & Ryvita Company Ltd., Biggleswade, United Kingdom.
Author Disclosure Statement
No competing financial interests exist.