
Abstract
Several biological and clinical studies have suggested that conjugated linoleic acid (CLA) prevents body fat accumulation and increases lean body mass. CLA is available as a concentrated dietary supplement and is purported to provide the aforementioned benefits for people who perform physical activity. This study was conducted to evaluate the effect of a CLA-supplemented diet combined with physical activity on the body composition of Wistar rats. Two groups of Wistar rats of both sexes, between 45 and 60 days old, were fed a diet containing 5.5% soybean oil (control group) or a CLA-supplemented diet (0.5% CLA and 5.0% soybean oil) (test group). Half the rats in both groups were assigned to exercise by running on a treadmill. The biochemical and anatomical body compositions were analyzed. In both groups, CLA had no effect on the dietary consumption or the weight of the liver, heart, and lungs. However, it did influence the overall weight gain of exercised male rats and the chemical and anatomical body composition in exercised and sedentary rats of both sexes. The results confirm that a CLA-supplemented diet with and without physical activity reduced body fat accumulation in rats of both sexes. However, there is no evidence of an increase in the lean body mass of the exercised rats.
Introduction
T
The main mechanism of action of CLA is the reduction in lipogenesis, which has been the subject of extensive research. 5 –8 The antilipogenic activity of CLA was initially verified through an experiment that demonstrated a significant reduction in lipogenesis due to the inhibition of lipoprotein lipase located on the surface of adipocytes, which would in fact limit the incorporation of fatty acids into the cell. 5 An increase in the carnitine palmitoyltransferase activity in the mitochondria of muscle cells was also verified. This enzyme transports fatty acids involved in the Krebs cycle and provides precursors for the synthesis of amino acids, which are essential for muscle tissue formation.
Preliminary results indicated that CLA affects body composition by increasing lean mass and reducing adipose tissue in different animal species, 9 –12 as well as in humans (but to a lesser extent). 6,13,14 The results from these previous studies have been controversial, probably because the the objective of the human studies was the reduction of existing fat deposits, whereas in animals, the aim was to decrease the number of adipocytes and consequently reduce fat deposition.
One concern is that although there is not enough scientific evidence to support the promising effects of CLA in converting adipose tissue into muscle mass, people often use CLA products indiscriminately, completely unaware of the effectiveness of the doses ingested. Therefore, the present study aims at evaluating the effect of a CLA-supplemented diet combined with physical activity on the body composition according to sex of Wistar rats.
Materials and Methods
Animals, diets, and exercises
Sixty-four adult Wistar rats (32 males and 32 females) between 45 to 60 days old and with an average weight of 265 g (males) and 232 g (females) were used in this study. The animals were provided by the Laboratory of Human Nutrition Vivarium, Department of Agroindustry, Food, and Nutrition, Luiz de Queiroz College of Agriculture of the University of São Paulo, São Paulo, Brazil. Each rat was placed in an individual cage at a temperature of 23±1°C and under a 12-hour light/12-hour dark cycle. They were subjected to a period of adaptation to physical exercises of 15 days and to diets with commercial CLA (a 50:50 ratio of the CLA isomers cis-9, trans-11 and trans-10, cis-12) of 1 week.
The animals were divided according to sex. The first group consisted of 16 rats that were subjected to physical activity (exercised rats) and included a group fed a CLA diet (n=8) and another fed a control diet (n=8). The second group consisted of 16 sedentary rats including a group fed a CLA diet (n=8) and another fed a control diet (n=8). The exercised rats were subjected to daily activity in a running wheel for 40 minutes at a speed ranging from 10 to 15 m/minute.
All animals were allowed daily access to water and 25 g of the corresponding diet. The experimental period lasted 65 days. Daily feed consumption and weight of the animals was recorded.
Table 1 shows the composition of the CLA-supplemented diet and control diet according to the animals' nutritional requirements. 15 The 0.5% CLA amount used in the diet was determined based on a review of previous successful studies. 16
Part of the standard AIN-93G diet. 15
Commercial dietary supplement conjugated linoleic acid (CLA).
This study was approved by the Ethics Committee for Research on Animals, Luiz de Queiroz College of Agriculture, following protocol number 2008-9. All experimental procedures were followed according to the national and institutional guidelines for the care and use of animals.
Body composition analysis
At the conclusion of the experiment, the animals were sedated with halothane following a 12-hour fast. Immediately after sacrifice, the organs and body fat (abdominal, subcutaneous, visceral, and retroperitoneal) were removed, separated, and weighed. The retroperitoneal and subcutaneous adipose tissues were added to the carcass weight. The biochemical composition of the carcass without the organs was calculated from a sum of the values obtained for carcass and adipose tissues. The carcasses were labeled, frozen, and stored at −20°C until ready for analysis. All biochemical analyses were performed according to the official analysis methods of AOAC International. 17 The water content of the carcass was calculated by subtracting the weight of the carcass after desiccation at 105°C for 12 hours from the initial carcass weight. The total nitrogen content was determined by the Kjeldahl method and multiplied by 6.25 to determine protein content. The lipid contents were determined by extraction with ethyl alcohol and using a Soxhlet apparatus. The minerals were quantified from the total ashes after incineration at 550°C for 6 hours.
Statistical analysis
Results are presented as mean±SD values. Analysis of variance was used to determine the significance of differences in body weight gain, organ weight, and adipose tissue, as well as the biochemical composition of the carcasses in the treatments. The data were analyzed based on a 2×2×2 factorial experimental design. Factors in the model were diet, physical activity, and sex and their interactions. When the effect on the diets was statistically significant (P<.05), the differences were determined using Tukey's test for comparison of the means. All analyses were carried out using SAS statistical software (version 9.0, SAS Institute Inc., Cary, NC, USA).
Results
Body weight, food intake, and organ weight
All the rats gained weight during the study. Male and female rats fed a CLA-supplemented diet exhibited body weights similar to the control group, except for exercised male rats (Figs. 1 and 2). Weight gain was significantly reduced at the end of the trial in exercised male rats fed a CLA-supplemented diet relative to their respective control group (Fig. 3). Male rats showed higher weight gain than female rats as shown in Figure 3.

Body weight of ) and CLA-supplemented diet ( ). The body weight values were analyzed using Tukey's test (*P<.05). Data are mean±SD values (n=8).
). The body weight values were analyzed using Tukey's test (*P<.05). Data are mean±SD values (n=8).

Body weight of ) and CLA-supplemented diet ( ). The body weight values were analyzed using Tukey's test (*P<.05). Data are mean±SD values (n=8).
). The body weight values were analyzed using Tukey's test (*P<.05). Data are mean±SD values (n=8).
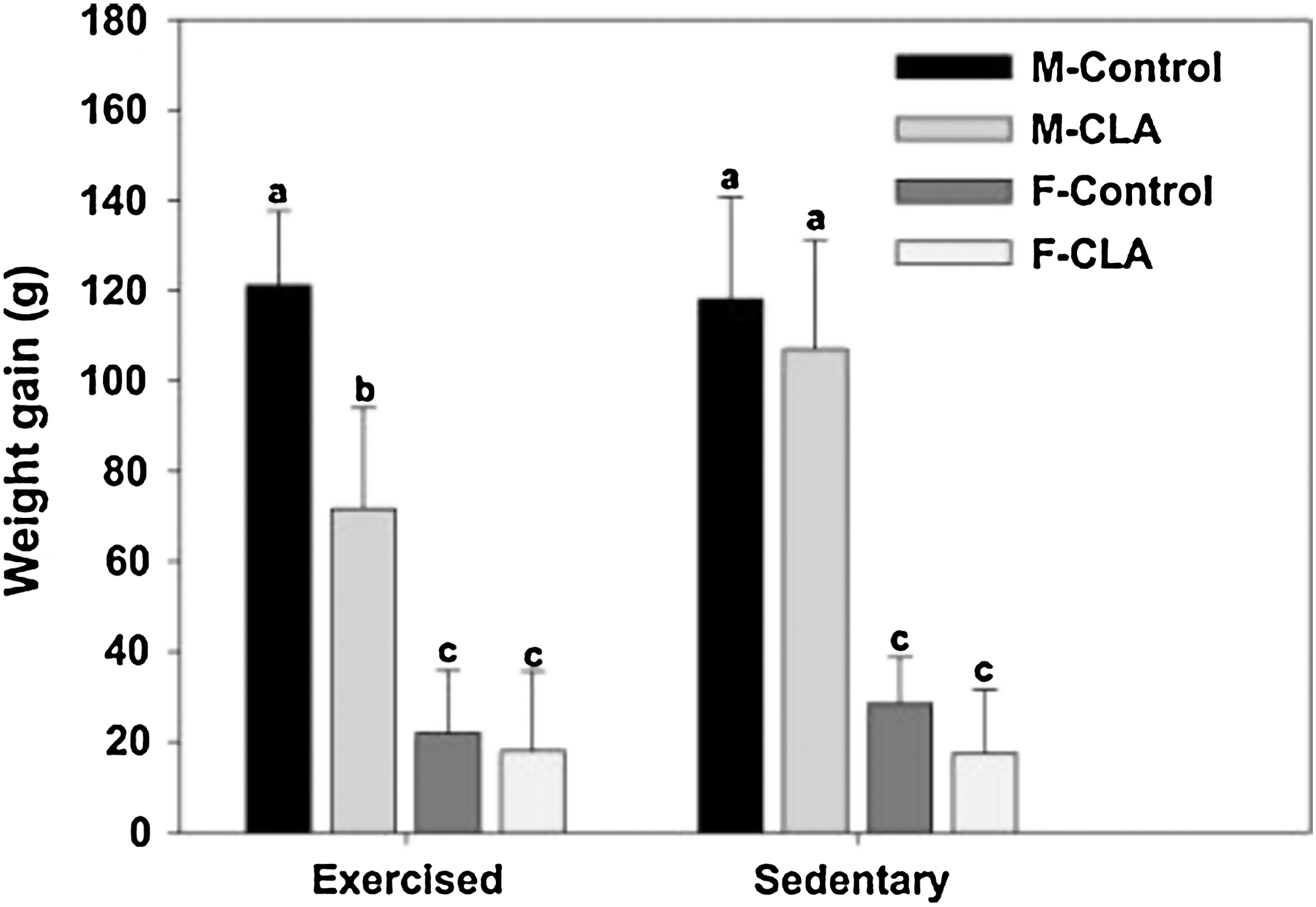
Weight gain at the end of the trial by male (M) and female (F) rats, exercised and sedentary, fed control diet and CLA-supplemented diet.
The CLA-supplemented diet did not have a significant effect on the organ weight of rats of both sexes compared with the control diet (Table 2). Physical activity was associated with an increase in the liver weight of exercised male rats fed a control diet (11.8 g) compared with those fed a CLA-supplemented diet and with sedentary rats fed a control diet (10.8 g). This same trend was not observed in the sedentary male and female rats fed either of the two diets. Sedentary male rats had an increase in lung weight compared with exercised rats. Organ weight was significantly different between the sexes (P<.0001). There was a significant interaction between diet and physical activity for liver weight, between physical activity and sex for lung weight, and between diet and sex for heart weight.
Data are mean±SD values (n=8 per treatment group).
Means in a row without a common letter are significantly different, P<.05.
Factors are diet (D), A (physical activity), and sex (S).
Expressed as (body weight gain [in g]/food intake [in g])×100.
NS, not significant.
Overall diet consumption was similar between the experimental groups, with the exception of greater consumption in male rats than in female rats. The significant difference of feed efficiency between diets is attributed to the reduced weight gain of the exercised male rats fed a CLA-supplemented diet compared with those fed a control diet.
Body composition
With regard to the carcasses, the exercised male rats presented a significant fat mass reduction when fed a CLA-supplemented diet (7.7%) compared with the control group (9.6%). Nevertheless, this reduction was not necessarily associated with a significant increase in the lean body mass (protein fraction) in these groups (Table 3). In contrast, in the group of sedentary male rats, the CLA-supplemented diet promoted a significant increase in protein content (29.8%) compared with the group of sedentary male rats fed control diet (26.6%), although it was not associated with decreased fat mass.
Data are mean±SD values (n=8 per treatment group).
Means in a row without a common letter are significantly different, P<.05.
Factors are diet (D), A (physical activity), and sex (S).
ECW, empty carcass weight; NS, not significant.
Female rats fed a CLA-supplemented diet had lower fat content compared with their control group. Nonetheless, this decrease in fat content was not associated with an increase in protein content in the subgroup of exercised rats and had the opposite result in the sedentary group. This result was similar to that in the sedentary male rats, in which there was a significant increase in the protein content of rats fed a CLA-supplemented diet (24.2%) compared with the control group (21.4%). Overall, there was significant interaction (P<.0001) between diet and physical activity for variables of body fat and protein in the animals.
Adipose tissue weight
Adipose tissue weight was greatly affected by a CLA-supplemented diet and physical activity (Table 4). Exercise decreased adipose tissue mass in both sexes. When exercise was associated with the consumption of a CLA-supplemented diet, the adipose tissue weight decreased considerably compared with that of male rats consuming control diet.
Data are mean±SD values (n=8 per treatment group).
Means in a row without a common letter are significantly different, P<.05.
Factors are diet (D), A (physical activity), and sex (S).
NS, not significant.
In female rats, physical activity combined with a CLA-supplemented diet promoted a significant reduction in abdominal (9.9 g) and subcutaneous (2.5 g) fat compared with that of exercised female control rats (14.1 and 5.2 g, respectively). The weights of abdominal, visceral, subcutaneous, and retroperitoneal adipose tissue of those animals were similar to those of sedentary female rats fed a CLA-supplemented diet.
Discussion
The results shown in Figures 1 –3 indicate that animals of both sexes fed a CLA-supplemented diet did not experience significant changes in body weight and weight gain compared with control animals, except for the group of exercised male rats. Previous studies 18 have shown that CLA did not significantly change the body weight of subjects in human clinical studies, 18 although discrepancies have been observed for body weight and weight gain in different biological models with CLA treatment. 19,20 Animals and humans have different metabolic rates, which may explain, in part, the different results of the studies. On average, the groups of exercised animals in the present study exhibited the lowest body weight compared with that of their sedentary peers. This is probably due to the association of physical activity with greater post-exercise energy expenditure in order to maintain high-intensity endurance of the resting metabolic rate. During this period of high metabolism, the muscle glycogen reserves are used during approximately the first minute of physical activity; glycogen decreases with the activity, such that the main energy substrate consumed by the organism comes from the adipose tissue, resulting in a decrease in the body lipid content. 21,22
In performing toxicological analysis of the diets supplemented with CLA, at doses from 1% to 1.5%, other studies have shown no treatment-related effects on the organ weight in rats. 23,24 This is in agreement with the results obtained in the present study, which suggest that supplementation with CLA at a single dose (0.5%) in the diet does not have adverse effects on liver, heart, and lung weights compared with that of the control diet, regardless of the physical activity and sex of the animals.
The results of the biochemical and anatomic body composition analysis of the rats confirm that a single dose of CLA (0.5%) promotes a reduction in body fat accumulation in rats of both sexes and that this reduction is more intense when combined with physical activity. However, CLA promotes a significant increase in protein content only in sedentary rats of both sexes, and not in exercised rats. This finding contradicts the initial hypothesis that an increase in lean body mass would occur when CLA was combined with physical activity. These results confirm the findings of studies in animal models investigating the effect of diets with CLA at 0.25–1% 25 and 3%. 26
When the consumption of CLA is associated with physical activity, the discrepancies in the studies become very evident. In mice that were fed a diet supplemented with 0.4% CLA, compared with mice that were fed a control diet, significant increases in lean body mass were observed in exercised animals but not in sedentary ones; 10 on the other hand, the body fat accumulation in sedentary and exercised mice fed a CLA-supplemented diet was lower than that of the group fed the control diet. Conversely, a study carried out with sedentary and exercised male Wistar rats fed a CLA-supplemented diet did not reveal any effects of CLA isomers on body fat and lean body mass in lean or fat adult animals; no significant interactions between the treatments with CLA and physical activity were observed in this study. 20 The results of a clinical trial involving 23 exercised subjects supplemented with CLA for 28 days did not reveal significant changes in the body composition, bone density, immunity, and catabolic markers of the individuals. 27 Discrepancies among studies may be attributed to different factors, such as type and amount of CLA isomers consumed, length of experimental period, difference in the biological models used, body weight, energy consumption, and, potentially, a lack of consistency among methodologies used. 13,16,28
The possible biochemical mechanisms of the functions of CLA are associated with different biological reasons. In a study with mice, supplementation with CLA isomers was associated with a reduction in lipoprotein lipase activity. However, there was an increase in the rate of lipolysis of adipocytes, causing a reduction in the concentrations of triglycerides and intracellular glycerol. In this study, an increase in the fatty acid oxidation in adipocytes and muscle cells was also observed. 5 However, another study on the incorporation of CLA isomers using the same animal model and adipocyte cells attributed cell activation in the integrated stress response pathway to CLA. This function is the probable cause of the later induction of pro-inflammatory cytokines in CLA-treated adipocytes. The release of pro-inflammatory cytokines may be associated with an activation of the integrated stress response pathway in adipocytes in their early stage, causing lipid loss in mature adipocytes. 29
More recently, rats fed a CLA-supplemented diet demonstrated an increase of fatty acid synthase activity and a reduction of glycerol 3-phosphate dehydrogenase activity. Consequently, the fatty acid synthesis pathway would be depressed in fat tissue. 30 A hypothesis suggested in another recent study regarding lipid level reduction in adipocytes is likely related (directly or indirectly) with the activation of AMP-activated protein kinase due to the action of the CLA isomers. It is hypothesized that this activation increases the phosphorylation of the acetyl-coenzyme A carboxylase protein in 3T3-L1 adipocytes, reducing the concentration of triglycerides in cells of the adipose tissue. 31
CLA could also depress body fat accumulation by reducing proliferation, inhibiting differentiation, and stimulating apoptosis of preadipocytes. 32
The activity of carnitine palmitoyltransferase in the muscle was stimulated in mice and rats fed with a CLA-supplemented diet, 5,33 thus favoring the β-oxidation of fatty acids in the muscle tissue. Another study suggested that the supplementation with CLA combined with arginine increased lean mass in male Sprague–Dawley rats by depressing the turnover of muscle proteins (sarcoplasmic or myofibrillar). 10 This mechanism may explain the increase of the lean mass in sedentary rats fed a CLA-supplemented diet compared with sedentary rats fed the control diet in our study.
In conclusion, under the experimental conditions of this study, the effects of physical activity on body composition can be confirmed. The effect of consuming a commercial mixture of CLA isomers combined with physical activity on the reduction of body fat was significant in rats of both sexes. The results obtained reject the hypothesis that the consumption of CLA combined with physical exercises promotes an increase in lean body mass in individuals, but the results suggest that this may occur in sedentary individuals (although more moderately than in the exercised individuals). Further studies are necessary to evaluate biologically active CLA isomers, either singly or in combination, and to explain their mechanism of action as well as their bioavailability.
Footnotes
Acknowledgments
The authors are grateful to the Fundação de Amparo à Pesquisa do Estado de São Paulo for financial support of this study.
Author Disclosure Statement
All of the authors report no conflicts of interest.