
Abstract
Konjac glucomannan (KGM) is a dietary fiber found in Amophophallus konjac. This fiber is fermentable based on human and animal trials, but short-chain fatty acid (SCFA) production profiles are unknown. The aim of this study is to characterize the digestibility and fermentability in vitro of two preparations of KGM, to better understand how KGM improves human health. Konnyaku (yam cake made of A. konjac), isolated KGM, inulin, and guar gum were subjected to in vitro digestion and in vitro fermentation. Fermentation samples were removed at 0, 4, 8, 12, and 24 hours for gas volume, pH, and SCFA measurements. Acetate, propionate, and butyrate were measured with gas chromatography. Results of the in vitro digestion confirm that KGM and konnyaku are resistant to degradation by digestive enzymes. Gas production in fermentation vessels containing konnyaku and KGM was lower than for inulin from 8 to 24 hours. Both samples produced SCFA concentrations similar to guar gum, which favored acetate and propionate over butyrate production. This study is the first to characterize SCFA production by KGM in its isolated form and in food form. Fermentation patterns presented in this study may provide a mechanism for the previously published health benefit of konnyaku and KGM.
Introduction
K
KGM is a fermentable fiber to which has been attributed many beneficial health effects such as relieving constipation, 2 –4 increasing counts of beneficial colonic bacteria (prebiotic effect), 5 reducing serum lipids, 6,7 attenuating blood glucose, 6 –8 and contributing to weight loss. 9 Physicochemical properties such as viscosity, water-binding, and bacterial short-chain fatty acid (SCFA) production may be mechanisms behind such beneficial effects. The gel-like nature of viscous fibers delays gastric emptying time and slows carbohydrate absorption in the small intestines. Water-binding capacity of some types of dietary fiber, such as psyllium, increases fecal bulk and aids in stool passage.
SCFAs—namely, acetate, propionate, and butyrate—are the primary end products of bacterial carbohydrate fermentation. Fibers that are rapidly fermented, such as inulin and fructooligosaccharides, result in high SCFA concentrations in the proximal colon and low SCFA concentrations in the distal colon. More slowly fermentable fibers such as partially hydrolyzed guar gum and some resistant starches (dextrins) may allow SCFAs to be produced in more distal areas of the colon. 10 This is of particular interest because most diseases of the colon, such as colorectal cancer and ulcerative colitis, occur in the distal colon. 11,12 The fermentation of KGM in the colon may also influence inflammatory factors, bacterial enzyme activity, and cytotoxicity. 5,13 –15 Current research has shown that fecal water is less toxic in both humans and mice when KGM is supplemented in the diet compared with no supplementation. 5,15 This is associated with greater fecal SCFA concentrations for the KGM-fed mice than the control-fed mice.
Although it is known that KGM is a fermentable carbohydrate, the profile of SCFA production has not been investigated. The aim of this study was to determine the digestibility of and fermentability of isolated KGM and KGM in food form (konnyaku [yam cake made from A. konjac]) compared with the known fermentable dietary fibers inulin and guar gum.
Materials and Methods
Four dietary fiber samples were subjected to in vitro digestion and fermentation: konnyaku (KGM from konnyaku, JFC International, San Francisco, CA, USA), glucomannan (isolated KGM, Nature's Way, Springville, UT, USA), inulin (Fiber-sure, Procter and Gamble, Cincinnati, OH, USA), and guar gum (Bob's Red Mill Natural Food, Milwaukie, OR, USA). A negative control sample (blank) was tested to assess the impact of residual digestive enzymes on the fermentation process. A positive control (glucose) was included in the fermentation to confirm the viability of the bacterial slurry. Konnyaku was freeze-dried to remove water content (760 torr at −45°C for 30 hours) (Virtis 50L, SP Industries, Warminster, PA, USA). Human salivary α-amylase (EC 3.2.1.1), porcine pepsin (EC 3.4.23.1), porcine pancreatin (EG/EC 232-369-0), bovine bile (EG/EC 232-369-0), and all chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA).
The digestion procedure was conducted according to the procedure of Amrein et al., 16 with one digestion vessel prepared for each treatment. In brief, 10 g of each treatment was suspended in phosphate-buffered saline (500 mL, 20 mM, containing 10 mM NaCl, pH 6.9) at 37°C. The entire experiment took place under continuous agitation in a water bath at 37°C. Human salivary α-amylase (0.25 mL, 20 mg/mL, in 1 mM CaCl2) was added to the mixture and incubated for 15 minutes. The pH was adjusted to 2.0 with HCl followed by addition of porcine pepsin (1.25 mL, 1 mg/mL, containing 9 g/L NaCl) and incubated for 30 minutes. The pH was increased to 6.9 with NaOH followed by addition of porcine pancreatin in phosphate-buffered saline (5 mL, 102 mg/mL) and 3.4 g of bovine bile. The mixture was incubated for 3 hours. Each treatment was placed in dialysis tubing with pore size of 3,500 Da molecular size cutoff (Spectrum Laboratories Inc., Rancho Dominguez, CA, USA) and subjected to continuous movement of distilled water for 24 hours. Digestion residues were removed from the tubing and freeze-dried (760 torr at −45°C for 30 hours) (Virtis 50L). Percentage recovery was calculated based on dry weight of digestion residue and starting weight of sample and enzymes.
Digestion residues and nondigested samples were fermented using an in vitro batch method. 17 One-half gram of nondigested treatment was placed in a sterile 100-mL serum bottle and sealed. Each treatment at all time points was examined in triplicate. Treatments were hydrated for 12 hours in 40 mL of sterile trypticase peptone fermentation medium at 4°C before the start of the fermentation. One liter of trypticase peptone medium contained 2 g of trypticase peptone, 0.8 g of ammonium bicarbonate, 2 g of anhydrous sodium phosphate dibasic, 1.25 g of anhydrous potassium phosphate monobasic, 0.5 g of magnesium sulfate, 100 mg of calcium chloride, 63.5 mg of manganous chloride, 15.5 mg of cobalt chloride, 5.2 mg of ferric chloride, and 0.01 mg of resazurin. Bottles were warmed to 37°C in a shaking water bath 2 hours prior to inoculation.
Feces were collected under anaerobic conditions from three healthy donors consuming an unspecified Western diet who had not taken antibiotics for 3 months prior to collection and who had no history of bowel conditions. Feces were homogenized and combined with phospahte-buffered saline in the ratio of 1:6. Two parts reducing solution (950 mL of distilled water, 6.25 g of cysteine hydrochloride, 40 mL of 1 N NaOH, and 6.25 g of sodium sulfide) were combined with 15 parts fecal slurry. 18 To initiate fermentation, 10 mL of the fecal inoculum was added to each bottle along with 0.8 mL of Oxyrase® (Oxyrase Inc., Mansfield, OH, USA) to remove oxygen. Bottles were flushed with carbon dioxide, sealed with rubber stoppers and aluminum seals, and placed in a 37°C shaking water bath. Bottles were removed at 0, 4, 8, 12, and 24 hours. Gas was extracted using a 40-mL syringe, and volume was recorded. The pH was determined using an Orion model 350 PerpHect Log R meter (Orion Research, Inc., Beverly, MA, USA). Copper sulfate (1 mL, 200 g/L) was added to each bottle to cease fermentation. A 2-mL aliquot was taken from each serum bottle and frozen at −20°C until SCFA extraction was completed.
Two milliliters was taken from completed fermented sample bottles for SCFA analysis. SCFAs of samples at each time points were extracted by centrifugation (3,000 g for 10 minutes) and filtering with a 1-mL syringe and a nylon filter (pore size, 0.45 μm). SCFAs were analyzed by gas chromatography. Ethyl butyrate was added as an internal standard. SCFA concentrations were determined on a HP 5890 series gas chromatograph (Hewlett Packard, Palo Alto, CA, USA) with a flame ionization detector using a Stabilwax–DA (30 m×0.53 mm i.d.; film thickness, 1 μm) fused silica column (Restek, Bellefonte, PA, USA). Sample injection volume was 0.5 μL. Inlet and detector temperatures were 200°C. The oven temperature was held at 90°C for 2 minutes, increased to 120°C at a rate of 60°C/minute, and held for another 10 minutes. Helium served as the carrier gas, and injections were made in split mode (1:100) with a flow rate of 31.5 mL/minute. Hydrogen and air flow rates were 30.0 mL/minute and 280 mL/minute, respectively. Peaks were identified by comparing their retention times with standard compounds. Integration of chromatographic peak areas was completed using HP Chem Station (Hewlett-Packard). SCFA values were quantified by using equations derived from a standard curve through the use of peak areas. Total SCFA concentration was determined from the total peak area and the ethyl butyrate peak area. The experiment was a randomized complete block. One block consisting of pooled fecal samples from three humans was used. The treatments were arranged in a 6×5 factorial pattern with six treatments measured at five time points. Each fiber×time point combination was measured in triplicate. SAS statistical software version 9.1.3 (SAS Institute, Cary, NC, USA) was used to conduct analysis of variance, with Tukey's pairwise comparison conducted to compare mean gas volume, pH, and SCFA concentrations. Statistical significance was achieved at P values of <.05.
Results
Percentage recovery after digestion was greatest for guar gum (98.8%). KGM and konnyaku were also well recovered after digestion (97.9% and 88.3% recovery, respectively). Slightly more than half of the inulin was recovered after digestion (57.6%).
Bacteria used in the in vitro fermentation were viable as shown by the low pH and high gas production in the glucose sample (Figs. 1 and 2). The pH values dropped sharply from 0 to 4 hours and were maintained relatively stable from 4 to 24 hours (Fig. 1). Konnyaku had consistently higher pH values than all other fibers from 8 to 24 hours. Guar gum and KGM had similar pH values at 4, 12, and 24 hours. Inulin consistently produced the lowest pH value of any fiber (excluding the glucose control).
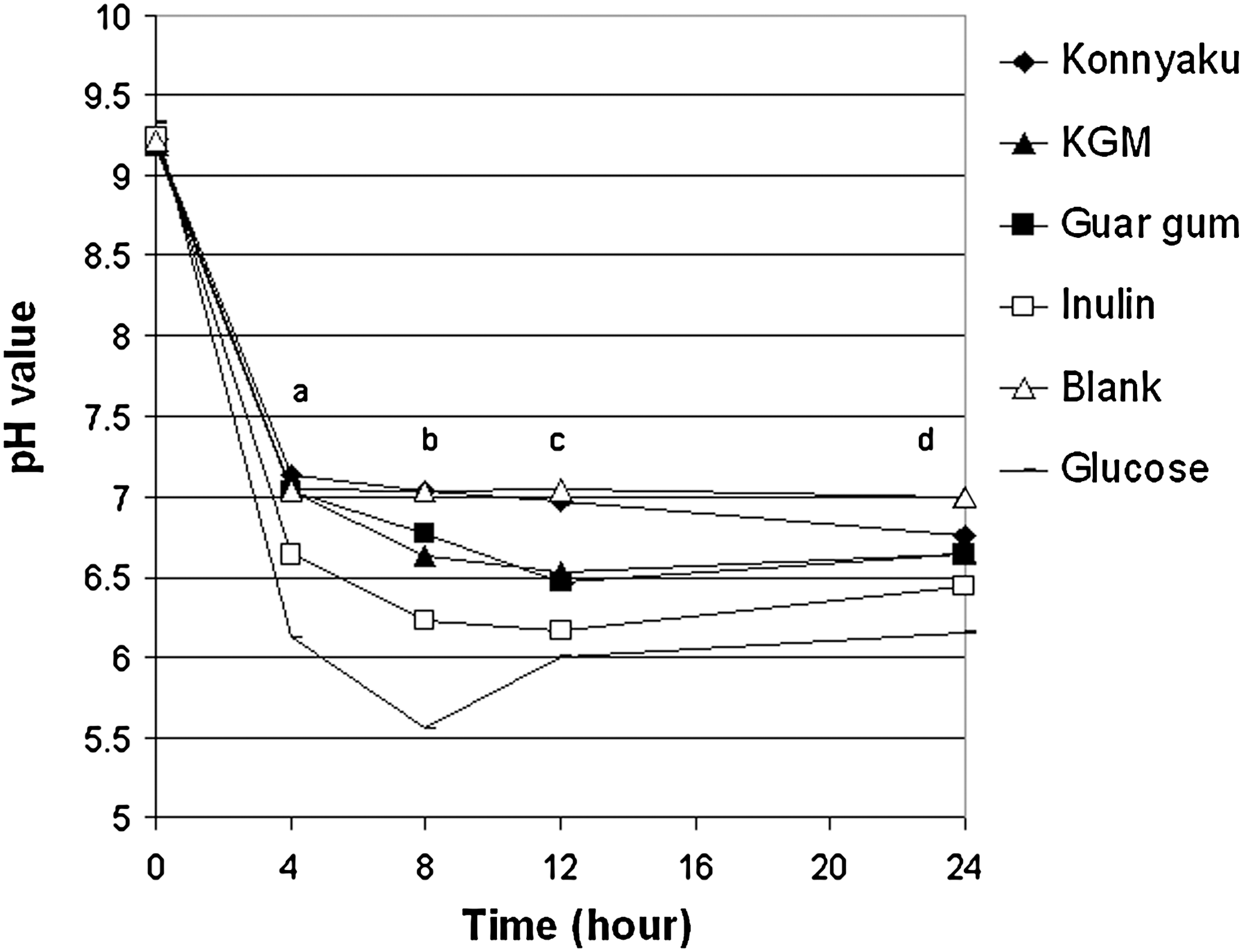
The pH of fermentation slurry over a 24-hour period of in vitro fermentation. Data are mean values (n=3). a P<.001, konnyaku=blank=konjac glucomannan (KGM)=guar gum>inulin>glucose; b P>.0001, konnyaku=blank>guar gum>KGM>inulin>glucose; c P<.001, blank=konnyaku>KGM=guar gum>inulin>glucose; d P<.001, blank>konnyaku>KGM=guar gum>inulin>glucose.
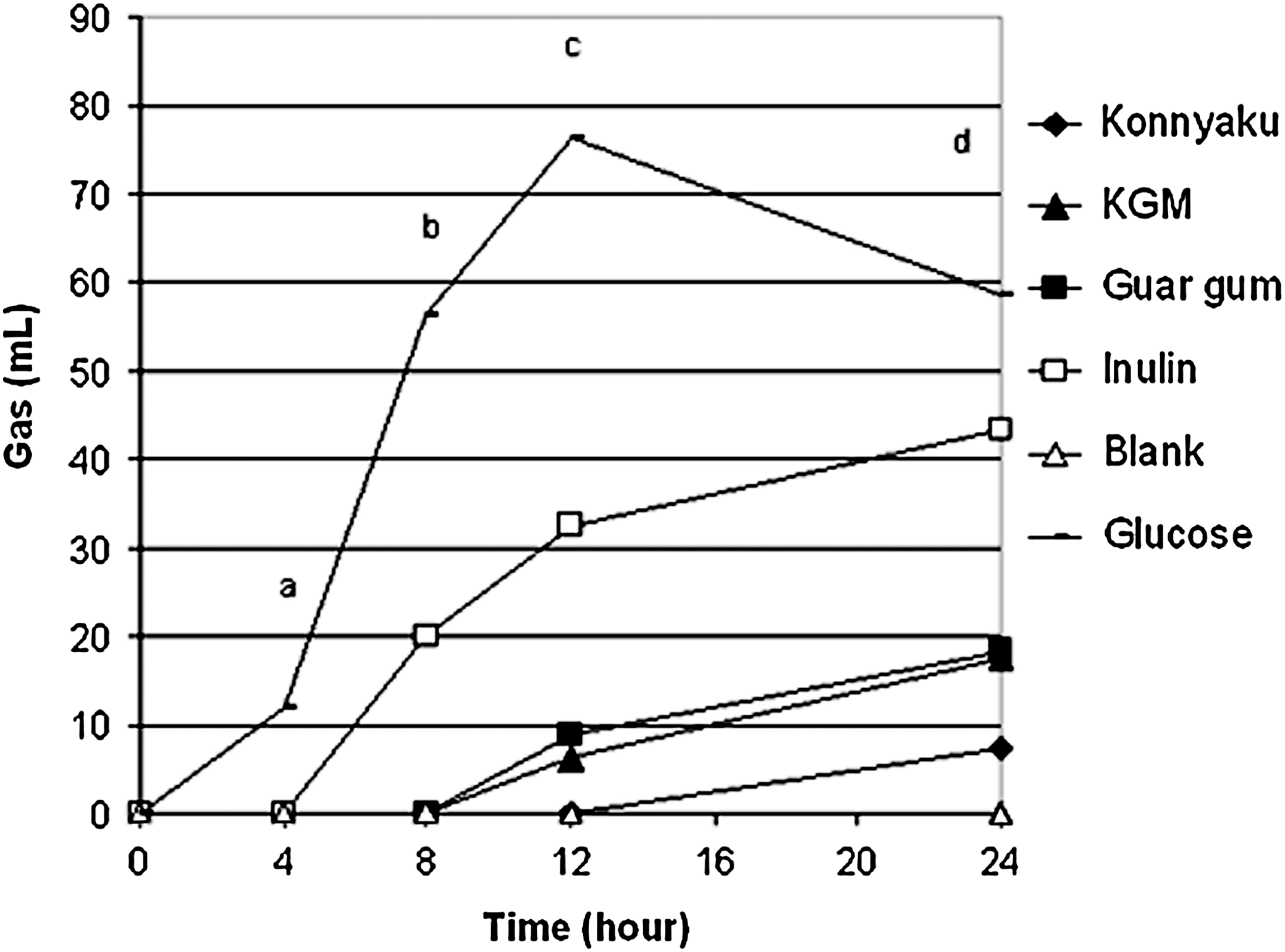
Gas volume of fermentation slurry over a 24-hour period of in vitro fermentation. Data are mean values (n=3). a P<.001, glucose>all others; b P<.001, glucose>inulin>all others; c P<.0001, glucose>inulin>guar gum=KGM=konnyaku=blank; d P>.0001, glucose>inulin>KGM=guar gum>konnyaku=blank.
Inulin produced the highest gas volumes from 8 to 24 hours (excluding the glucose control [Fig. 2]). Gas production for konnyaku was measurable only at the 24-hour time point. Guar gum and KGM produced similar gas volumes during the entire fermentation period.
Acetate production differed the most among samples at 4 hours (Fig. 3A). Inulin resulted in the greatest acetate production, whereas konnyaku resulted in the lowest acetate production. At 8 hours, inulin continued to support high acetate production, whereas guar gum, KGM, and konnyaku yielded lower concentrations of acetate. By 12 hours, bacterial production of acetate was similar among all fibers, excluding glucose. At 24 hours, glucose resulted in greater acetate than both guar gum and konnyaku; however, fiber treatments (guar gum, konnyaku, KGM, and inulin) did not differ. Bacterial propionate production was low in response to all fibers from 4 to 12 hours (Fig. 3B). By 24 hours, inulin resulted in significantly higher propionate production than KGM. Butyrate production was not different among the samples (excluding the glucose control [Fig. 3C]). Bacterial SCFA production in response to four different fibers showed that samples favored acetate and propionate production more than butyrate production (Fig. 4). Total SCFA production did not differ among the four fiber samples.
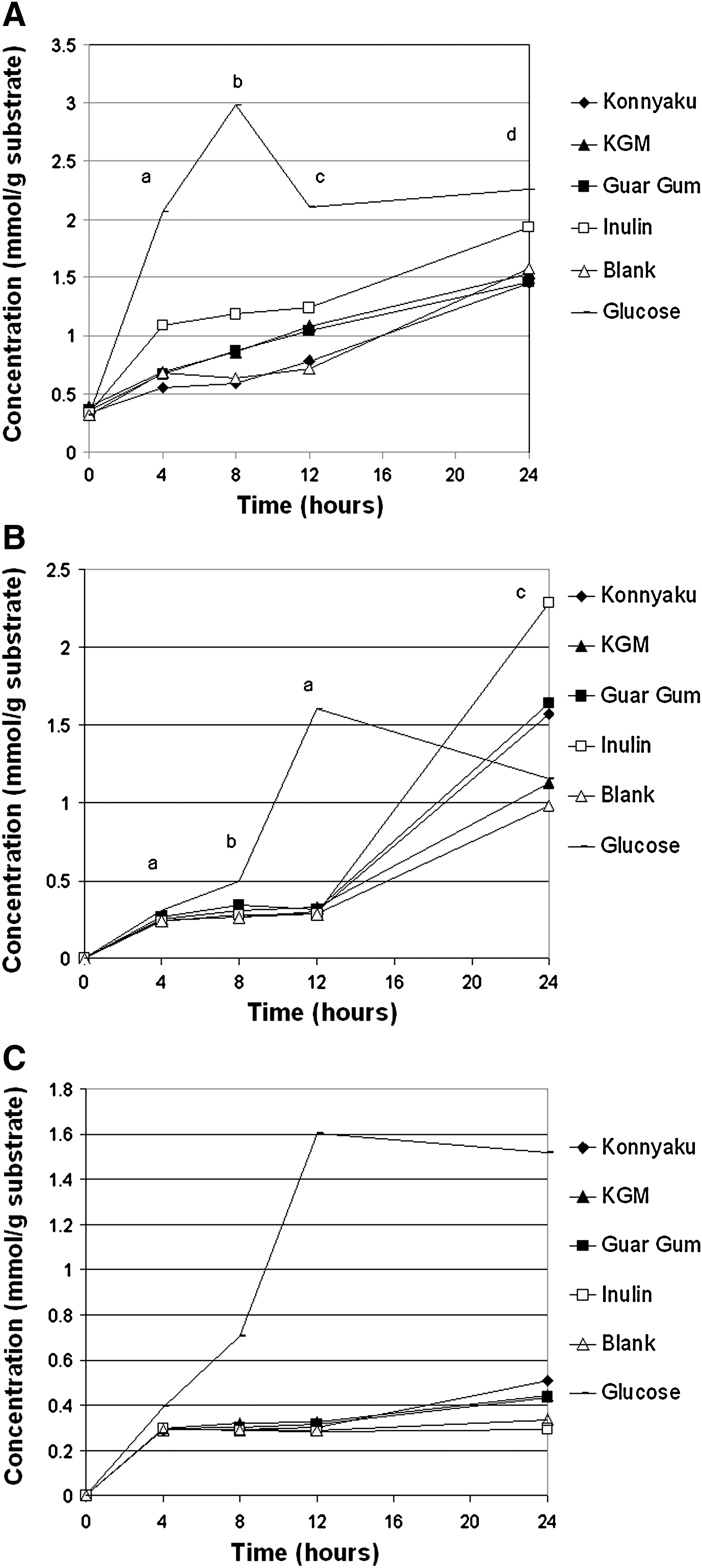
Bacterial short-chain fatty acid production over a 24-hour period of in vitro fermentation: (
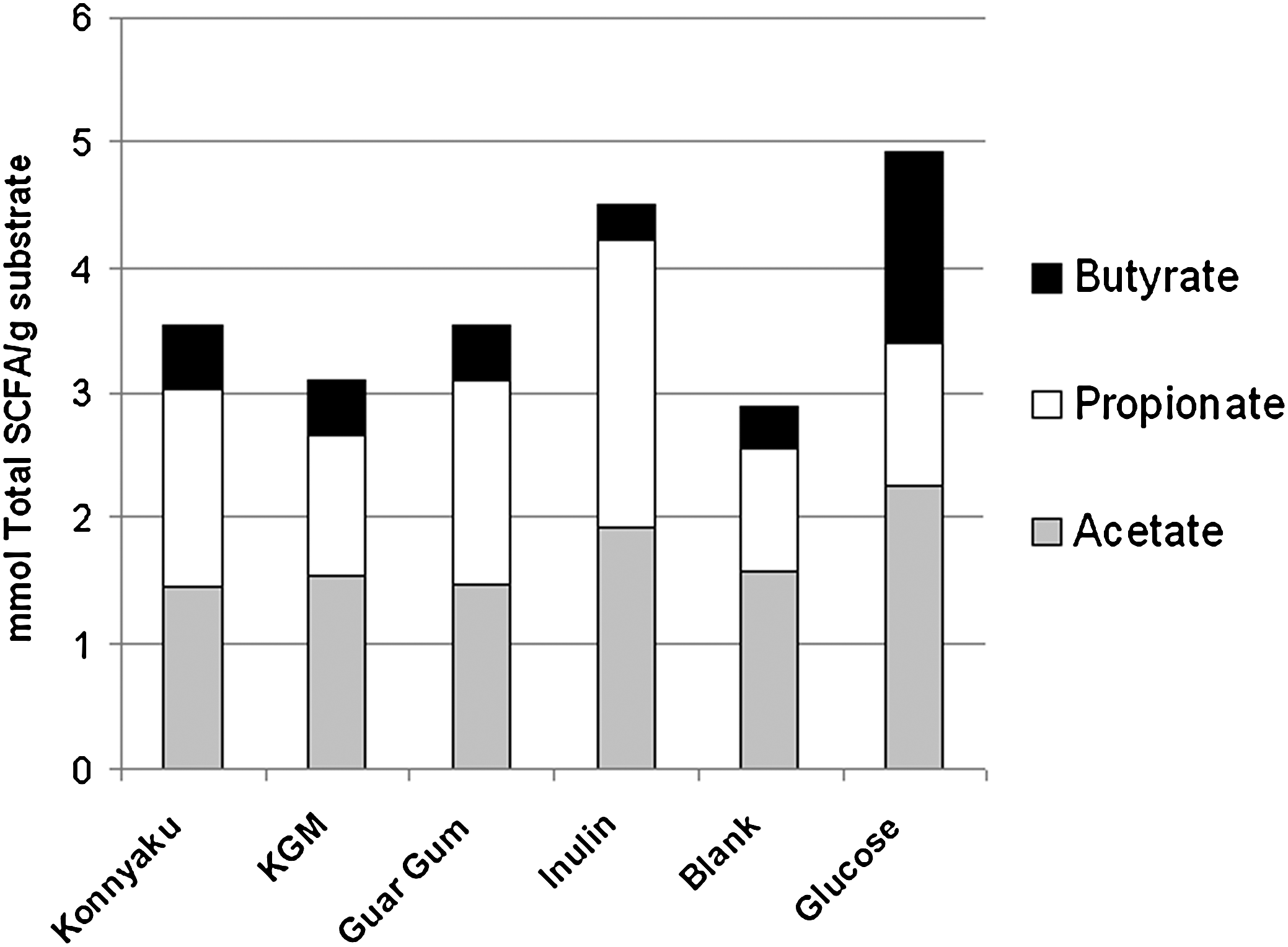
Short-chain fatty acid (SCFA) concentrations at 24 hours of in vitro fermentation. Data are mean values (n=3). For total SCFA production at 24 hours: P=.0021, glucose > KGM, blank. All other comparisons not significant.
Discussion
This study is the first to report on digestibility and bacterial SCFA production in response to konnyaku (yam cake made from A. konjac) and KGM (isolated glucomannan derived from A. konjac) in vitro. Konnyaku, KGM, and guar gum resisted digestion with high recovery rates, confirming that the majority of each substance should reach the colon intact. Percentage recovery of inulin was lower than expected (57.6%) and may be a result of the dialysis tubing used to filter digested material from polymers. Inulin is an oligofructan with 20 units as an average chain length. Potentially, shorter oligofructans may have escaped the dialysis tubing, resulting in a lower recovery rate. Future in vitro digestions should use dialysis tubing with a smaller pore size (<3,500 Da molecular size cutoff) to reduce the risk of losing low-molecular-weight compounds at the end of the digestion.
Changes in pH, gas production, and production of SCFA confirmed that konnyaku and KGM are fermentable carbohydrates, with steady fermentation profiles similar to that of guar gum. In vivo, it is estimated that SCFA production will extend beyond the proximal colon and may reach the distal colon, thus exerting a protective effect. However, this is dependent on the presence of saccharolytic bacteria and should be confirmed in vivo. The molecular weight of KGM influences fermentation rates. Using a mouse model (BALB/c), Chen et al. 19 reported that hydrolyzed KGM was more fermentable than conventional KGM. Hydrolyzed KGM and KGM both resulted in greater fecal acetate, propionate, and n-butyrate concentrations than control-fed mice after 4 weeks of supplementation. Fermentation of KGM was documented in humans based on decreased fecal pH and increased fecal total SCFA, acetate, propionate, and n-butyrate concentrations compared with control diet. 4 More recently, BALB/c mice supplemented with KGM or inulin presented higher fecal acetate concentrations than control mice. 15 Fecal butryate and propionate concentrations were lower after KGM supplementation than for inulin. High cecal acetate concentrations were reported in Sprague–Dawley rats consuming KGM or inulin supplemented chow. 14 Individual and total SCFA concentrations did not differ between KGM- and inulin-supplemented groups.
Data from this study at 24 hours of in vitro fermentation confirm the propionate findings by Chen et al. 15 but are in disagreement with the butyrate findings. Our study reports that butyrate production between KGM and inulin was not different at 24 hours. These differences may be due to colonic absorption rates in the mouse study and/or difference in colonic bacteria and metabolic capacity. Total SCFA concentrations at 24 hours of in vitro fermentation confirm the results found in mouse, rat, and human studies: KGM and inulin produced similar concentrations of total SCFAs.
This study builds the current body of knowledge by quantifying SCFA production over the average 24-hour colonic retention time. 20,21 Low gas production by konnyaku and KGM during the first 12 hours suggests that these fiber sources should cause minimal flatulence and bloating in humans. Chen et al. 4 confirmed our findings in humans. Humans consuming 4.5 g of KGM/day for 3 weeks reported minimal gastrointestinal side effects (abdominal cramping, borborygmi, bloating, and flatulence) compared with 0 g of KGM/day.
Fermentation demonstrated in this study may provide a mechanism by which KGM exerts a laxative effect. KGM has been shown to increase laxation in adults and children as measured by the number of defecations per day. 2 –4 The dose effective for adults was 4.5 g of KGM/day. 2,4 Children consumed 100 mg of KGM/kg of body weight and did not exceed 5 g of KGM/day. 3 A strong placebo effect was seen in childhood constipation, with significantly softer stools for both treatment and control groups. KGM increased total intestinal motility based on mouth to cecum transit time. 22 KGM at 3 g/day for 10 days normalized mouth to cecum transit times for constipated subjects (not different than healthy control transit times). However, conflicting reports have indicated that KGM delays gastric emptying. 22 Changes in fecal wet weight were inconsistent in adults, so laxative properties cannot be clearly linked to increased fecal bulk. 2,4 In both adult studies, the sample size was small (seven or eight subjects, respectively). Fecal wet weight was not assessed in children. Laxative properties may be linked to KGM's prebiotic effect. Two studies in mice have demonstrated that KGM possesses a prebiotic effect by increasing counts of bifidobacteria after 2–4 weeks of supplementation. 19,23 In humans, KGM exhibited a prebiotic effect after consuming 4.5 g/day for 3 weeks. 4 Lactobacillus counts increased with treatment. Other prebiotics, such as inulin, lactulose, and galactooligosaccharides, have produced laxative effect in some studies, but results are inconclusive. 24
In conclusion, konnyaku and KGM are resistant to digestion and are steadily degraded by colonic bacteria over a 24-hour period with minimal gas production in vitro. These findings support previous studies conducted in mice, rats, and humans. Together, these data link beneficial physiological effects attributed to KGM with bacterial SCFA production. Future work should be conducted to identify if the source of KGM (isolated or as a part of konnyaku) produces similar physiological effects in humans.
Footnotes
Acknowledgment
This research was supported by the College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa, Honolulu, HI.
Author Disclosure Statement
The authors declare no conflict of interest.