
Abstract
Pomegranate (Punica granatum L.) is known to possess pharmacological activities, such as antioxidant and anticancer. In this study, we evaluated the antioxidant potency of a methanolic pomegranate fruit peel extract (PPE) and the relation with its antiproliferative and apoptotic effects on MCF-7 human breast cancer cells. Total phenolic content and antioxidant activity of PPE were determined using the Folin–Ciocalteau and the 2,2-diphenyl-l-picrylhydrazyl free radical methods, respectively. Phenolic acids present in the extract were characterized by a reverse-phase high-performance liquid chromatography (HPLC) method. Cell proliferation was assessed by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide reduction assay. The apoptotic effects were determined by in situ Tdt-mediated dUTP nick end-labeling assay, and Bax/Bcl-2 mRNA expression levels were measured by reverse transcription–polymerase chain reaction. The extraction yield as a percentage of plant material was 37.97% (wt/wt), and total phenolic content was 331.28 mg of gallic acid equivalents/g of extract. According to HPLC analysis, the most abundant phenolic acid detected in the extract was ellagic acid. MCF-7 cell proliferation decreased depending on PPE concentration (25, 50, 100, 200, and 300 μg/mL) and incubation times (24, 48, and 72 hours). After 48 and 72 hours, the apoptotic cell numbers were significantly increased at 100, 200, and 300 μg/mL PPE concentrations. In addition, expression of the pro-apoptotic gene Bax was increased, and that of the anti-apoptotic gene Bcl-2 was decreased after 200 and 300 μg/mL PPE treatment for 48 and 72 hours. Because PPE reduced cell proliferation and induced apoptosis on MCF-7 cancer cells, we believe that PPE has important antioxidant and apoptotic effects.
Introduction
B
Pomegranate (Punica granatum L.) is a rich source of many phenolic compounds including flavonoids, anthocyanins, catechins, other complex phenolics, and hydrolyzable tannins (punicalin, pedunculagin, punicalagin, gallagic, and ellagic acid esters of glucose). 6 Pomegranate fruit peel extract (PPE) with an abundance of flavonoids and tannins exhibits a high antioxidant activity. 7 –9 It has been shown that consumption of phytochemical-rich compounds such as pomegranates can reduce the risk of cancer. Flavonoid-rich polyphenol fractions from the pomegranate fruit exert antiproliferative, anti-invasive, and pro-apoptotic actions and anti-angiogenic activities. 10 Pomegranate peel contains substantial amounts of polyphenols such as ellagic acid and gallic acid. Recently studies have demonstrated that pomegranate fruit extract possesses remarkable antitumor-promoting effects in some cancer cells. 2,10 –12 Kim et al. 1 have reported that in both MCF-7 and MB-MDA-231 breast cancer cell lines, fermented pomegranate juice polyphenols consistently showed about twice the antiproliferative effect as fresh pomegranate juice polyphenols; in addition, pomegranate seed oil inhibited proliferation of MCF-7 cells and invasion across Matrigel™ (BD Biosciences, Franklin Lakes, NJ, USA) and resulted in apoptosis in MDA-MB-435 estrogen receptor–negative metastatic human breast cancer cells.
In this study, we evaluated the antioxidant potency of methanolic PPE and the relation with its antiproliferative and apoptotic effects on MCF-7 cells.
Materials and Methods
Extraction
Pomegranate fruits were obtained from Antalya, Turkey. Fresh pomegranate fruits were peeled and dried. Dried fruit peels were extracted with petroleum ether in a Soxhlet apparatus. Fat-free air-dried material was extracted with methanol in the same apparatus. The methanol extract was concentrated to dryness under vacuum. All extracts obtained were weighed to determine the yields of soluble constituents. 13
Determination of total phenolic content (Folin–Ciocalteu method)
The amounts of total phenols were determined by spectrophotometrically at 750 nm based on the absorbance measurement for extracts in the form of gallic acid as described in a previous method. 14 This method gives a general measurement of phenolic content, as it is not completely specific for phenolic compounds and not all phenolic compounds exhibit the same level of activity in the assay.
Free radical scavenging activity using the 2,2-diphenyl-l-picrylhydrazyl method
The 2,2-diphenyl-l-picrylhydrazyl (DPPH) assay was used as a rapid spectroscopic method to provide an evaluation of antioxidant activity due to scavenging free radicals. Being a stable free radical with purple color, DPPH• is reduced to the yellow-colored diphenylpicryl hydrazine. Free radical scavenging effects of PPE on DPPH• were estimated according to the method of Sanchez-Moreno et al. 15 with some modifications. The reaction mixture was left at ambient temperature for 30 minutes in darkness; absorbance of the resulting solution was then measured spectrophotometrically at 517 nm. The scavenging activity was plotted against concentration, and the concentration that showed 50% DPPH scavenging activity was calculated following the logarithmic procedures.
High-performance liquid chromatography analysis
A high-performance liquid chromatography (HPLC) system consisting of a model 600 E HPLC pump, model 717 Plus autosampler, model 996 photodiode array detector, and Millennium 32 data processor was used (Waters Corp., Milford, MA, USA). Ultrapure water (18.2 μS/cm) from a Millipore (Molsheim, France) water purification system was used. The HPLC analysis of phenolic acids was performed on a reverse-phase C18 Ultrasphere column (Teknokroma, Barcelona, Spain) (100×4.6 mm inner diameter; particle size, 3 μm) by using a gradient program with a two-solvent system (A, methanol:water:formic acid, 10:88:2 by volume; B, methanol:water:formic acid, 90:8:2 by volume) as reported previously. 16 The flow rate was always 1 mL/minute, and the injection volume was 10 mL. The signals were detected at 280 nm. The internal standard technique was applied to the analysis to increase the repeatability. Propylparaben is a suitable internal standard for this system.
Cell culture and treatment
MCF-7 human breast adenocarcinoma cells were provided by Dr. Yusuf Baran, Department of Molecular Biology and Genetics, Izmir Institute of Technology, Izmir, Turkey. The cells were grown in RPMI 1640 medium supplemented with 2 mM
Cytotoxicity/cell proliferation assay
Proliferation of the cells was assessed by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, which is based on the reduction of MTT by the mitochondrial dehydrogenase of intact cells to a purple formazan product. Yellow MTT is reduced to purple formazan in the mitochondria of living cells. This reduction takes place only when mitochondrial reductase enzymes are active, and therefore conversion can be directly related to the number of viable (living) cells. 17 Cells were inoculated into 96-well culture plates at densities of 5×103 cells per well. After 24 hours, they were treated with PPE (concentrations of 25, 50, 100, 200, and 300 μg/mL) for 24, 48, and 72 hours. After the incubations, MTT solution (5 mg/mL) was added to each well and incubated for 3 hours at 37°C. At the end of the incubations, the purple MTT–formazan crystals were dissolved by adding 100 μL of dimethyl sulfoxide to each well. The absorbance of the samples was measured with an enzyme-linked immunosorbent assay reader (wavelength of 570 nm). In the experiment, each group was tested in eight wells. The data are mean values from three different experiments. The effect of PPE on the proliferation of cancer and normal cells was expressed as relative cell viability, using the following formula: Percentage cell proliferation=(optical density of drug-treated sample/optical density of untreated sample)×100. 18,19
In situ Tdt-mediated dUTP nick end-labeling assay
Tdt-mediated dUTP nick end-labeling (TUNEL) was performed by using an Apop Tag Plus apoptosis detection kit (catalog number-S7101 in situ apoptosis detection kit, Chemicon International, Hofheim, Germany). DNA fragmentation characteristic of apoptotic cells was quantified by TUNEL according to the manufacturer's instructions. 20 MCF-7 cells were grown on coverslips and treated with PPE for 24, 48, and 72 hours. Cells were then analyzed for apoptosis with the TUNEL assay. Dark-blue staining of nuclei indicates apoptosis. To calculate the percentage of TUNEL-positive cells, five random microscopic fields at ×400 magnifications were assessed.
RNA extraction
MCF-7 cells were seeded in six-well plates, in which each well contained 1×106 cells, and they were allowed to incubate for 24 hours. At the end of the incubation period, 25, 50, 100, 200, and 300 μg/mL concentrations of PPE were applied onto MCF-7 cells in fresh medium solution, and plates were incubated in an incubator with 5% CO2 at 37°C for 48 and 72 hours. After incubations, total cellular RNA was extracted for each treatment using a BDtract™ RNA isolation kit (Maxim Biotech. Inc., Rockville, MD, USA) according to the manufacturer's protocol. Quantification and purification of the isolated RNA were performed by determining the ratio of absorbance at 260 nm to absorbance at 280 nm using a spectrophotometer. 21,22
Reverse transcriptase–multiplex polymerase chain reaction of Bax and Bcl-2 mRNA expressions
Expression of Bax and Bcl-2 genes was analyzed by reverse transcription (RT)–polymerase chain reaction (PCR) amplification in contrast with the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase. Therefore, cDNA was synthesized from 10 μg of total RNA using the Ready-to-use First Strand cDNA Synthesis kit (Maxim Biotech, Inc.). Then, equal volumes of cDNA were amplified by PCR using a multiplex PCR kit for Human Apoptotic Genes Set-2 (Maxim Biotech, Inc.) according to the protocol instructions. 23 The multiplex PCR thermal cycler conditions were as follows: initial denaturation at 96°C for 1 minute (one cycle), followed by two cycles of denaturation at 96°C for 1 minute and primer annealing at 70°C for 4 minutes, and 35 cycles of denaturation at 94°C for 1 minute and primer annealing at 70°C for 2 minutes. Final extension was at 70°C for 10 minutes, followed by soaking at 4°C. A TaKaRa Dice™ (Takara Bio Inc., Shiga, Japan) instrument was used as the thermal cycler. PCR products were electrophoresed on a 2% agarose gel after ethidium bromide staining, imaged by using a UVP (Cambridge, United Kingdom) EC3 bioimaging system, and analyzed by VisionWorks®LS Image Acquisition and Analysis software (UVP). Differences in gene expression were determined by normalizing its expression against expression of glyceraldehyde 3-phosphate dehydrogenase (housekeeping gene; internal standard included in the multiplex PCR kit). We analyzed human Bax (272 bp), Bcl-2 (235 bp), and glyceraldehyde 3-phosphate dehydrogenase (500 bp) mRNA gene expression.
Statistical analysis
All data were recorded as mean±SEM values. Results were analyzed by one-way analysis of variance plus Tukey's test. P values of <.05 were considered significant (*P<.05, **P<.01, ***P<.001).
Results
Determination of antioxidant activities of PPE
The extraction yield as a percentage of plant materials was 37.97% (wt/wt), and total phenolic content was found to 331.28±9.05 mg of gallic acid equivalents/g of extract (dry weight). The values are given as means of triplicate analyses. The relatively stable organic radical DPPH• has been widely used in the determination of the antioxidant activity of pure compounds. Free radical scavenging potential of the extract was also tested by the DPPH method in comparison with that of a synthetic antioxidant, tert-butyhydroxytoluene, at different concentrations. The PPE at the three different concentrations investigated (5 (g/mL, 10 (g/mL, and 20 (g/mL) showed dose-dependent radical scavenging activities. Percentage inhibition value is a parameter widely used to measure the free radical scavenging activity. That is, a higher percentage inhibition value corresponds to a higher antioxidant activity. 24 These data indicate that PPE had higher activity than tert-butyhydroxytoluene at the same concentrations (Fig. 1A).
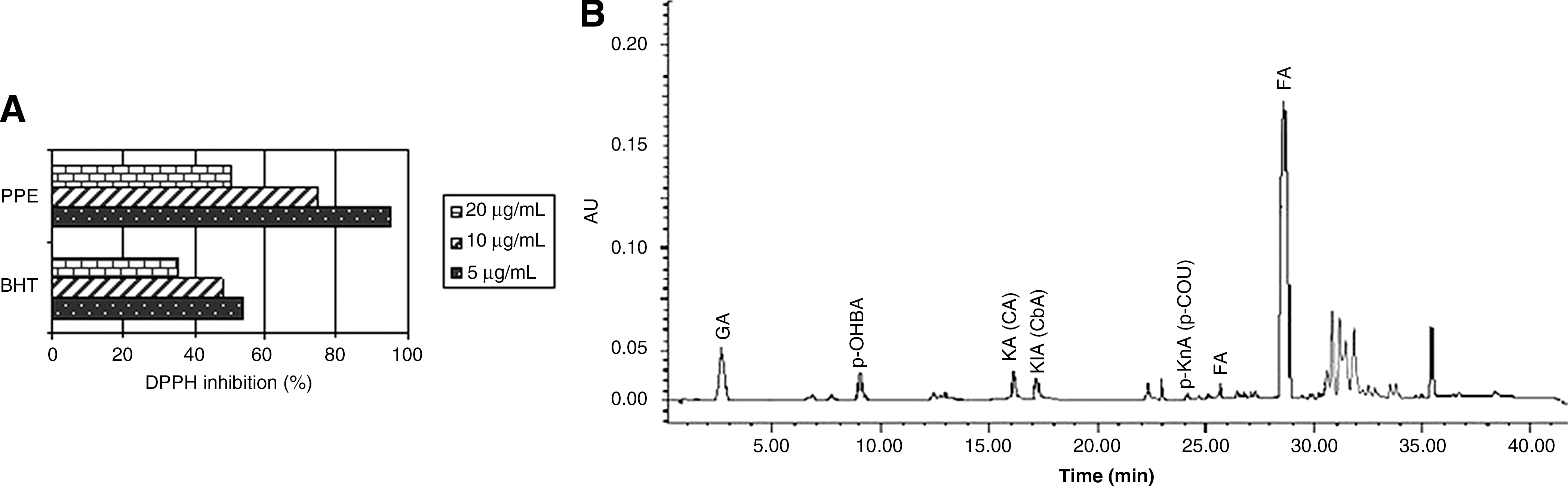
(
HPLC analysis
Most of the phenolic acids have absorption maxima in the ultraviolet absorption spectra at a wavelength of 280 nm, and they were identified by matching their retention times (peak normalization) and ultraviolet spectra of samples with those of authentic standards, using the HPLC–photodiode array detection system. All of the phenolic acids were resolved entirely from each other (Fig. 1B). In addition, the phenolic acid amounts in PPE are given in Table 1. The results show that ellagic acid dominated in the extract. Ellagic acid was followed by gallic acid, p-hydroxybenzoic acid, caffeic acid, and chlorogenic acid. Our results also are consistent with the findings of the literature. 25,26
Cytotoxicity/cell proliferation assay
Morphological differentiations of MCF7 cells after 72 hours of treatment with the different concentrations of PPE are shown in Figure 2. Decreasing cell proliferation was observed with increasing PPE concentration.
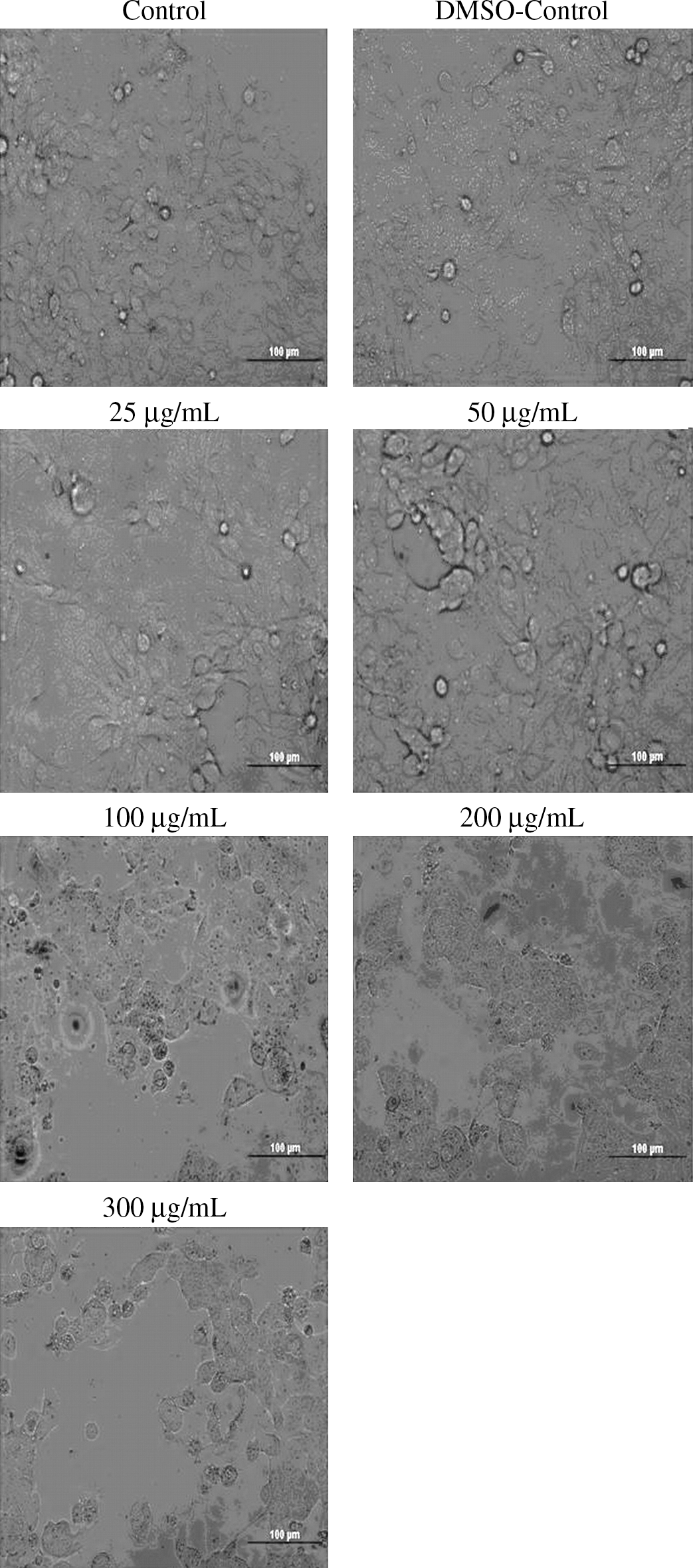
Morphological differentiations of MCF-7 cells after incubation for 72 hours with different concentrations of PPE. Cells were photographed under a fluorescent inverted microscope (Olympus). Magnification ×400. DMSO, dimethyl sulfoxide.
Effects of PPE on MCF-7 cell proliferations were determined with the MTT reduction assay (Fig. 3). Using the MTT reduction assay, percentage cell proliferations with the different PPE concentrations were calculated for the control group at the end of incubation for 24, 48, and 72 hours. The decrease in MCF-7 cell proliferation was statistically apparent with treatment with PPE. For the solvent control group (dimethyl sulfoxide control) the percentage cell proliferation value was 98.7% (P>.05). Percentage cell proliferations for the control group with 25, 50, 100, 200, and 300 μg/mL PPE concentrations after a 24-hour incubation were found to be 96.4% (P>.05), 95.3% (P>.05), 94.5% (P<.05), 90.1% (P<.05), and 86.6% (P<.01), respectively. The cell proliferation values at 48 hours with increasing PPE concentration were calculated to be 85.2% (P<.01), 84.5% (P<.001), 81.6% (P<.001), 75.9% (P<.001), and 69.2% (P<.001), respectively. At the end of the 72-hour incubation of MCF-7 cells with the different PPE concentrations, the cell proliferation values, depending on the concentration increase, were 70.8%, 70.5%, 67.7%, 57.1%, and 48.6% (P<.001), respectively (Fig. 3). In parallel with the PPE concentration and increase of the incubation time, substantial decreases occurred in MCF-7 cell proliferation. The most significant decreases in cell proliferation were particularly observed after incubations with 200 and 300 μg/mL PPE concentrations for 48 and 72 hours. Calculated from the effects of PPE concentrations on MCF-7 cells, the 50% DPPH scavenging activity value after a 72-hour incubation was 274.15 μg/mL.
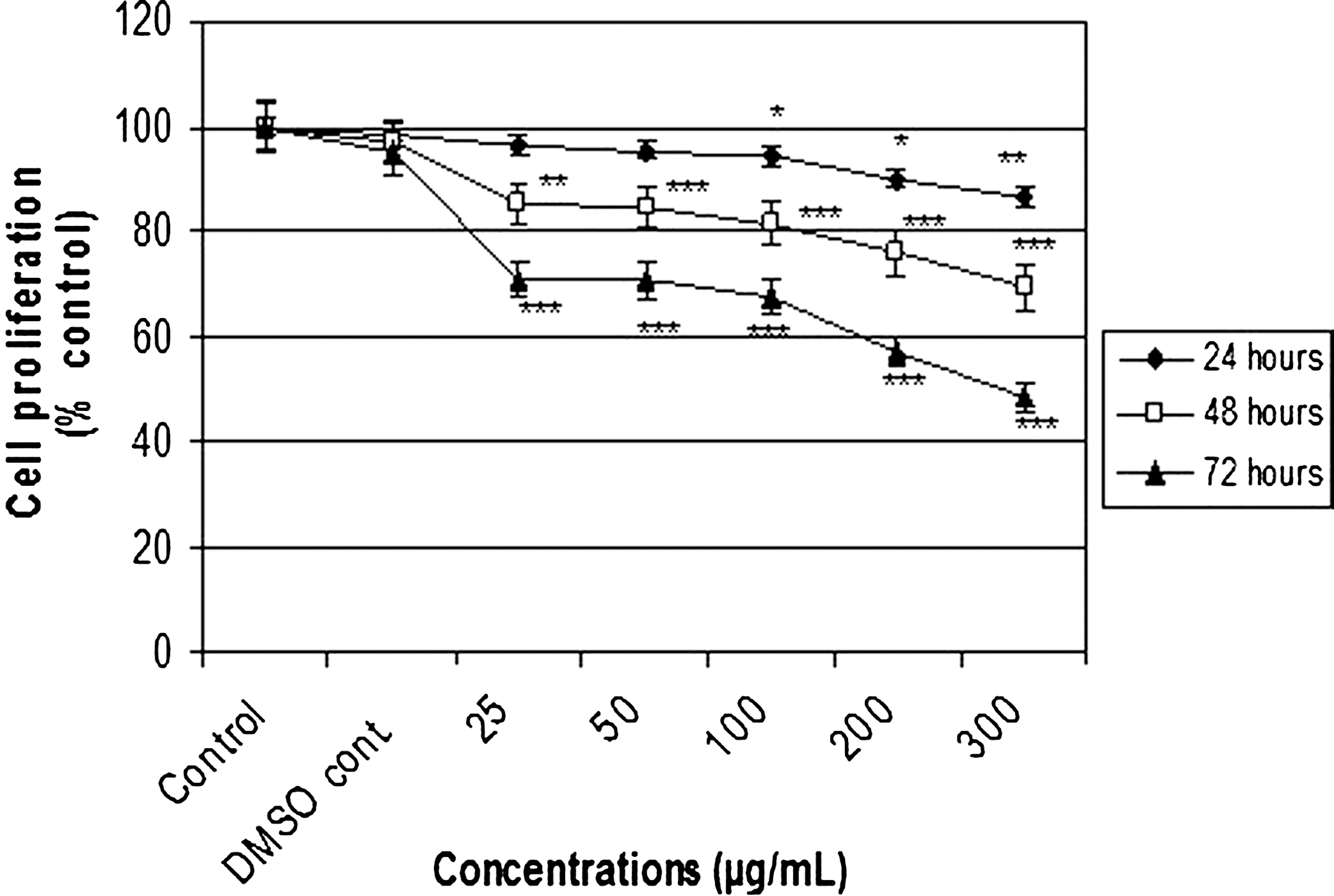
Effects of different concentrations of PPE on MCF-7 cell proliferations estimated by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reduction assay. Cells were treated with nothing (control, only medium), vehicle (DMSO, 0.1% in medium), or different concentrations of PPE for 24, 48, and 72 hours. MTT reduction was then calculated to indicate cellular proliferation. Data are mean±SEM values (n=8). *P<.05, **P<.01, ***P<.001, significantly different from control values.
Apoptotic effects (TUNEL assay and Bax/Bcl-2 mRNA expressions)
After the TUNEL assay, the light microscopic images of MCF-7 cells incubated with different concentrations of PPE can be seen in Figure 4. Apoptotic cells were colored reddish brown, so that they can be distinguished from the normal cells. We determined that PPE caused an increase in apoptotic cell percentage depending on incubation interval and PPE concentration on MCF7 cells using the TUNEL assay. There was a significant increase particularly in the apoptotic cell number after 48 and 72 hours with 100, 200, and 300 μg/mL PPE concentrations. Percentages of apoptotic cells after 72 hours were found to be higher than those after 48 hours. At the end of a 48-hour incubation, percentages of apoptotic cells were calculated to be 1.23% in the control group, 2.84% (P<.05) with 100 μg/mL extract, 3.03% (P<.01) at 200 μg/mL, and 4.57% (P<.001) at 300 μg/mL. At the end of a 72-hour incubation, the apoptotic cell percentage of the control group was 1.40%, and the value increased to 3.85% (P<.01), 4.66 (P<.001), and 5.98 (P<.001) with 100, 200, and 300 μg/mL extract concentrations, respectively. No statistical difference was found between the control and dimethyl sulfoxide control groups in terms of the cell percentages at the end of each incubation (P>.05) (Table 2).
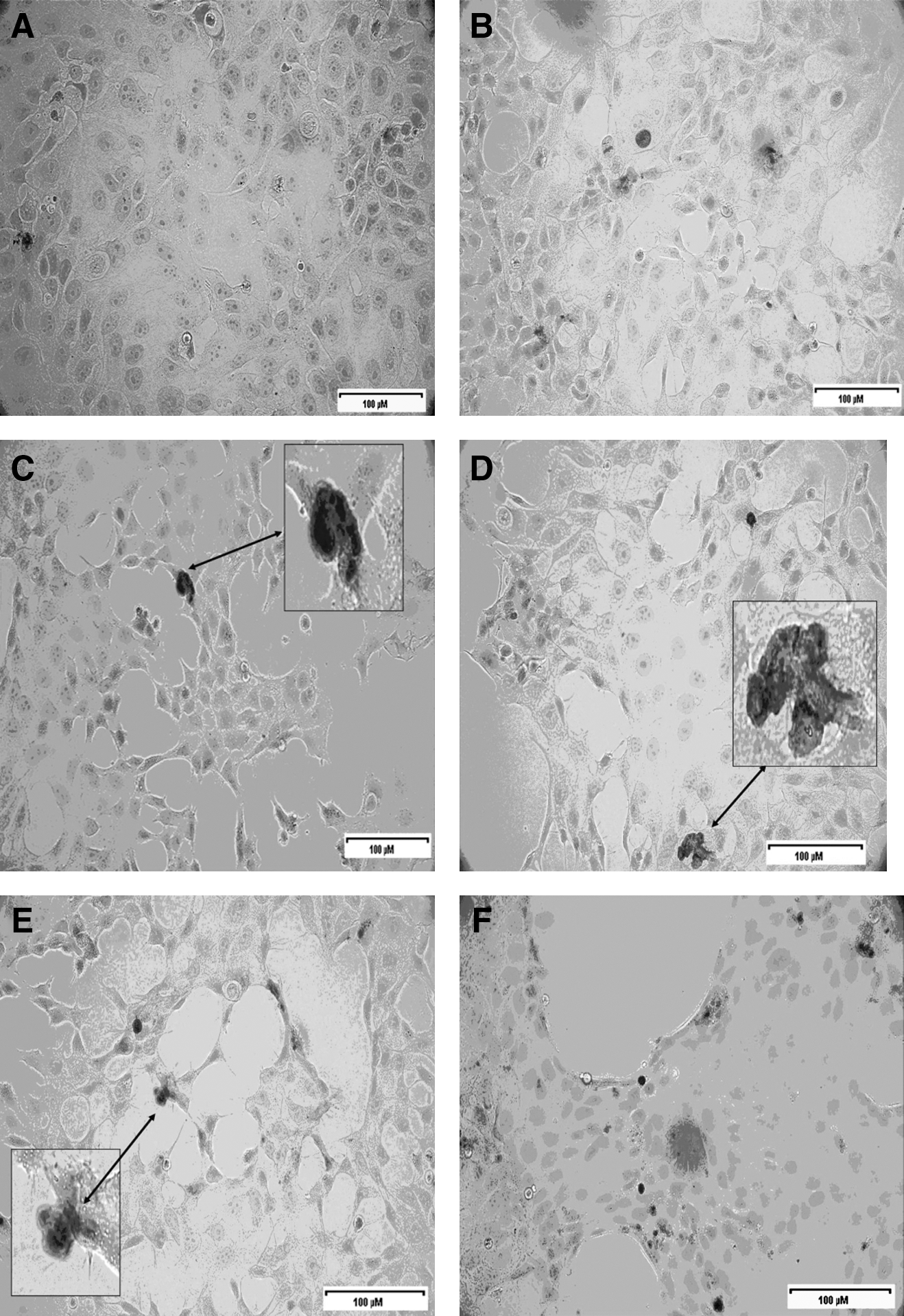
Tdt-mediated dUTP nick end-labeling staining of MCF-7 cells treated with different PPE concentrations: (
P values of <.05 were considered significantly different from control values: for 24 hours, a versus g (P<.01), b versus g (P<.05); for 48 hours, a versus e (P<.05), a versus f (P<.01), a versus g (P<.001), b versus e (P<.05), b versus f (P<.05), b versus g (P<.001), c versus f (P<.05), c versus g (P<.001), d versus f (P<.05), d versus g (P<.001), e versus g (P<.01), f versus g (P<.05); and for 72 hours, a versus e (P<.01), a versus f (P<.001), a versus g (P<.001), b versus e (P<.01), b versus f (P<.001), b versus g (P<.001), c versus e (P<.01), c versus f (P<.001), c versus g (P<.001), d versus f (P<.001), d versus g (P<.001), e versus g (P<.01).
In parallel with the results of the TUNEL assay, after RT-PCR, the agarose gel images and densitometric analysis results of MCF-7 cells incubated with different concentrations of PPE for 48 and 72 hours are shown in Figure 5. Markers at 100 bp were used in agarose gel images. Genes areas such as those for Bax (272 bp), Bcl-2 (235 bp), and glyceraldehyde 3-phosphate dehydrogenase (500 bp; housekeeping gene; internal standard), depending on the features of the primers used in the PCR process, were amplified by PCR. Bax and Bcl-2 mRNA expressions were calculated in proportion to glyceraldehyde 3-phosphate dehydrogenase expression densitometrically. Although Bax and Bcl-2 mRNA expressions had similarities in terms of agarose gel images after 48- and 72-hour incubations with different PPE concentrations in MCF-7 cells, increases in the rates of Bax/Bcl-2 mRNA expressions were detected densitometrically following incubations with both 200 and 300 μg/mL PPE concentrations relative to the control group. This finding supports the increase of the apoptotic cell number with 200 and 300 μg/mL PPE concentrations after 48 and 72 hours as measured by the TUNEL method.
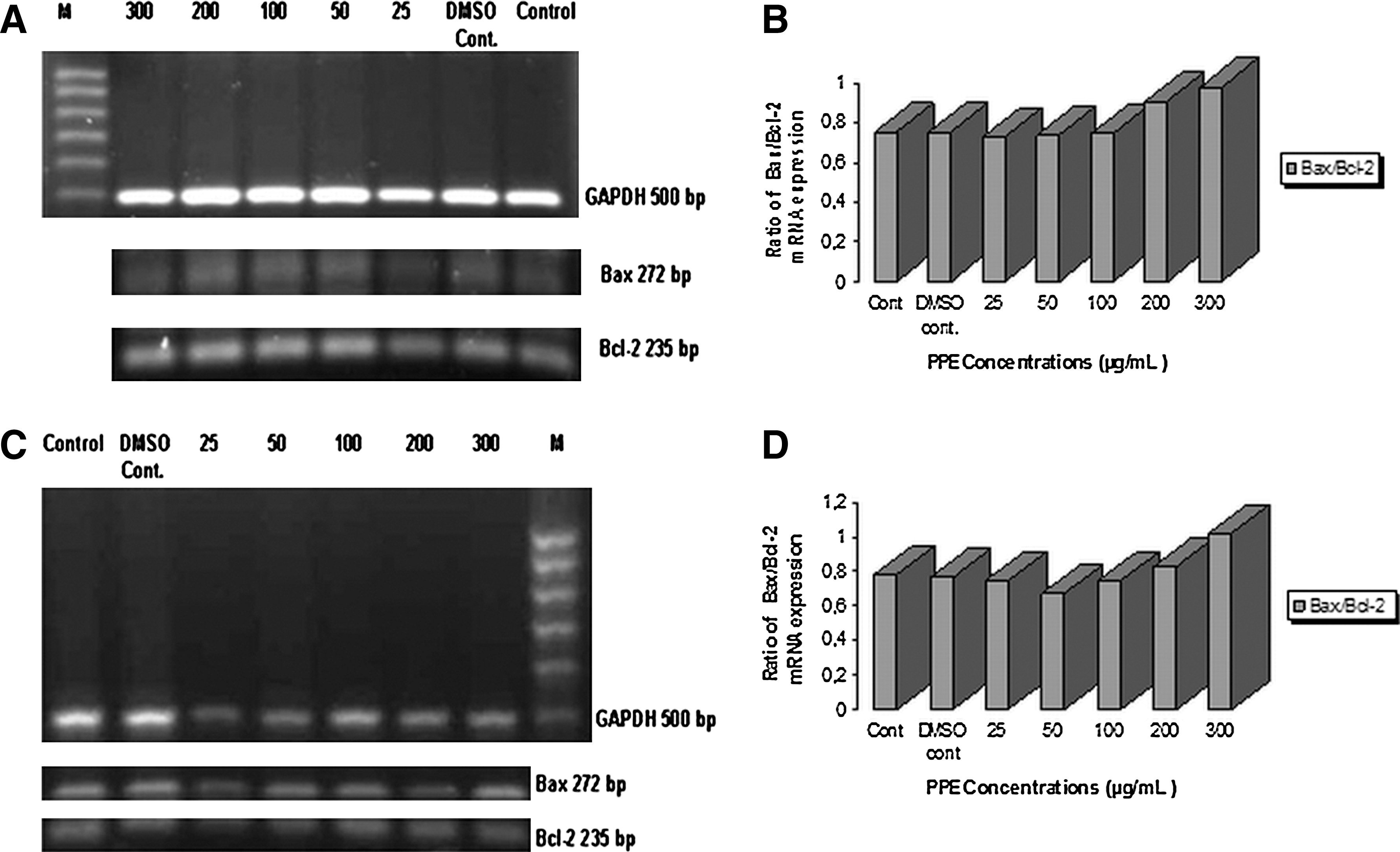
mRNA expression of Bcl-2 and Bax in MCF-7 cells after treatment with various concentrations of PPE for (
Discussion
Pomegranate, its juice, and extracts have been used for treatment of diseases since the ancient times. Because it is high in antioxidants and other nutrients, some people think that drinking pomegranate juice regularly may help prevent cancer. Some in vitro and in vivo studies have revealed that the pomegranate has powerful antioxidant properties because of the presence of polyphenols and inhibits the growth of cancer cells. 19,27 –29 Pomegranate has some phenolic groups such as punicalagins, ellagic acid, gallotanin, anthocyanins, and tannins that are able to be hydrolyzed. 27,30,31 It has been reported by Noda et al. 31 that the juice of pomegranate also has anthocyanins (delphinidin, cyanidin, and pelargonidin). Another study has stated that polyphenols are the active substances of the pomegranates' peels and seeds and have different curative effects. 30 Like pomegranate juice, the peels of pomegranate include ellagic acid, ellagic tannin, and gallic acid. 30 Many plant phenols such as ellagic, chlorogenic, caffeic, and ferulic acids and catechins are seen as crucial antimutagenic and anticarcinogenic agents. 12,32 Ricci et al. 33 have reported that the polyphenol content of the pomegranate peel (1.892±0.1070 mg/g dry weight) is much higher than that of fruit juice (0.063±0.0003 mg/g dry weight) and its seed (0.051±0.0075 mg/g dry weight). In our study, we determined the radical scavenging activity and the phenolic and anthocyanin contents of the methanol extract, which we prepared from the peels of pomegranates obtained from the Mediterranean region. There are studies reporting the antiproliferative and apoptotic effects of the pomegranate extracts on cancer cells. 7,34,35 The apoptotic effect possesses a great importance in inhibiting cancer formation or eradicating the cancer in terms of a successful treatment. Apoptosis occurs by means of mechanisms in which many molecules take charge and interact with each other. Bcl-2 family members are responsible for delivery of the apoptotic stimulus to the mitochondrial level, and Bax, Bid, Bcl-2, and Bcl-xL activations affect the cascade of caspases. 3
In this study, a methanol extract of peels showed an antiproliferative effect on MCF-7 breast cancer cells depending on concentration and incubation interval. In the TUNEL assay, it was also shown immunohistochemically that the apoptotic effect occurred particularly after 48 and 72 hours and with 100, 200 and 300 μg/mL PPE. PPE was as effective on Bcl-2 and Bax mRNA expressions by using the RT-PCR method. Bcl-2 and Bcl-xL are proteins assigned in the mitochondrial pathway of apoptosis. Bcl-2 is an effector molecule suppressing the apoptosis. Bcl-2 forms a heterodimer complex with Bax, a pro-apoptotoic molecule. The Bax/Bcl-2 ratio is of great importance in determining death and life. 26,36
In our study, we found that the Bax/Bcl-2 mRNA expression level, one of the most significant signs in apoptosis, increases densitometrically with 200 and 300 μg/mL PPE concentrations after 48 and 72 hours. This result shows parallelism with results of the apoptotic effect found via our TUNEL assay. DNA is broken into molecule-based fragments in early apoptosis, and the fragments of DNA are defined with the TUNEL assay. Besides, the formation of the internucleosomal DNA fragments may change according to the characteristic features of the cells. Therefore, using a couple of methods affects the investigation of the apoptotic effect beneficially. 37 The reduction in Bcl-2 mRNA expression and increase in Bax mRNA expression explain the fact that the mitochondrial apoptotoic pathway is influential.
Khan et al., 38 consistent with our results, found in one of their studies that 50, 100, and 150 μg/mL concentrations of the pomegranate extract having high antioxidant activity decrease A549 cancer cell viability at rates of 33%, 44%, and 47%, respectively, and they have demonstrated the inhibitory effect depending on the pomegranate extract concentrations and the increase in the cells at G1 phase. In another study performed with pomegranate extracts (seed oil, fermented pomegranate juice, and pericarp polyphenols), it has been reported that extracts exhibit inhibitory properties on the proliferation of human prostate cancer cells in vitro and antitumor activities. 28 Using the MTT assay, it has been indicated that the increase in MDA-MB-231 human cancer cells is decreased at the end of the 48- and 72-hour incubations with 0.5%, 1.0%, and 5.0% concentrations of pomegranate juice depending on incubation time and concentration. 39 In an in vivo study on the investigation of anti-ulcer and antioxidant activities of 70% methanolic extract of pomegranate fruit rind, it has been found that the pomegranate rind causes a decline in lipid peroxidation levels and a rise at glutathione degree in ulcerous experimental animals. 26 Mehta and Lansky 4 have reported that pomegranate fruit extracts show antiproliferative and proapoptotic effects depending on their concentrations in MCF-7, MD-MBA-231, and MD-MBA-435 breast cancer cells; it has been noted that a 50–100 μg/mL concentration of the pomegranate extract inhibited the cell proliferation in MDA-231 breast cancer cells at a rate of 67% on day 5 as measured by MTT assay. In a concentration range of 100-300 μg/mL, the pomegranate extract has been found to cause apoptotic effects on MDA-231 and SUM149 breast cancer cells. 4 Toklu et al. 7 have stated that PPE enhances early and post-apoptotic cell numbers in rat leukocytes after irradiation as a result of flow cytometric analysis. Also, Hong et al. 27 have reported that pomegranate products and polyphenols induce apoptosis in prostate cancer cells and diminish the tumor growth. It has been demonstrated using the TUNEL assay that the pomegranate extract inhibits the cell viability and nuclear factor-κB expression in prostate cancer cells and increases the apoptosis in cancer cells. 34
Pomegranate peel has been reported to inhibit the growth of human prostate cancer cells (PC3) by regulating kinase pathways dependent on cyclin (cyclin kinase inhibitor) and to exhibit apoptotic effects. When PC3 cells were incubated with 10–100 μg/mL pomegranate peel for 48 hours, the extract inhibited the cell growth depending on concencentration using the MTT method. In agreement with our mRNA expression results, it has been documented in PC3 cells that Bcl-2 protein expression is decreased and Bax protein expression is increased at the end of a 48-hour incubation with PPE, and, consequently, the apoptotic effect is observed. 40
In conclusion, it was determined for the first time in the present study that methanolic PPE exhibits antiproliferative and apoptotoic effects on MCF-7 breast cancer cells depending on concentration and incubation time. We explained in particular the apoptotic effect after 48 and 72 hours and at a high concentration of PPE by the pathway indicated by the Bax/Bcl-2 mRNA densitometric rate increase and by the determination of the apoptotic cell number as measured by the TUNEL assay. Based on our RT-PCR results, we suggest that PPE uses the mitochondrial apoptotic mechanism on MCF-7 cells. Therefore, we are of the opinion that the study of polyphenolic substances present in PPE as single or combined effects and the investigation of their apoptosis mechanisms in MCF-7 cells will contribute to the knowledge for the development of novel chemotherapeutic and chemopreventive agents to treat breast cancer.
Footnotes
Acknowledgments
This study was supported by a generous grant of the Scientific Research Projects of Anadolu University, Eskisehir, Turkey (project number 060314). Experiments were studied in the Cell Culture Laboratory of the Medicinal Plants, Drugs and Scientific Research Center and the Molecular Pharmacology Laboratory, Faculty of Pharmacy, Anadolu University.
Author Disclosure Statement
No competing financial interests exist. All the authors report no conflict of interest.