
Abstract
Bromelain is a proteolytic enzyme extracted from the stems and the immature fruits of pineapple that was found to be antitumorigenic in different in vitro models. Bromelain has been reported to promote apoptosis, particularly in breast cancer cells, with the up-regulation of c-Jun N-terminal kinase and p38 kinase. Our study was designed to determine if bromelain could induce apoptosis in GI-101A breast cancer cells. GI-101A cells were treated with increasing concentrations of bromelain for 24 hours. The effect of bromelain for inducing cell death via activation of the apoptosis mechanism in GI-101A cells was further determined by using caspase-9 and caspase-3 assays along with the M30-Apoptosense assay to measure cytokeratin 18 (CK18) levels in the cytoplasm of the cultured cancer cells. A dose-dependent increase in the activities of caspase-9 and caspase-3 coinciding with elevation of CK18 levels was found in bromelain-treated cells compared with control cells. Furthermore, the apoptosis induction by bromelain was confirmed by DNA fragmentation analysis and 4,6′-diamino-2-phenylindole dihydrochloride fluorescence staining of the nucleus. Our results indicate an increase in apoptosis-related cell death in breast cancer cells with increasing concentrations of bromelain.
Introduction
F
The GI-101A breast cancer cell line used in this study was derived from a poorly differentiated mammary carcinoma that was also metastatic to the lungs and lymph nodes. 9,10 These cells are positive for estrogen receptors but are resistant to anti-estrogen drugs such as tamoxifen. 10 The aim of this study was to explore the effect of bromelain at different concentrations on the estrogen receptor–positive GI-101A breast cancer cell line. The extent of apoptosis was assessed by measuring the activities of caspase-9 and caspase-3, the level of caspase-cleaved cytokeratin 18 (CK18)-Asp396 neo-epitope using the M30-Apoptosense enzyme-linked immunosorbent assay (ELISA) kit, and DNA fragmentation.
Materials and Methods
Cell line and reagents
The GI-101A human breast carcinoma cells were derived from a xenograft of a patient with recurrent ductal adenocarcinoma at the Rumbaugh Goodwin Institute for Cancer Research, Plantation, FL, USA. The RPMI-1640 growth medium, amphotericin B,
Cell culture and bromelain treatment
The human breast carcinoma cell line GI-101A was grown as a complete monolayer in RPMI-1640 growth medium supplemented with 10% fetal bovine serum, 10,000 U/mL penicillin, 10,000 μg/mL streptomycin, 1% (+)-
Assay of caspase-9 and caspase-3 activities
Breast cancer cells were treated with 5, 10, and 20 μg/mL bromelain for 24 hours, and control cells were incubated without any bromelain. After the incubation interval, cells were harvested, washed, resuspended in cell lysis buffer, and kept on ice for 10 minutes. The cell lysate was centrifuged for 1,400 g for 5 minutes, and after centrifugation the supernatant was assayed for protein concentration and then diluted with the lysis buffer to adjust the protein concentration. Equal amounts of protein from each sample were added to 96-well plates and mixed with the 2× reaction buffer (100 mM HEPES [pH 7.4], 200 mM NaCl, 20 mM dithiothreitol, 2 mM EDTA, and 0.2% CHAPS) and the respective substrates acetyl-Leu-Glu-His-Asp-p-nitroaniline and acetyl-Asp-Glu-Val-Asp-p-nitroaniline at 2 mM. For measuring the specific activity of caspase-9 and caspase-3, the respective inhibitors acetyl-Leu-Glu-His-Asp-CHO and acetyl-Asp-Glu-Val-Asp-CHO were used. Release of the cleaved p-nitroanilide from the tetrapeptide substrates was measured using the 96-well microplate reader at 405 nm.
M30-Apoptosense ELISA immunoassay
During the process of apoptosis, cleavage of CK18 by caspase 3 occurs, resulting in fragments containing the CK18-Asp396 neo-epitope. This neo-epitope is recognized by the M30 antibody provided with the Apoptosense ELISA kit, allowing for the quantification of apoptotic cells. 11 For this assay approximately 1×104 cells were grown in a 96-well plate, using complete RPMI growth medium. Once the cells attached to the plate, the cells were treated for 24 hours with increasing concentrations of bromelain at 5, 10, or 20 μg/mL, and the control was incubated similarly without bromelain. At the end of the incubation period the cells were lysed, and aliquots of the lysates were transferred to the CK18 monoclonal antibody (M30)–coated microplate wells and assayed per the manufacturer's instructions. The M30 conjugate dilution buffer was added to each well, and the plate was placed on a shaker for 4 hours at room temperature. The wells were washed and then incubated with tetramethylbenzidine substrate for 20 minutes in the dark at room temperature. The reaction was stopped by adding 1.0 M sulfuric acid, and the absorbance was read at 450 nm using a microplate reader. A standard curve was run using the standards provided with the Apoptosense assay kit. Quantification of the cleaved CK18 was achieved according to the manufacturer's directions.
DNA fragmentation assay
Monolayers of GI-101A cells were grown in T-25 culture flasks and incubated with different concentrations of bromelain and without bromelain for 24 hours. After incubation the cells were harvested and washed with PBS. Cells were resuspended in 200 μL of PBS, and 20 μL of proteinase K was added. The DNA was extracted using the Qiagen (Chatsworth, CA, USA) DNeasy kit by following the manufacturer's protocol. The samples were subjected to electrophoresis at 80 V for 2 hours in 1.5% agarose gel containing 5 μL of ethidium bromide. Separated DNA fragments were viewed with a UVP (Upland, CA, USA) image analyzer.
4,6′-Diamino-2-phenylindole fluorescence staining
4,6′-Diamino-2-phenylindole (DAPI) is a nuclear stain that binds to DNA, allowing for DNA visualization when the stain fluoresces. This staining also enables the viewing of chromatin structures that are condensing in cells that are undergoing apoptosis. For conducting this experiment the GI-101A cells were plated at a density of 1×105 cells per well in a six-well plate, grown in complete RPMI growth medium, and treated with increasing bromelain concentrations: 0, 5, 10, and 20 μg/mL. Following the 24-hour treatment the cells were fixed with 3.7% paraformaldehyde and incubated with 0.1% Triton X-100 in PBS at room temperature for 20 minutes. After this incubation, the cells were washed gently with PBS and then incubated for another 10 minutes at room temperature with 200 ng/mL DAPI solution. At the end of the incubations, the cells were resuspended in PBS and examined quickly under an Olympus (Center Valley, PA, USA) fluorescent microscope (model BX51) with appropriate fluorescence filters and differential interference contrast optics. Images were captured at ×100 magnification using an Olympus DP70 digital camera and associated imaging software.
Statistical analysis
The data presented here represent mean±SD values from at least four individual experiments. Statistical analyses were performed using a one-way analysis of variance followed by Student–Newman–Keuls multiple comparisons tests. Values of P<.05 were considered as significant and are presented in Results.
Results
Bromelain-induced cell death and caspase activities
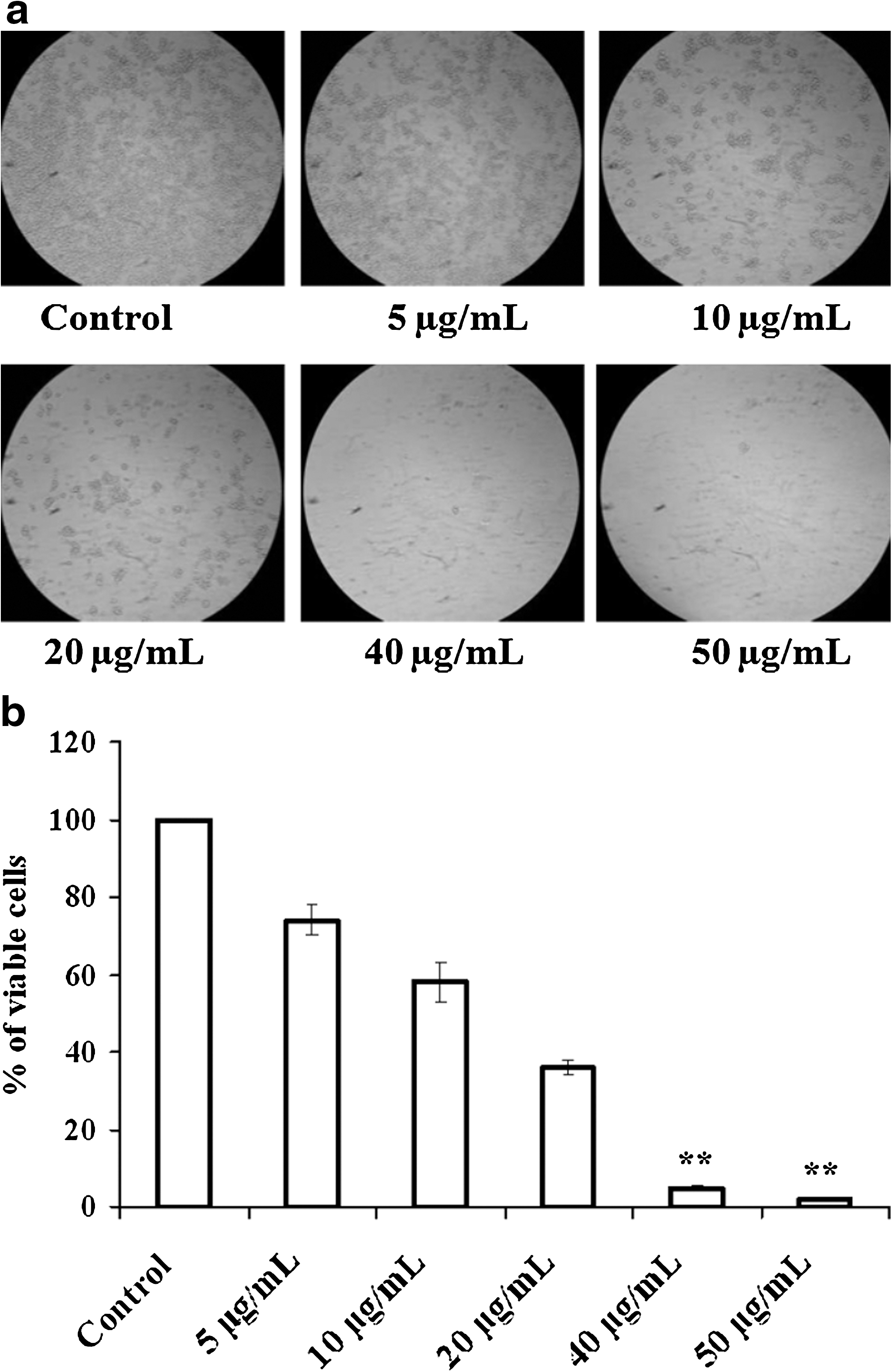
A decrease in viable cell number was observed in GI-101A breast cancer cells after treatment with different concentrations of bromelain (5, 10, 20, 40, and 50 μg/mL) following 24 hours of incubation (Fig. 1a). A bromelain concentration of 20 μg/mL was found to effectively reduce the percentage of viable cells to 36% after a 24-hour treatment. Doses higher than 20 μg/mL caused cell death in the range of 95% or greater within 24 hours (Fig. 1b). Because caspase-9 is an initiator caspase in the apoptotic pathway, activation of this isoform leads to cleavage of procaspase-3 into its active form. Therefore, we analyzed the activities of caspase-9 and caspase-3. Results in Figure 2 clearly show that the cells treated with bromelain exhibited a significant increase in the activities of both caspase-9 and caspase-3 after 24 hours with 5, 10, and 20 μg/mL concentrations.

Activities of caspase-9 and caspase-3 in GI-101A cells following a 24-hour bromelain treatment. Caspase-9 and caspase-3 activities were measured using the synthetic tetrapeptide substrates acetyl-Asp-Glu-Val-Asp-p-nitroaniline and acetyl-Leu-Glu-His-Asp-p-nitroaniline, respectively. Data are mean±SD values of four or more experiments. **P<.01 for comparison with respective controls.
M30-Apoptosense ELISA immunoassay
The M30-Apoptosense results showed that the cells treated with bromelain contained significantly higher levels of cleaved CK18 containing the CK18-Asp396 neo-epitope than the control cells. Additionally, as the bromelain concentration increased, the level of CK18 containing CK18-Asp396 neo-epitope also increased as shown in Figure 3.
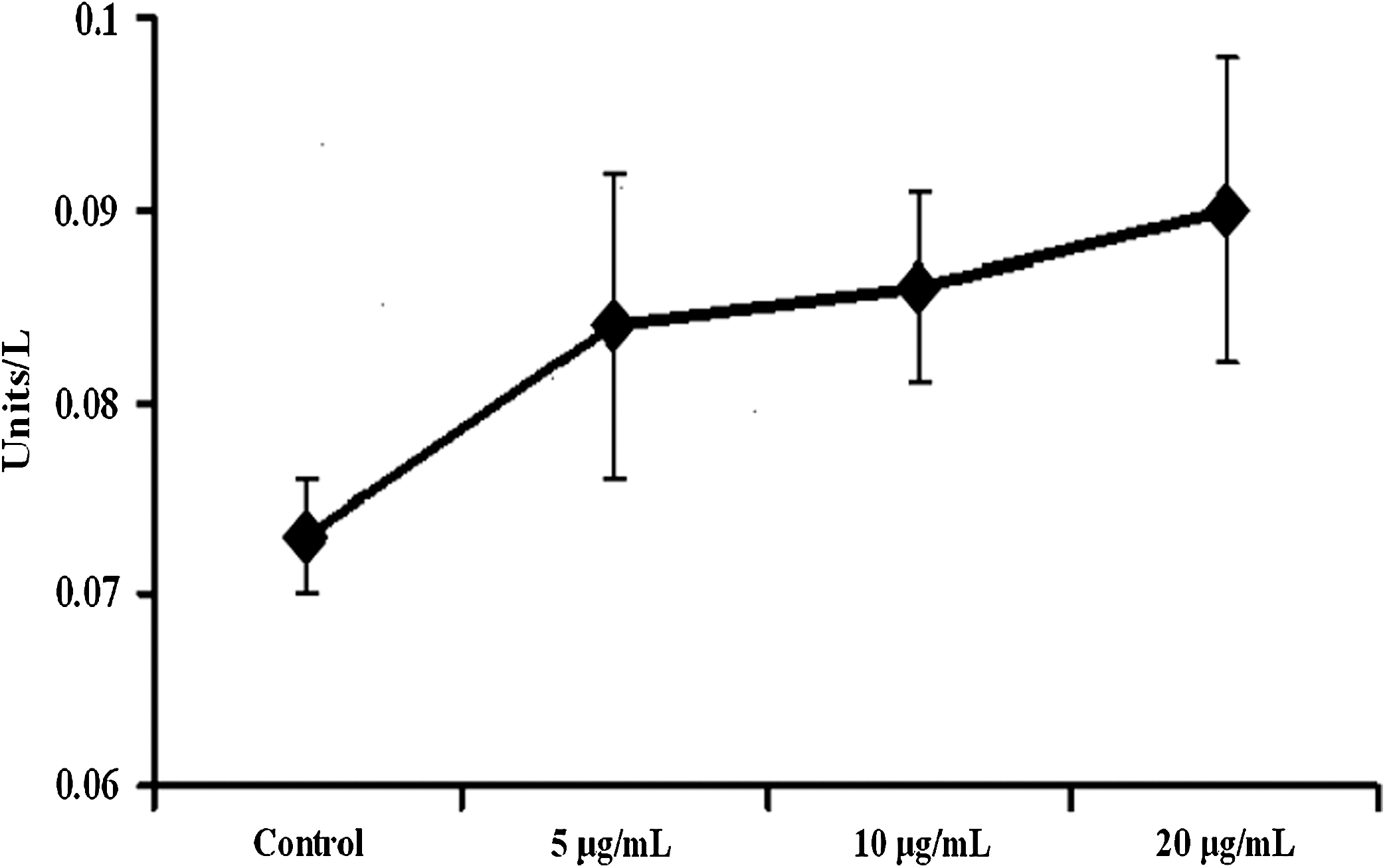
M30-Apoptosense levels by enzyme-linked immunosorbent assay in the GI-101A cell line following a 24-hour bromelain treatment. Data are mean±SD values of four or more experiments.
Effect of bromelain treatment on DNA fragmentation
To determine whether bromelain treatment induced DNA fragmentation, DNA was isolated from treated and untreated control GI-101A cells and separated by agrose gel electrophoresis. A typical ladder pattern of internucleosomal fragmentation of DNA was observed in cells treated with bromelain (Fig. 4, lanes 3–5), whereas in untreated control cells no fragmentation of DNA was observed (Fig. 4, lane 2).

A representative photograph of the agarose gel showing DNA fragmentation following bromelain treatment: lane 1, DNA marker; lane 2, DNA from control cells; and lanes 3–5, DNA from bromelain-treated cells, 5, 10, and 20 μg/mL respectively. The DNA was separated by electrophoresis using 1.5% agarose gel. DNA fragments stained with ethidium bromide were visualized using a UVP image analyzer.
Determination of apoptosis using DAPI staining
The DAPI staining method was used to assess the extent of apoptosis in control and bromelain-treated GI-101A cells. The DAPI staining results, shown in Figure 5, revealed good contrast between the control group (lightly stained) and the cells treated with bromelain (brightly stained). The bromelain-treated cells, especially those treated with 10 and 20 μg/mL, showed notable chromatin condensation and fragmentation compared with the control sample.
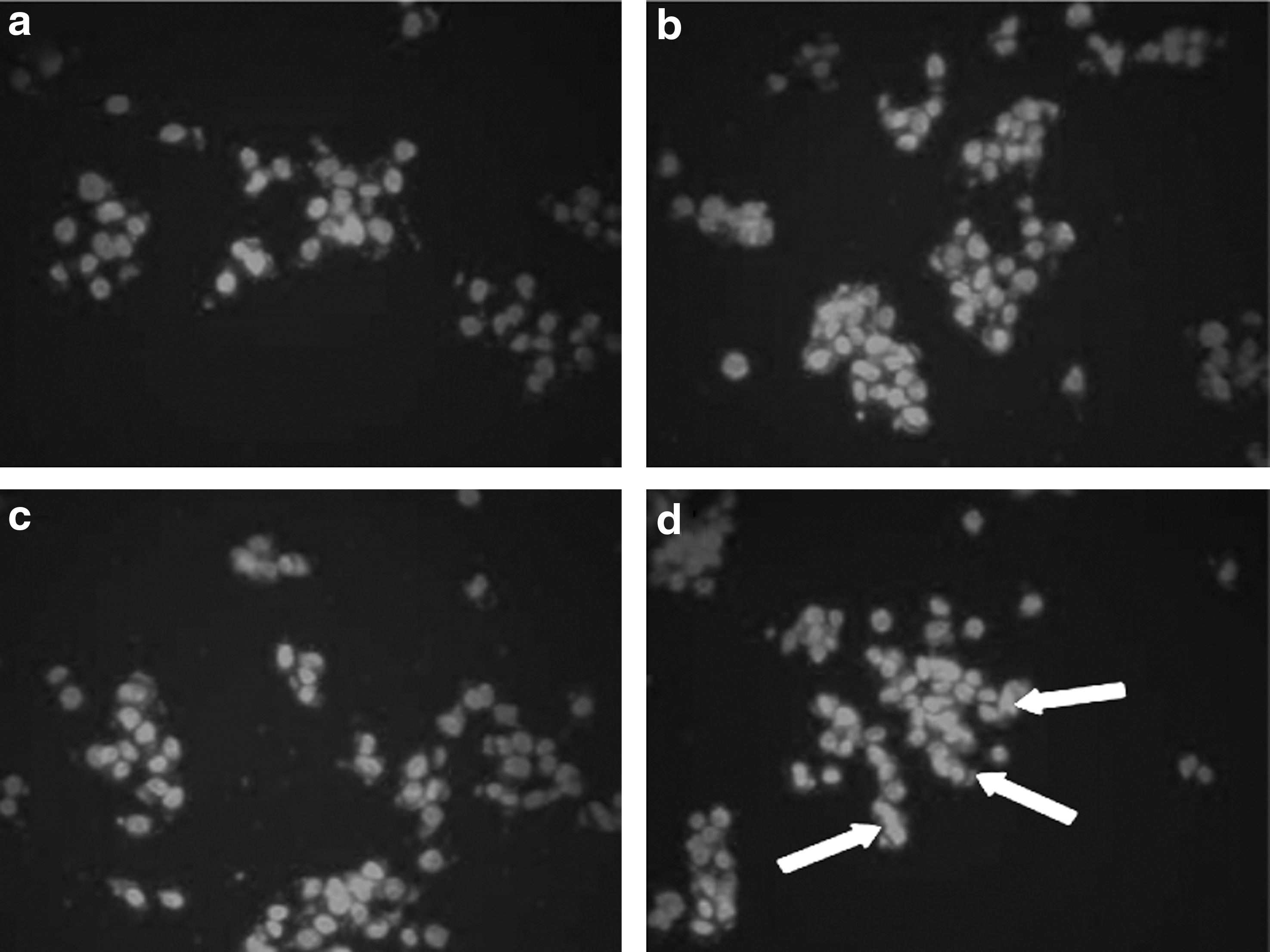
Representative photographs of control and bromelain (5, 10, and 20 μg/mL)-treated GI-101A cells stained with 4,6′-diamino-2-phenylindole after 24 hours of treatment:
Discussion
Over the past two decades, determination of the pharmacological effects of bioactive compounds used for cancer treatment and prevention has increased dramatically. 12 Therefore, evaluation of the potential benefits of bioactive compounds that are derived from consumable fruits and vegetables may lead to the identification of additional chemopreventive tools and strategies. Bromelain, a proteolytic enzyme isolated from pineapple, has been reported to possess antimetastatic and antitumorigenic activity. 6 In the present study, GI-101A breast cancer cells treated with bromelain resulted in a significant decrease in proliferation by inducing apoptosis. Because disrupted apoptotic progression could lead to cancer growth, induction of apoptosis in cancer cells by bioactive compounds is a key target for chemotherapeutic and chemopreventive applications. Our study demonstrates that bromelain may induce apoptosis in breast cancer cells through activation of a caspase-dependent pathway. Bromelain induced a maximum of 67% cell death at a concentration of 20 μg/mL with a 24-hour treatment interval, and it was almost 100% beyond that concentration.
Induction of apoptosis observed in bromelain-treated GI-101A cells is consistent with previous reports that bromelain induces apoptosis by activating the preexisting apoptosis machinery through caspase-3. Once activated, the effector caspase-3 appears to stimulate the DNase and cause DNA fragmentation. From the results obtained in our study, it is evident that the increase in the activity of caspase-3 and caspase-9 plays a pivotal role in apoptotic cell death induced by bromelain. Apoptotic protease activating factor 1, cytochrome c, and caspase-9 are the actual important participants in a complex pathway necessary for caspase-3 activation. Some of the in vitro studies have shown that depletion of caspase-9 from cytosolic fractions could result in the failure of caspase-3 activation. 13 The results from this study have demonstrated that bromelain significantly induced the activation of pro-caspase-9 to its active form and furthermore amplified the activity of caspase-3 to cause apoptosis.
A consequence of induction of apoptosis is the damage to the cytoskeleton, where the cytokeratins are found in abundance, resulting in the release of CK18 to the extracellular environment (caspases). 14,15 Recently, it has been shown that the determination of cell death (e.g., apoptosis mode) is possible by measuring the soluble CK18 fragments (CK18-Asp396 neo-epitope, also called M30 antigen) that are formed by caspase activation. 16 CK18 is a major component of the intermediate filament of simple epithelial cells and epithelial-derived tumors and makes up approximately 5% of total protein. 17 It undergoes proteolytic cleavage during apoptosis into fragments, 11,18 exposing the CK18-Asp396 neo-epitope. 19,20 Treatment with the proteolytic enzyme bromelain was found to enhance the level of CK18 in the GI-101A breast cancer cell line in our study. Recently it has been reported that an increased level of CK18 in a patient's serum with breast cancer, after treatment with docetaxel, was indicated as the primary measure of cell death due to apoptosis. 21 Our results are similar to the observation reported earlier in terms of intracellular cleavage and increase in the levels of CK18 during apoptosis. However, we did not measure the extracellular levels of CK18 in our samples, and therefore it is not reported here. It appears that the intracellular cleavage of CK18 in estrogen receptor–positive GI-101A cells may occur by a mechanism similar to the one that exists in estrogen receptor–negative MDA-MB-231 cells, indicating that the apoptotic mechanism leading to CK18 cleavage was probably estrogen receptor independent.
The characteristics of apoptotic cell death are the induction of chromatin condensation, fragmentation of nuclei, fragmentation of DNA, and cleavage of specific proteins. 22,23 Several anticancer agents are being tested these days that are expected to kill cancer cells by inducing pro-apoptotic signals. In this regard, we have so far determined that bromelain treatment induces chromatin condensation and internucleosomal fragmentation of DNA in GI-101A cells. The morphological changes of apoptosis in most of the cell types are contraction in cell volume and condensation of the nucleus, which allows the intracellular organelles to retain their normal morphology. This change is followed by the plasma membrane blebbing and nuclear fragmentation to form apoptotic bodies. 24 A closer look at the pattern of DAPI staining in bromelain-treated GI-101A cells in our study suggests that DNA fragmentation is initiated at nuclear periphery and progresses toward the center. Although DAPI staining enables the determination of cells undergoing apoptosis, DNA from the cells forms a characteristic ladder pattern on agarose gel that also confirms the biochemical changes involved in the fragmentation of chromosomes into nucleosome units. 25 As shown in our results, multiple units of apoptotic DNA ladder were detected in bromelain-treated GI-101A cells, whereas in the control, there was no such fragmentation.
Finally, our results confirm the cytotoxic effects of bromelain on the GI-101A breast cancer cells in a dose-dependent manner. Furthermore, bromelain increased the level of CK18, one of the markers indicating the cell death via apoptosis. Our results also suggest that bromelain induced the apoptotic signal through activation of a caspase pathway, resulting in nuclear condensation and disintegration, which are hallmarks of apoptotic cell death.
Footnotes
Acknowledgments
Financial support from Chancellor's Faculty Research and Development Grant of Nova Southeastern University is gratefully acknowledged. We also would like to thank the Royal Dames of Cancer Research, Ft. Lauderdale Inc., for their generous support in conducting this research.
Author Disclosure Statement
No competing financial interests exist with this work.